Evaluation von RNA-Extraktionsmethoden zur Bestimmung von HLA-G im Überstand von Embryonenkulturen nach ART - unipub
←
→
Transkription von Seiteninhalten
Wenn Ihr Browser die Seite nicht korrekt rendert, bitte, lesen Sie den Inhalt der Seite unten

Manuela Mißbach Evaluation von RNA-Extraktionsmethoden zur Bestimmung von HLA-G im Überstand von Embryonenkulturen nach ART Masterarbeit zur Erlangung des akademischen Grades Master of Science im Rahmen des Universitätslehrganges Klinische Embryologie Wissenschaftliche/r BegutachterIn: Univ.-Prof. Mag. Dr. Thomas Ebner Ao.Univ.-Prof. Mag. DDr. Erwin Petek Karl-Franzens-Universität Graz und UNI for LIFE Aßling, Juni 2022

Ehrenwörtliche Erklärung Ich erkläre ehrenwörtlich, dass ich die vorliegende Arbeit selbstständig und ohne fremde Hilfe verfasst, andere als die angegebenen Quellen nicht benutzt und die den Quellen wörtlich oder inhaltlich entnommenen Stellen als solche kenntlich gemacht habe. Die Arbeit wurde bisher in gleicher oder ähnlicher Form keiner anderen inländischen oder ausländischen Prüfungsbehörde vorgelegt und auch noch nicht veröffentlicht. Die vorliegende Fassung entspricht der eingereichten elektronischen Version. 12.06.2022 Manuela Mißbach
Danksagung Mein besonderer Dank an dieser Stelle gilt Herrn Professor Dr. Dr. Wolfgang Würfel für die Überlassung der interessanten Thematik und der fachlichen Expertise. Des Weiteren bedanke ich mich bei Dr. Ralph Wirtz und all seinen Mitarbeitern der STRATIFYER Molecular Pathology GmbH, Köln, die es ermöglichten, die gesammelten Überstände aus Embryonenkulturen zu analysieren und die somit grundlegend an der Entstehung dieser Studienarbeit beteiligt waren. Ein großer Dank geht ebenso an Franziska Würfel, für die Vermittlung und Bearbeitung der Proben im molekularpathologischen Labor STRATIFYER sowie der Hilfe und Unterstützung bei wissenschaftlichen Fragen. Außerdem möchte ich mich bei meinen Kollegen im IVF-Labor und der Geschäftsleitung der Praxisklinik Kinderwunsch Centrum München bedanken, die mir die Zeit zur Durchführung des Studiums zur Verfügung gestellt haben, um meine beruflichen Ziele erreichen zu können. Vielen Dank an meine Familie, die mir den Rücken für das Schreiben dieser Masterarbeit freigehalten haben, für die mentale Unterstützung sowie für das Vertrauen und die Zuversicht in die Fertigstellung dieser Studienarbeit.

Inhaltsangabe Unter den verschiedenen Mechanismen zum Schutz des Embryos/Fetus vor der Ab- stoßungsreaktion durch das mütterliche Immunsystem spielt die Expression der HLA- Klasse-Ib-Moleküle (HLA-E, -F, -G) eine besondere Rolle. Der Embryo blockiert einen Angriff mittels der Interaktion von embryonalen HLA-Gruppen mit spezifischen Rezeptoren der Immunzellen. Aus seiner geschützten Position heraus werden uterine NK-Zellen durch das fremde HLA-C aktiviert und unterstützen somit den sich einnistende Embryo durch die Freisetzung von Zytokinen und Wachstumsfaktoren. Ziel der vorliegenden Studie war es, durch die Evaluierung verschiedener Extraktionsmethoden, HLA-G-mRNA aus dem Überstand von Embryonenkulturen zu gewinnen und mittels qRT-PCR semiquantitativ nachzuweisen. Anschließend sollte ein Zusammenhang zwischen dem HLA-G-Nachweis und dem Zyklusausgang in Abhängigkeit der Behandlungsmethode (Frischzyklus versus Kryozyklus), der Embryonenqualität (A, B, C) und der Kulturdauer (ET Tag 2/3 versus Tag 5) ermittelt werden. Um die RNA-Extraktionsmethoden vergleichen zu können, erfolgte eine statistische Auswertung basierend auf den Testgütekriterien der Sensitivität, der Spezifität, des positiven und negativen Vorhersagewerts sowie der Korrekt- und Falschklassifikationsrate. Im ersten Durchlauf wurden verschiedene Methoden zur Gewinnung von mRNA ausgetestet, wobei die Stratifyer Blutextraktion am häufigsten angewendet wurde. Diese Gewinnungsmethode konnte aber wegen seiner geringen Spezifität (71% der Leerproben mit positivem HLA-G-mRNA-Nachweis) nicht überzeugen. Auf Grund dessen kam in der zweiten Testserie hauptsächlich der zuverlässigere miRNeasy Serum/Plasma Kit (Qiagen) zum Einsatz (87% der Leerproben ohne HLA-G-RNA- Nachweis). Insgesamt wurde in 55% der Proben HLA-G-mRNA nachgewiesen (36,2% in E, 18,6% in L). In Kryozyklen wurde etwas häufiger HLA-G-mRNA nachgewiesen als in Frischzyklen. Hinsichtlich der Embryonenqualität konnte eine gleichmäßige Verteilung des HLA-G-Nachweises auf Embryonen guter und schlechter Qualität festgestellt werden, wobei qualitativ gute Embryonen eine höhere SSR ergaben. In Proben nach Kultur bis Tag 2/3 wurde häufiger HLA-G nachgewiesen, jedoch bei einer geringeren SSR. Der Transfer von Tag-5-Embryonen resultierte in einer hohen SSR, obwohl weniger als die Hälfte der Embry- okulturproben einen HLA-G-mRNA-Nachweis zeigten. Im Gegensatz zur Literatur konnte eine Korrelation zwischen HLA-G-Nachweis und Zyklusausgang nicht dargestellt werden.

Abstract The expression of HLA class Ib molecules (HLA-E, -F, -G) plays a special role among the various mechanisms for protecting the embryo/fetus from rejection by the maternal immune system. The embryo blocks an attack through the interaction of embryonic HLA groups with specific receptors on immune cells. From its protected position, uterine NK cells are activated by the foreign HLA-C and thus support the implanting embryo by releasing cytokines and growth factors. The aim of the present study was to obtain HLA-G mRNA from the supernatant of embryo cultures by evaluating different extraction methods and to detect it semi- quantitatively using qRT-PCR. Subsequently, a connection between the HLA-G detection and the cycle outcome depending on the treatment method (fresh cycle versus cryocycle), embryo quality (A, B, C) and culture duration (transfer at day 2/3 versus day 5) should be determined. In order to be able to compare the RNA extraction methods, a statistical evaluation was carried out based on the test quality criteria of sensitivity, specificity, the positive and negative predictive value as well as the correct and incorrect classification rate. In the first run, various methods for obtaining mRNA were tested, with the Stratifyer blood extraction being used most frequently. However, this extraction method was not convincing due to its low specificity (71% of the blank samples with positive HLA- G mRNA detection). For this reason, the more reliable miRNeasy Serum/Plasma Kit (Qiagen) was mainly used in the second test series (87% of the blank samples without HLA-G-RNA detection). Overall, HLA-G mRNA was detected in 55% of the samples (36.2% in embryo culture sample, 18.6% in blank). HLA-G mRNA was detected slightly more frequently in cryocycles than in fresh cycles. With regard to embryo quality, HLA-G detection was evenly distributed between good and poor quality embryos, with good quality embryos yielding a higher pregnancy rate. HLA-G was detected more frequently in samples post-culture up to day 2/3, but at a lower pregnancy rate. Transfer of day 5 embryos resulted in a high pregnancy rate, although less than half of the embryo culture samples showed HLA-G mRNA detection. In contrast to literature no correlation between HLA-G detection and cycle outcome could be shown.
Inhaltsverzeichnis Inhaltsverzeichnis Inhaltsverzeichnis ....................................................................................................... 1 Abbildungsverzeichnis ................................................................................................ 4 Tabellenverzeichnis .................................................................................................... 5 Formelverzeichnis ...................................................................................................... 7 Abkürzungsverzeichnis ............................................................................................... 8 1. Einführung.......................................................................................................... 11 2. Theorie ............................................................................................................... 12 2.1 Das HLA-System (Human-Leukozyten-Antigen-System) ............................ 12 2.1.1 HLA-Klasse-I ......................................................................................... 14 2.1.2 HLA-Klasse-II ........................................................................................ 15 2.1.3 HLA-Klasse-III ....................................................................................... 16 2.2 Die nicht-klassischen HLA-Gruppen E bis G (Klasse Ib) ........................... 17 2.2.1 HLA-E ................................................................................................... 17 2.2.2 HLA-F ................................................................................................... 18 2.2.3 HLA-G ................................................................................................... 19 2.3 Der Sonderfall: Die klassische HLA-Gruppe C (Klasse Ia) .......................... 21 2.4 Bedeutung einzelner immunkompetenter Zellen ......................................... 21 2.4.1 Natürliche Killerzellen (NK) ................................................................... 21 2.4.2 Lymphozyten......................................................................................... 23 2.4.3 Makrophagen ........................................................................................ 24 2.5 Die Schwangerschaft - ein Zusammenspiel von Fetus, Immunzellen und HLA-Molekülen ...................................................................................................... 25 2.5.1 Die Rolle des HLA-Systems in der Schwangerschaft ........................... 25 2.5.2 Immunologische Interaktion an der embryo-maternalen Grenzfläche ... 26 2.5.3 Die Bedeutung von HLA-G in der Schwangerschaft ............................. 30 1

Inhaltsverzeichnis 3. Ziel ..................................................................................................................... 33 3.1 Zielvorlage - ein Rückblick ........................................................................... 33 3.2 Zielstellung .................................................................................................. 34 4. Material und Methoden ...................................................................................... 35 4.1 Patientenkohorte ......................................................................................... 35 4.2 Embryonenkultur und Probengewinnung ..................................................... 35 4.3 RNA-Extraktionsmethoden .......................................................................... 37 4.3.1 Stratifyer Blutextraktion (STLE)............................................................. 37 4.3.2 Stratifyer Blutextraktion ohne Lysebuffer (STLE o. Lyse) ..................... 39 4.3.3 Stratifyer Blutextraktion mit ExoQuick® Buffer....................................... 39 4.3.4 exoRNeasy Kit Qiagen .......................................................................... 40 4.3.5 QIAamp® DNA Blood Kit Qiagen ........................................................... 42 4.3.6 miRNeasy Serum/Plasma Kit Qiagen ................................................... 43 4.3.7 Desoxyribonuklease-Verdau ................................................................. 45 4.4 Die PCR ...................................................................................................... 46 4.4.1 Das Prinzip der PCR ............................................................................. 46 4.4.2 Das Protokoll der qRT-PCR .................................................................. 47 4.5 Datenerfassung zu den Embryonenkulturen ............................................... 48 4.5.1 Behandlungsmethoden ......................................................................... 48 4.5.2 Embryonenqualität ................................................................................ 48 4.5.3 Embryonenkultur und Transfer.............................................................. 50 4.5.4 Zyklusausgang ...................................................................................... 50 4.6 Statistische Auswertung der Labormethoden .............................................. 51 4.6.1 Die Sensitivität ...................................................................................... 53 4.6.2 Die Spezifität ......................................................................................... 53 4.6.3 Der positive und negative Vorhersagewert ........................................... 54 4.6.4 Die Korrekt- und Falschklassifikationsrate ............................................ 55 2
Inhaltsverzeichnis 5. Ergebnisse ......................................................................................................... 56 5.1 Extraktionsmethoden und PCR-Ergebnisse ................................................ 56 5.1.1 Stratifyer Blutextraktion ......................................................................... 56 5.1.2 ExoRNeasy Kit Qiagen ......................................................................... 57 5.1.3 QIAamp DNA Blood Kit Qiagen ............................................................ 57 5.1.4 miRNeasy Serum/Plasma Kit Qiagen ................................................... 58 5.1.5 Zusammenfassung Extraktionsmethoden ............................................. 58 5.2 Ergebnisse der Embryonenkulturen ............................................................ 59 5.2.1 Behandlungsmethode, Zyklusausgang und HLA-G-mRNA-Nachweis .. 60 5.2.2 Embryonenqualität, Zyklusausgang und HLA-G-mRNA-Nachweis ....... 61 5.2.3 Embryotransfertag, Zyklusausgang und HLA-G-mRNA-Nachweis ....... 63 6. Diskussion .......................................................................................................... 65 6.1 Auswertung der Extraktionsmethoden und PCR-Ergebnisse ...................... 65 6.2 Auswertung der Embryonenkulturen ........................................................... 66 6.3 Fazit ............................................................................................................. 69 6.4 Schlusswort ................................................................................................. 72 Literaturverzeichnis .................................................................................................. 73 Anhang - Datenblätter............................................................................................... 80 a. Stratifyer Blutextraktion ................................................................................... 80 b. ExoRNeasy Kit Qiagen ................................................................................... 82 c. QIAamp DNA Blood Kit Qiagen ...................................................................... 82 d. miRNeasy Serum/Plasma Kit Qiagen ............................................................. 83 e. Embryonenkultur 1. Lauf ................................................................................. 85 f. Embryonenkultur 2. Lauf ................................................................................. 86 3

Abbildungsverzeichnis Abbildungsverzeichnis Abbildung 1: Grafische Darstellung des HLA-Locus auf dem Chromosom 6............ 13 Abbildung 2: Grafische Darstellung des MHC-Klasse-I-Proteinkomplexes ............... 15 Abbildung 3: Grafische Darstellung eines MHC-Klasse-II-Moleküls ......................... 16 Abbildung 4: Schematische Darstellung von HLA-E (Würfel, 2019) .................... 17 Abbildung 5: Grafische Darstellung der mRNA-Splicevarianten und Proteinisoformen ................................................................................................................................. 18 Abbildung 6: Grafische Darstellung der mRNA-Splicevarianten und der Protein- isoformen von HLA-G (Würfel, 2019)................................................................... 19 Abbildung 7: Funktion der uNK (Kuon, 2018) .................................................. 23 Abbildung 8: Die Plazenta mit ihrer HLA-Signatur als Pufferzone zwischen embryonalem und maternalem Gewebe ................................................................... 25 Abbildung 9: Wichtige immunologische Grenzflächen zwischen Embryo/Fetus und Mutter nach unterschiedlichem Gestationsalter ........................................................ 27 Abbildung 10: An der Immuntoleranz beteiligte Immunzellen mit Darstellung ihrer Rezeptoren und Signalproteine (Chemokine, Zytokine, Wachstumsfaktoren) .......... 28 Abbildung 11: Expression der HLA-Klasse-I und -II in der menschlichen Plazenta .. 28 Abbildung 12: Übergang der Steuerung der Schwangerschaft von den uNKs auf die Lymphozyten sowie Expression und Freisetzung von HLA-E, -F und -G im Schwangerschaftsverlauf.......................................................................................... 29 Abbildung 13: Rolle von HLA-G an der embryo- / feto-maternalen Grenzfläche ...... 31 Abbildung 14: Qualitätsmerkmale der Embryonen im Teilungsstadium am Tag 2 und Tag 3……………………………………………………………………………………49 Abbildung 15: Morphologische Kriterien der Blastozyste am Tag 5 ...................... 49 Abbildung 16: Einteilung der Qualitätsmerkmale am Tag 5 zur Vergabe der Note A, B oder C………………………………………………………………………………….49 Abbildung 17: Verteilung der Proben auf die Labormethoden ............................... 52 4
Tabellenverzeichnis Tabellenverzeichnis Tabelle 1 Arbeitsprotokoll XTRAKT BLOOD Kit, Firma Stratifyer ............................. 39 Tabelle 2 Arbeitsprotokoll ExoQuick®-Fällung vor der Anwendung von STLE und Qiagen DNA-Extraktion ............................................................................................ 40 Tabelle 3 Arbeitsprotokoll exoRNeasy Kit Qiagen .................................................... 41 Tabelle 4 Arbeitsprotokoll QIAamp® DNA Blood Kit Qiagen ..................................... 43 Tabelle 5 Arbeitsprotokoll miRNeasy Serum/Plasma Kit Qiagen .............................. 45 Tabelle 6 Arbeitsprotokoll DNase I Verdau ............................................................... 45 Tabelle 7 Statistik für Stratifyer Blutextraktion .......................................................... 56 Tabelle 8 Statistik für ExoRNeasy Kit ....................................................................... 57 Tabelle 9 Statistik für QIAmp DNA Blood Kit ............................................................ 57 Tabelle 10 Statistik für miRNeasy Serum/Plasma Kit ............................................... 58 Tabelle 11 Ergebnisse und Testgütekriterien der Extraktionsmethoden nach PCR- Analyse………………………………………………………………………………………59 Tabelle 12 Zusammenfassung Zyklusausgang und HLA-G-mRNA-Nachweis ......... 59 Tabelle 13 Übersicht zu Behandlungsmethode, HLA-G-mRNA-Nachweis und Zyklusausgang des 1. Laufs ..................................................................................... 60 Tabelle 14 Übersicht zu Behandlungsmethode, HLA-G-mRNA-Nachweis und Zyklusausgang des 2. Laufs ..................................................................................... 60 Tabelle 15 a) - d) Darstellung des Verhältnisses HLA-G-Nachweis zu Zyklusausgang von Frisch- und Kryozyklen im 1. und 2. Lauf ........................................................... 61 Tabelle 16 Übersicht zur Embryonenqualität, Zyklusausgang und HLA-G-mRNA- Nachweis des 1. Laufs.............................................................................................. 62 Tabelle 17 Übersicht zur Embryonenqualität, Zyklusausgang und HLA-G-mRNA- Nachweis des 2. Laufs.............................................................................................. 62 Tabelle 18 a) - b) Darstellung des Verhältnisses HLA-G-mRNA-Nachweis zu Zyklusausgang entsprechend der Embryonenqualität A+B sowie C jeweils im 1. und 2. Lauf……………………………………………………………………………………….63 Tabelle 19 Übersicht zu Embryotransfertag, Zyklusausgang und HLA-G-mRNA- Nachweis des 1. Laufs.............................................................................................. 63 Tabelle 20 Übersicht zu Embryotransfertag, Zyklusausgang und HLA-G-mRNA- Nachweis des 2. Laufs.............................................................................................. 64 5
Tabelle 21 a) - b) Darstellung des Verhältnisses HLA-G-Nachweis zu Zyklusausgang am Tag 2 bzw. 3 des Embryotransfers ..................................................................... 64 6
Formelverzeichnis Formelverzeichnis Formel 1 Berechnung der Sensitivität der Labormethode .................................... 53 Formel 2 Berechnung der Spezifität der Labormethode....................................... 53 Formel 3 Berechnung des positiven Vorhersagewerts ......................................... 54 Formel 4 Berechnung des negativen Vorhersagewertes ..................................... 54 Formel 5 Berechnung der Korrektklassifikationsrate ............................................ 55 Formel 6 Berechnung der Falschklassifikationsrate ............................................. 55 7
Abkürzungsverzeichnis Abkürzungsverzeichnis APC antigen presenting cells ART artificial reproductive technology Breg regulatorische B-Zellen BZ Blastozyste Calm2 Calmodulin2 COC cumulus oocyte complex CT Zytotrophoblast DC dendritische Zellen E Embryokulturprobe ELISA Enzyme-Linked Immunsorbet Assay ER Endoplasmatisches Retikulum ERA endometrial receptivity array ET Embryotransfer EtOH Ethanol EV extrazelluläre Vesikel EVTs extravillöse Trophoblastzellen Ex Exon FKR Falschklassifikationsrate G-CSF Granulocyte-Colony Stimulating Factor, GM-CSF Granulozyten-Makrophagen-Kolonie-stimulierenden Faktor HCG humanes Choriongonadotropin HELLP Haemolysis Elevated Liver enzymes Low Platelet count HLA Human-Leukozyten-Antigen ICM Inner Cell Mass ICSI Intrazytoplasmatische Spermieninjektion IFN-γ Interferon-gamma IL Interleukin ILT Lymphozyten-immunglobulinähnliche Transkripte IVF In-Vitro-Fertilisation 8
Abkürzungsverzeichnis KIR Killer-immunglobulinähnlicher Rezeptor KKR Korrektklassifikationsrate KLAR Killer-lezithinähnlicher Rezeptor L Leerprobe LILR Lymphozyten-immunglobulinähnlicher Rezeptor LILRB1 Lymphozyten-immunglobulinähnlicher-Rezeptor-B1 MHC Major Histocompatibility Complex MMIF/MIF Makrophagen-Migrations-Inhibitionsfaktor MMP Matrix-Metalloproteasen NK Natürliche Killerzellen NKG/NKG2 NK-Rezeptoren der Gruppe 2 NKR NK-Rezeptoren NPV negative predictive value OC open conformer PIBF Progesteron-induzierter Blockierfaktor PID Präimplantationsdiagnostik PN Pronuclei pNK periphere Natürliche Killerzellen PPV positive predictive value PRP Pattern Recognition Receptors RIF recurrent implantation failure RSA recurrent spontaneous abortion RT Reverse Transkription SE Sensitivität sHLA soluble HLA SNPs Single Nucleotide Polymorphism SS Schwangerschaft ß2m beta-2-Mikroglobulin SSR Schwangerschaftsrate SSW Schwangerschaftswoche ST Syncytiotrophoblast STLE Stratifyer Blutextraktion SZ Spezifität 9
Abkürzungsverzeichnis TE Trophektoderm TH T-Helferzellen TNF-α Tumornekrosefaktor-alpha Treg regulatorische T-Zellen TRP thrombozytenreiches Plasma uNK uterine Natürliche Killerzellen 10
Einführung 1. Einführung Die Geburt eines einzelnen gesunden Kindes und die Vermeidung von Mehrlings- schwangerschaften, die nicht nur mit medizinischen sondern auch mit sozial- ökonomischen Problemen verbunden sind, ist zentrales Ziel einer erfolgreichen re- produktionsmedizinischen Behandlung. Dabei ist die Übertragung von nur einem Embryo anzustreben. Es wurden große Fortschritte erzielt, um Kriterien zu etablie- ren, die den Präimplantationsembryo identifizieren, der das höchste Potential hat, sich im Uterus der Frau nach Transfer einzunisten und sich weiterzuentwickeln. Hauptsächlich wird die Qualität der Präimplantationsembryonen immer noch an Hand der Morphologie bewertet, die aber oftmals nicht mit der Fähigkeit zur Einnistung übereinstimmt. Zunehmend rücken weitere nicht-invasive Methoden in den Fokus der Forschung, wie z. B. die Bestimmung von Stoffwechselprodukten (sog. Metabolom) in der Embryonalkultur. Solch ein nicht-invasiver Marker kann ein wichtiges Instru- ment sein, um die Implantationsrate zu erhöhen und gleichzeitig die Zahl der zu transferierenden Präimplantationsembryonen, zur Eindämmung von Mehrlings- schwangerschaften, zu reduzieren. (Rebmann, 2010; Diaz, 2019) Der entscheidende Schritt für den Eintritt einer Schwangerschaft ist die erfolgreiche Implantation. Durch den Einsatz der Präimplantationsdiagnostik (PID) zur Untersu- chung der Präimplantationsembryonen vor dem Transfer in die Gebärmutter konnte gezeigt werden, dass ein wiederholtes Implantationsversagen (RIF) auch bei Embry- onen stattfindet, die morphologisch und genetisch unauffällig sind. Ursache dafür kann neben der Embryonenqualität eine schlechte Rezeptivität des Endometriums sein. (Kuon, 2018) Neben der Interaktion des Präimplantationsembryos mit dem Endometrium spielen möglicherweise immunologische Faktoren eine Rolle. Folgt man den Gesetzen der Transplantationsmedizin, so müsste der Fötus als semiallogenes Transplantat vom Immunsystem der Mutter abgestoßen werden (Kuon, 2018). Stattdessen entwickelt sich eine maternale Immuntoleranz, vermittelt durch eine komplexe Interaktion im- munkompetenter Zellen der Mutter mit dem Embryo. Defekte in der gegenseitigen immunologischen Anpassung zwischen Embryo und Mutter können die Plazentaent- wicklung und das Wachstum des Embryos beeinträchtigen (Tersigni, 2020). Eine Fehlsteuerung immunologischer Prozesse kann zu Schwangerschaftskomplikatio- 11
Theorie nen, wie Aborten und rezidivierendem Implantationsversagen (RIF), schlechter Pla- zentation und gestörter utero-plazentare Perfusion führen (Kuon, 2018; Tersigni, 2020). Circa ein bis drei Prozent der Kinderwunschpaare sind von wiederholten spontanen Fehlgeburten (RSA) betroffen. In der Hälfte der Fälle ist die Ursache unklar, weshalb die Diagnostik von infektiologischen und immunologischen Faktoren zielführend sein kann (Kuon, 2018). Unter den verschiedenen Schutzmechanismen, die beim Menschen in der Schwan- gerschaft involviert sein können, scheint die Expression nicht-klassischer HLA- Klasse-I-Moleküle und das völlige Fehlen der Expression von HLA-Klasse-II- Molekülen im trophoblastischem bzw. plazentarem Gewebe die wichtigsten embryo- fetalen Escape-Mechanismen zu sein. Die Bedeutung von HLA-Molekülen für die embryofetale Toleranz gegenüber dem mütterliche Immunsystem wird durch Schwangerschaftskomplikationen verdeutlicht, die in Fällen einer abnormalen Ex- pression der HLA-Moleküle an der embryo-/fetomaternalen Grenzfläche verursacht werden (Tersigni, 2020). 2. Theorie 2.1 Das HLA-System (Human-Leukozyten-Antigen-System) Das Human-Leukozyten-Antigen-System steht im zentralen Fokus der zellulären und immunologischen Kommunikation. Es spielt eine essentielle Rolle in der immunologi- schen Identifikation von körpereigenen, körperfremden und entarteten Zellen (Würfel F. M., 2019). Gegenwärtig existieren Studien über die Expression von HLA- Molekülen im Zusammenhang mit Fertilität, Krankheiten, Tumorerkrankungen und der Transplantation von Organen (Arnaiz-Villena, 2020). Die Durchführung des HLA-Matchings ist ein wichtiger diagnostischer Schritt vor ei- ner Organ- oder Knochenmarkstransplantation, denn bei Transplantationen besteht direkter Kontakt zwischen dem Spenderorgan, typisiert durch die HLA-Klasse I und II, und dem Immunsystem des Empfängers, das mehr oder weniger stark darauf rea- giert. Auch bei hoher HLA-Übereinstimmung ist zusätzlich eine immunsuppressive Therapie notwendig, damit das transplantierte Gewebe nicht abgestoßen wird. 12
Theorie Der Fetus besitzt humane Leukozyten-Antigene zu jeweils 50 Prozent von der Mutter und vom Vater, die in verschiedene Untergruppen gegliedert und von unterschiedli- chen Genorten codiert werden (Kuon, 2018). Auf Grund dessen wird der Embryo oft als „Semiallotransplantat“ bezeichnet, da er für das mütterliche Immunsystem zur Hälfte als „fremd“ erscheint. Bei z.B. der Eizellspende stimmen bekanntermaßen die Mutter und der Embryo zu 100 Prozent genetisch nicht überein, so dass man in die- sem Fall von einem „Allotransplantat“ oder „Holotransplantat“ spricht. Um eine er- folgreiche Schwangerschaft, auch aus Eizellspende, Leihmutterschaft oder Embryo- nenspende, zu ermöglichen, ist ein enges zelluläres Netzwerk zwischen Trophoblast und mütterlichen Gewebe erforderlich. Die Zellen des Trophoblasten exprimieren in dieser speziellen Situation die nicht-klassischen HLA-Gruppen zur Erhaltung der Schwangerschaft und um die Abwehrreaktion des mütterlichen Immunsystems zu umgehen. Das HLA-System ist Teil des Haupthistokompatibilitätskomplexes (engl. Major Histo- compatibility Complex, MHC) der Wirbeltiere, deren Oberflächenmoleküle durch eng gekoppelte Gene auf dem kurzen Arm von Chromosom 6 kodiert werden (Abbildung 1) (Mayer, 1997). Jeder Mensch erbt jeweils ein Allel von der Mutter und vom Vater und besitzt zwei HLA-Merkmale pro Genort. Das HLA-System wird aufgrund seiner biologischen Funktion in drei Klassen mit ihren entsprechenden Untergruppen unter- teilt. Abbildung 1: Grafische Darstellung des HLA-Locus auf dem Chromosom 6 Quelle: Acta Naturae 11(4):4-12 (Zakharova, 2019) 13
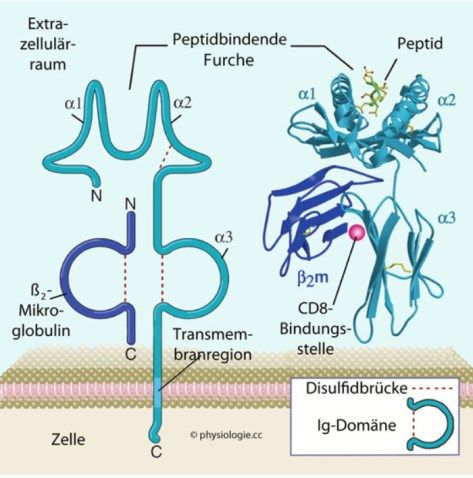
Theorie 2.1.1 HLA-Klasse-I Die HLA-Klasse-I-Antigene bestehen auf der Peptidebene aus drei alpha-Ketten, die die Bindungsstelle für das fremde Antigen, für den T-Zell-Corezeptor CD8 und dem löslichem beta-2-Mikroglobulin bilden (Butzeck, 2020). Die MHC der Gruppe I formen Heterodimere mit dem ß2-Mikroglobulin (Abbildung 2) (Würfel, Würfel, & Seelbach- Göbel, 2019). Für das ß2-Mikroglobulin sind keine Polymorphismen beschrieben. Es wird auf dem Chromosom 15 codiert. HLA-Klasse-I-Moleküle kommen nahezu auf allen kernhaltigen somatischen Zellen und Thrombozyten vor und sind sehr variabel, d.h. polymorph (Würfel, Würfel, & Seelbach-Göbel, 2019). Die Klasse Ia beinhaltet die klassischen MHC-I-Proteine HLA-A, HLA-B und HLA-C (Würfel, Würfel, & Seelbach-Göbel, 2019). Die nicht-klassischen Merkmale HLA-E, HLA-F und HLA-G der Klasse Ib sind wenig variabel, d.h. sie sind nahezu monomorph, gegebenenfalls finden sich Spleißvarian- ten. Die sog. Pseudogene (HLA-H bis -Z) in dieser Klasse zeichnen sich durch eine genetische Kodierung aus, die nach derzeitigem Kenntnisstand nicht für eine Pro- teinsynthese translatiert wird. Eine Ausnahme scheint HLA-H bei der Hämatochro- matose darzustellen (Feder, 1997). Die HLA-Antigene der Klasse Ib kommen so gut wie nicht im adulten Gewebe vor, davon ausgenommen sind Spermien, Hodengewebe oder einige ovarielle Zelleinhei- ten (Würfel, Würfel, & Seelbach-Göbel, 2019). Im Unterschied zur HLA-Klasse II werden auf den MHC-Gruppe-I-Molekülen intrazellulär produzierte Proteinfragmente, wie z. B. virale Antigene, gekoppelt, was zur Erkennung und Abtötung infizierter oder entarteter Zellen führt. 14
Theorie polymorphe α-Kette monomorphes ß2- Mikroglobulin Abbildung 2: Grafische Darstellung des MHC-Klasse-I-Proteinkomplexes Quelle: Nach einer Vorlage in Abbas / Lichtman / Pillai: Cellular and Molecular Immunology, 9th ed. 2018 (Abul K. Abbas, 2018) In der vorliegenden Arbeit steht vor allem das embryofetale Merkmal HLA-G der Klasse Ib im Mittelpunkt. 2.1.2 HLA-Klasse-II Der Proteinkomplex besteht aus zwei α- und zwei ß-Ketten. Die α1- und ß1-Kette bilden die Antigenbindungstelle und die ß2-Kette stellt die Bindungsstelle für den T- Zell-Corezeptor CD4 dar (Butzeck, 2020). Die Proteine der MHC-Klasse-II bilden ebenfalls Heterodimere (Abbildung 3). Sie besteht einerseits aus den drei großen Proteinen HLA-DP, -DQ und -DR, deren α-Ketten des Heterodimers im A-Lokus und deren ß-Ketten im B-Lokus kodiert werden, wonach sich die einschlägige Nomenkla- tur orientiert. Andererseits enthält die HLA-Klasse-II die zwei kleinen Proteine HLA- DM, -DO, die intrazellulär z.B. die Aufgabe des Processing von Antigenen erfüllen. Die HLA-Klasse-II-Moleküle werden auf phagozytierenden Zellen, wie Monozyten, Makrophagen und B-Lymphozyten exprimiert und kommen hauptsächlich auf den antigenpräsentierenden Zellen (APC) vor, die v.a. den T-Lymphozyten fremde Anti- gene von phagozytierten extrazellulären Erregern präsentieren sollen (Würfel, Würfel, & Seelbach-Göbel, 2019). Das HLA-II-Molekül liegt in inaktiver Form im Endoplasmatischem Retikulum (ER) vor. Fremdantigene werden in den Endosomen durch Proteasen in kurze Peptid- fragmente umgebaut (Richard, 2013). Das HLA-II-Molekül gelangt über den Golgi- Apparat zu den Endosomen und wird aktiviert. Somit wird im Endosom das fremde 15
Theorie Peptid an HLA gebunden und der Komplex an der Zelloberfläche präsentiert, wodurch T-Helferzellen angelockt werden (Butzeck, 2020). Abbildung 3: Grafische Darstellung eines MHC-Klasse-II-Moleküls Quelle: Nach einer Vorlage in Abbas / Lichtman / Pillai: Cellular and Molecular Immunology, 9th ed. 2018 (Abul K. Abbas, 2018) Die grafische Abbildung verdeutlicht die verschiedenen Regionen des MHC- Moleküls. Klasse-II-Moleküle bestehen aus einer polymorphen α-Kette, die nicht- kovalent an eine polymorphe ß-Kette gebunden ist. Beide Ketten sind glykosyliert; Kohlenhydratreste werden nicht dargestellt. Das Banddiagramm zeigt die Struktur des extrazellulären Teils des HLA-DR1-Moleküls mit einem gebundenen Peptid, das durch Röntgenkristallographie aufgelöst wurde (Abbildung 3). 2.1.3 HLA-Klasse-III Die Komplementfaktoren C2, C4, Properdinfaktor Bf und verschiedene Zytokine, wie z.B. der Tumornekrosefaktor (TNF) gehören zu den Klasse-III-Komplexen, die zwi- schen dem MHC-I- und -II-Komplex auf dem Chromosom 6 lokalisiert sind (Abbildung 1). Diese Plasmaproteine sind durchwegs an der unspezifischen Immunabwehr be- teiligt, im Gegensatz zu den anderen oben genannten Gruppen (Miller, 2020). 16
Theorie 2.2 Die nicht-klassischen HLA-Gruppen E bis G (Klasse Ib) 2.2.1 HLA-E Abbildung 4: Schematische Darstellung von HLA-E (Würfel, 2019) Das HLA-E-Antigen ist ein nicht-polymorphes HLA-Klasse-I-Molekül. Es sind von ihm 13 Allele bekannt, aus denen 5 Proteine resultieren (Nattermann, 2005). Für eine stabile Oberflächenexpression muss HLA-E an intrazelluläre Peptidsequenzen bin- den (Abbildung 4), die von Signalpeptiden der klassischen HLA-Klasse I und HLA-G bereitgestellt werden; es ist von ihnen abhängig. Ist die Expression von HLA-Ia und -G reduziert, dann wird auch HLA-E reduziert exprimiert. HLA-E wird in verschiede- nen kernhaltigen Zellen des menschlichen Organismus exprimiert. Die Expression findet aber hauptsächlich durch den Trophoblasten im ersten Trimester der Gestation statt und sie nimmt im weiteren Verlauf ab (Würfel F. M., 2019). HLA-E zielt vor allem auf die Natürlichen Killerzellen (NK). Es interagiert mit den NK- Rezeptoren der Gruppe 2 und wirkt z.B. via den NKG2A-Rezeptor inhibierend. Die NK-Zellaktivierung und -vermehrung wird gehemmt und die antikörperabhängige Zy- totoxizität vermindert. Durch die Aktivierung anderer Rezeptoren, besonders von NKG2D, können demgegenüber Zytokine und Wachstumsfaktoren durch die uterinen NKs freigesetzt werden. HLA-E besitzt auch antigenpräsentierende Eigenschaften, indem es zeitweise Teile der adulten HLA-Signatur des Embryos (HLA-A bis -DR) präsentiert und demzufolge immunkompetente Zellen anlockt. Die Darbietung der fremden Antigene findet unter dem Schutzmantel von HLA-E und der anderen emb- ryofetalen HLA-Gruppen statt, so dass das embryonale bzw. fetale Gewebe nicht abgestoßen wird. (Würfel, Würfel, & Seelbach-Göbel, 2019) 17
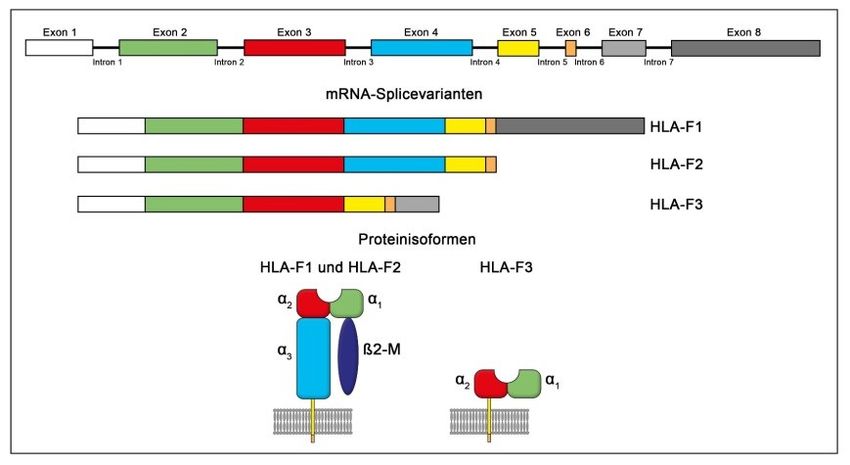
Theorie 2.2.2 HLA-F Abbildung 5: Grafische Darstellung der mRNA-Splicevarianten und Proteinisoformen von HLA-F (Würfel, 2019) HLA-F ist von den nicht-klassischen HLA-Genen am wenigsten bekannt. Bisher wur- den 22 HLA-F-Allele beschrieben, die für vier separate Proteine kodieren (Pan, 2013; Pyo, 2006). Es sind drei mRNA-Transkriptvarianten (HLA-F 1, 2, 3) bekannt (Abbildung 5). Die Expression ist hauptsächlich auf den intrazellulären Teil der Zelle begrenzt, speziell auf das ER. Das Molekül kann aber auch an der Oberfläche von extravillösen Trophoblastzellen und aktivierten lymphatischen Zellen (z.B. B- Lymphozyten) exprimiert werden (Würfel F. M., 2019; Lee, 2010; Langkilde, 2020). HLA-F tritt in zwei verschiedenen Formen auf, als Komplex mit beta-2-Mikroglobulin (ß2m) und einem Peptid (peptidgebundenes HLA-F-ß2m) und als offener Konformer (OC) ohne Peptid und ß2m (peptidfreies OC) (Lin, 2019). Im Komplex mit ß2m prä- sentiert es insbesondere Eigenpeptide. Der peptidgebundene HLA-F-ß2m-Komplex fungiert dagegen als Ligand für den inhibierenden Rezeptor LILRB1 (Lymphozyten- immunglobulinähnlichen-Rezeptor-B1), einen Hauptakteur bei der maternal-fetalen Toleranz. HLA-F ist fähig an aktivierenden und hemmenden NK-Zellrezeptoren zu binden (Ho, 2019). Die Expression von HLA-F nimmt im Schwangerschaftsverlauf zu. Es erscheint auf den Trophoblastzellen im ersten Trimenon erst in geringer Konzent- ration. Diese steigt in der Mitte des zweiten Trimenons dann deutlich an, zu einem Zeitpunkt, wo die Steuerung der Schwangerschaft von den Lymphozyten dominiert wird (Würfel, Würfel, & Seelbach-Göbel, 2019; Würfel F. M., 2019). 18
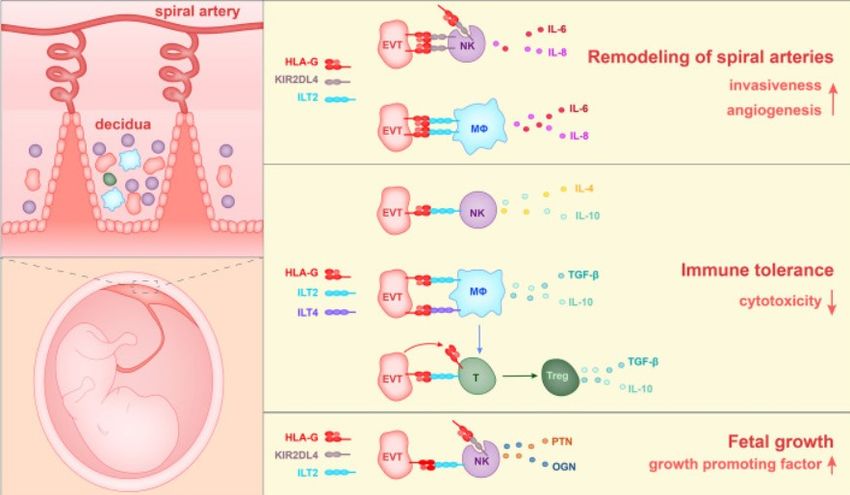
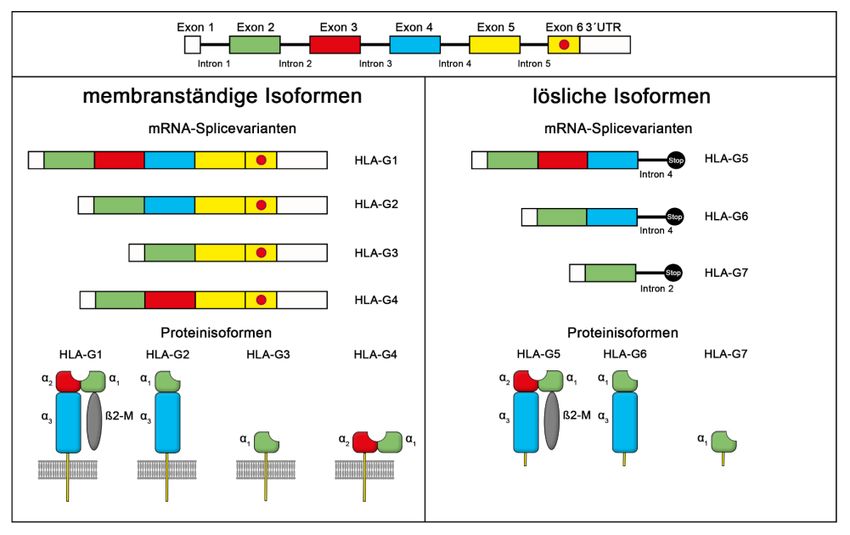
Theorie 2.2.3 HLA-G Das bekannteste Gen innerhalb der nicht-klassischen HLA-Gruppe ist HLA-G (Abbildung 6). Es ist ein sehr polymorphes Gen mit 75 Single Nucleotide Polymor- phismen (SNPs) auf der HLA-G-codierten Region. Es beinhaltet 74 Allele, die 24 Pro- teine in voller Länge kodieren und 4 Null-Allele, die die verkürzte Proteinform kodie- ren (IPD78/IMGT/HLA; März 2020). Der HLA-G-Locus besteht aus 8 Exons und 7 Introns, ähnlich wie andere klassische HLA-I-Loci (Amodio, 2020). Exon 1 kodiert das Signalpeptid. Die Exons 2, 3, 4 kodie- ren jeweils die extrazelluläre α1-, α2- und α3-Domäne. Die transmembrane und zyto- plasmatische Domäne der schweren Kette werden durch Exon 5 und 6 kodiert. In der Gegenüberstellung mit den klassischen Gruppe-I-Molekülen hat HLA-G eine kurze zytoplasmatische Domäne, angesichts eines vorzeitigen Stop-Codon in Exon 6. Exon 7 ist deshalb nicht in der reifen mRNA vorhanden. Exon 8 wird nicht gelesen, da es eine untranslatierte Region (3’UTR) besitzt (Carosella, 2008). Durch alternatives Splicing der primären mRNA können bekanntermaßen sieben HLA-G-Transkripte entstehen, vier membrangebundene Isoformenen (HLA-G1 bis - G4) und drei lösliche (engl. soluble, s) Isoformen (sHLA-G5 bis -G7). primäre mRNA Abbildung 6: Grafische Darstellung der mRNA-Splicevarianten und der Protein- isoformen von HLA-G (Würfel, 2019) 19
Theorie G1 und G5 repräsentieren die komplette Extrazellular-Protein-Struktur, bestehend aus den Alpha-Domänen (α1 - α3) und dem Beta-2-Mikroglobulin (ß2m), das auf Chromosom 15 kodiert ist. Andere Isoformen sind nicht mit ß2m verbunden und wei- sen unterschiedliche Proteinstrukturen auf. Die löslichen HLA-G-Isoformen zeigen eine hohe strukturelle Ähnlichkeit mit den membrangebundenen Varianten HLA-G1 und -G2 (Li, 2021). Das sHLA-G7 enthält nur die α1-Domäne und ähnelt deshalb der membranständigen HLA-G3-Proteinisoform (Zhao, 2012). Die genetischen und struk- turellen Ähnlichkeiten zwischen klassischen HLA-Klasse-I-Genen und HLA-G bzw. der weiteren HLA-G-Peptide und -Proteine werden durch posttranskriptionelle Modifi- kation generiert (Rodríguez, 2012; Ishitani, 2003). Es wird angenommen, dass sie einen Einfluss auf die T-Zell-Antwort haben. HLA-G kann zusätzlich in verschiedenen Proteinkomplexen vorkommen. Genauso wie die klassischen HLA-Proteine existiert HLA-G1 als Monomer, gebunden an ß2m. HLA-G1 und -5 können Heterodimere bil- den, aber auch Homotrimere durch Ausbildung von Disulfidbrücken zwischen den α1- Domänen. Die verbleibenden membrangebundenen Isoformen bilden keinen Kom- plex mit ß2m. Sie können aber Hetero- und Homodimere bilden, wie HLA-G1 und -G5. Die HLA-G-Expression ist auf die Trophoblastzellen beschränkt. Der Hauptvertreter HLA-G1 wird in extravillösen Trophoblastzellen (EVTs), wie z. B. in den endothelialen EVTs, den interstitiellen EVTs und den villösen Zellen des Trophoblasten exprimiert (HoWangYin, 2012). Die EVTs exprimieren außerdem die Isoformen HLA-G2, -G5 und -G6. Dagegen exprimieren die villösen Trophoblastzellen nur das lösliche Homo- log von HLA-G1, nämlich HLA-G5, das in den mütterlichen Blutkreislauf freigesetzt wird (Würfel F. M., 2019). HLA-G hat eine hemmende Wirkung auf NK-Zellen und Lymphozyten, die solublen Isoformen 4-7 wirken also immunsuppressiv. HLA-G interagiert mit Rezeptoren von Zellen des adaptiven und angeborenen Immunsystems. Die Expression nimmt nach der 12. Schwangerschaftswoche (SSW) ab und ist in der 16. bis 18. SSW am ge- ringsten (Würfel, Würfel, & Seelbach-Göbel, 2019). 20
Theorie 2.3 Der Sonderfall: Die klassische HLA-Gruppe C (Klasse Ia) Das väterlicherseits vererbte fremde Gen HLA-C ist die einzige HLA-Gruppe aus dem HLA-Komplex A bis DR des Embryos, die auf dem Trophoblasten bzw. der Pla- zenta exprimiert wird und aktivierend auf die uNK-Zellen wirkt. Das Antigen agiert nur mit einigen aktivierenden KIR (Killer-immunglobulinähnlichen Rezeptor), wodurch Zytokine und Wachstumsfaktoren von den uterinen Natürlichen Killerzellen (uNK) zur Förderung des zellulären Wachstums des Embryos und des Trophoblasten freige- setzt werden. Stimmen die HLA-C-Merkmale der Partner weitestgehend überein oder sind die paternalen HLA-C-Gruppen nur schwach wirksam, werden die NK-Zellen nur geringfügig aktiviert. Zusammenfassend entfaltet das Immunsystem der Mutter via HLA-C seine wichtige unterstützende Funktionen, da die Entwicklungsfähigkeit von Säugetierembryonen allgemein eher als schlecht bewertet wird und sie sehr empfind- lich auf äußere Einflüssen reagieren (Würfel, Würfel, & Seelbach-Göbel, 2019). 2.4 Bedeutung einzelner immunkompetenter Zellen 2.4.1 Natürliche Killerzellen (NK) Die NK gehören zum angeborenen Immunsystem und bilden die Brücke zum adapti- ven Immunsystem über die Ausschüttung verschiedener Immunmediatoren. NK- Zellen besitzen keine antigenspezifischen Rezeptoren, dafür aber inhibierende und aktivierende Rezeptoren sowie Mustererkennungsrezeptoren (Pattern Recognition Receptors, PRRs, z.B. CD56) (Doebis, 2019). Zu den Rezeptoren mit hemmender Wirkung gehören die Killer-immunglobulinähnlichen Rezeptoren (KIR, D-Rezeptoren) und die Lymphozyten-immunglobulinähnlichen Rezeptoren (LILR) oder die Lympho- zyten-immunglobulinähnlichen Transkripte (ILT) (Würfel, Würfel, & Seelbach-Göbel, 2019). ILT-2 kann z.B. an HLA-G koppeln. Die NK-Rezeptoren der Gruppe 2 (NKG/NKG2) oder die Killer-lezithinähnlichen Rezeptoren (KLR) gehören zu den ak- tivierenden Rezeptoren, der HLA-E erkennt. Die inhibierenden Rezeptoren binden an MHC-I-Moleküle (HLA-A, -B, -C) und erkennen somit körpereigene Zellen. Dadurch werden die NK-Zellen inhibiert, was der Selbsttoleranz dient. Die aktivierenden Re- zeptoren, die sog. NKRs (+S-Rezeptoren), interagieren mit stressinduzierten Ligan- den auf Zellen (z.B. Tumorzellen, virusbefallenen Zellen). Eine folgende Aktivierung der NK-Zellen führt zur Mobilisierung lytischer Granula und zur Produktion von pro- 21
Theorie entzündlichen Zytokinen (IFN-γ, TNF-α) sowie dem Granulozyten-Makrophagen- Kolonie-stimulierenden Faktor (GM-CSF) und anderer Chemokine. Zytotoxische Granula lösen in der Zielzelle die Apoptose aus, wodurch z.B. eine entartete Zelle zerstört wird (Doebis, 2019). Die Granula der NKs enthält außerdem Wachstumsfaktoren und Zytokine, die durch die Aktivierung synthetisiert und freigesetzt werden, wodurch der Embryo zu Beginn seiner Einnistung und Entwicklung einen wichtigen Booster für Wachstum und Diffe- renzierung erfährt (Würfel, Würfel, & Seelbach-Göbel, 2019). Nach der Aktivierung der NK kann eine zytotoxische als auch regulatorisch Reaktion ausgelöst werden (Kuon, 2018). NK kommen im peripheren Blut (sog. pNK CD56dim) und im uterinen Endometrium (sog. uNK CD56bright) vor. Der größte Teil der uNK (ca. 90%) zeichnen sich durch eine niedrige Zytotoxizität bei hoher Zytokinproduktion aus. Im Gegensatz dazu weisen die pNK eine hohe Zytotoxizität bei niedriger Zyto- kinproduktion auf. Eine erhöhte Zahl der uNK und v.a. der pNK wurde bei Fehlgebur- ten, Hypertonie, Präeklampsie und fetaler Wachstumsretardierung nachgewiesen (Kuon, 2018). Die uNK machen mit ca. 70 Prozent den hauptsächlichen Anteil der Immunzellen in der embryo-/fetomaternalen Grenzzone in der frühen Phase der Schwangerschaft aus (Kuon, 2018). Die Anzahl der uNK variiert im weiblichen Zyklus und in der Schwangerschaft. Sie spielen eine zentrale Rolle bei der Plazentation (Abbildung 7). Eine erhöhte uNK-Zellzahl führt zu einer Aggregatbildung um die Spiralaterien im Endometrium und beeinflusst dadurch den Umbau uteriner Spiralaterien vermutlich negativ (Kuon, 2018). Folglich kommt es durch die Neubildung von Blutgefäßen am Einnistungsort zu einem vermehrten Blutfluss, wodurch die Nidation beeinträchtigt werden kann. Bei Patientinnen mit RSA wurde eine signifikante Erhöhung der uNK- Zellzahl festgestellt (Kuon, 2018). 22
Theorie - Regulation der uterinen Spiralaterien - Schutz des Embryos vor Infektionen - Abschirmung väterlicher Antigene vor den Im- munzellen der Mutter - Regulation der Trophoblasteninvasion Abbildung 7: Funktion der uNK (Kuon, 2018) 2.4.2 Lymphozyten Die Aktivierung der Lymphozyten erfolgt über die Präsentation von fremden Antige- nen. Antigenpräsentierende Zellen (APC), wie Monozyten oder Vorstufen der B- und T-Lymphozyten, haben die Aufgabe fremde Antigene zu erkennen und dem Immun- system zu präsentieren (Würfel, Würfel, & Seelbach-Göbel, 2019). Dendritische Zel- len (DC) wie DC-10 repräsentieren die Mehrheit innerhalb der APC, der zweithäufigs- te Leukozytenpopulation in der Dezidua und vermitteln die Induktion der feto- maternalen Immuntoleranz in einem hohen Maße. Das Interleukin-10, ein immun- suppressives Zytokin, wird von DC-10-Zellen synthetisiert, die als einzige Untergrup- pe die HLA-G-Expression auslösen können (Xu D. P., 2016). DC sind ebenfalls in der Lage, Fremdantigene zu präsentieren und zudem eine spezifische Immunreaktion über B- und T-Lymphozyten auszulösen (Souza-Fonseca-Guimaraes, 2012). Unreife DC im Endometrium nehmen Antigene durch Phagozytose abgestorbener Zellen, Pinozytose von Molekülen oder rezeptorvermittelter Endozytose gezielt ausgewählter Substanzen auf (Würfel, Würfel, & Seelbach-Göbel, 2019). Gesteuert durch den Rei- fungsprozess der DC stoppt die Antigenaufnahme und die Antigenpräsentation auf der Zelloberfläche erfolgt über die Kopplung an Moleküle der HLA-Klasse-II (Bachy, 2008). Die DC wandern in das nächstgelegene lymphatische Organ (z.B. regionale Lymphknoten), wo sie auf B- und T-Lymphozyten treffen (Boddy, 2015). Es kommt zur Rekrutierung sog. regulatorischer T-Zellen (Treg) und ebenso der B-Zellen (Breg) (Kinder, 2014; Würfel, Würfel, & Seelbach-Göbel, 2019). Diese Zellen leisten einen wichtigen Beitrag zu einer gesteigerten Immuntoleranz hinsichtlich der vom Embryo präsentierten fremden Antigene (Betz, 2012). 23
Theorie Die Kommunikation zwischen APC, DC und Lymphozyten erfolgt über Rezeptor- Liganden-Systeme (sog. Immuno-Checkpoints) und einem Zusammenspiel aus Akti- vierung und Hemmung bezüglich der TH-1- und TH-2-Antwort, also der Ausschüttung von pro- oder antiinflammatorischer Zytokine (Würfel, Würfel, & Seelbach-Göbel, 2019). Mitte des II. Trimenons nimmt die HLA-F-Expression zu und somit auch die Population der Lymphozyten und ihre Bedeutung in der Regulation der Schwanger- schaft (Abbildung 12). 2.4.3 Makrophagen In der zweiten Zyklushälfte findet man Makrophagen in allen Schichten des Endomet- riums, d.h. im superfiziellen und luminalen Epithel sowie im Stroma (Würfel, Würfel, & Seelbach-Göbel, 2019). Zu ihren Aufgaben gehört es, eingedrungene pathogene Keime zu eliminieren und sich an der Desquamationsphase vor und während der Menstruation zu beteiligen (Thiruchelvam, 2013). Im Rahmen der Dezidualisierung und Nidation reichern sich Makrophagen (sog. Hofbaur-Zellen) in der Decidua basalis an (Würfel, Würfel, & Seelbach-Göbel, 2019). Ihre Konzentration in der Decidua pa- rietalis ist jedoch gering. Vermutlich ist ihre Hauptfunktion während der Einnistung und in der frühen Schwangerschaft, die Migration des villösen und extravillösen Tro- phoblasten sowie die Invasionstiefe vermittels des Makrophagen-Migrations- Inhibitionsfaktors (MMIF/MIF) zu steuern (Jovanović Krivokuća, 2021). MIF wird au- ßerdem von uNK-Zellen und trophoblastären Zellen exprimiert (Würfel, Würfel, & Seelbach-Göbel, 2019; Arcuri, 2001). Neben den DC vermitteln die dezidualen Makrophagen den Umbau der Spiralaterien im Endometrium und das Wachstum der Plazenta durch die Sekretion angiogener Wachstumsfaktoren, Matrix-Metalloproteasen (MMP) und Zytokine, darunter Interleu- kine und des Tumornekrosefaktors-α (TNF-α) (Würfel, Würfel, & Seelbach-Göbel, 2019). 24
Theorie 2.5 Die Schwangerschaft - ein Zusammenspiel von Fetus, Immun- zellen und HLA-Molekülen 2.5.1 Die Rolle des HLA-Systems in der Schwangerschaft Die Bedingungen während der Implantation und in der Schwangerschaft verändern sich stetig. Der Embryo bzw. Fetus hat nur über den Trophoblasten bzw. der Plazen- ta direkten Kontakt mit dem maternalen Immunsystem. Die trophoblastischen Zellen haben zwar die gleiche ursprüngliche Herkunft wie die anderen embryonalen Zellen, weisen dennoch nicht die originale adulte HLA-Signatur des Embryos/Fetus auf. Es werden außer HLA-C keine klassischen HLA-I- und -II-Proteine, wie HLA-A, -B, -DP, -DS exprimiert, stattdessen werden die nicht-klassischen HLA-Gruppen der Klasse Ib, wie HLA-E, -F, -G präsentiert (Abbildung 8). Die nicht-klassischen HLA-Gruppen unterdrücken immunkompetente Zellen des maternalen Immunsystems durch die Interaktion mit spezifischen Rezeptoren der Natürlichen Killerzellen und der Lympho- zyten. Bei gestörter Schwangerschaft bzw. Plazentalstörungen aufgrund untypischer Expression der nicht-klassischen HLA-Gruppen läuft dieser Mechanismus nicht ord- nungsgemäß ab. Tumorzellen nutzen oft den gleichen Funktionsweg, um das Im- munsystem zu umgehen. In diesen Zusammenhang ist es von großem Interesse, die nicht-klassischen HLA-Gruppen und ihre Interaktion mit Rezeptoren und Rezep- torfamilien der verschiedenen immunkompetenten Zellen zu untersuchen (Würfel F. M., 2019). HLA-C Abbildung 8: Die Plazenta mit ihrer HLA-Signatur als Pufferzone zwischen embryona- lem und maternalem Gewebe Quelle: Vorlage aus dem Buch „Schwangerschaft mit 40 Plus“ (Würfel, Würfel, & Seelbach-Göbel, 2019) 25
Theorie HLA-G und -E sind die Vertreter der HLA-Klasse-Ib, die hauptsächlich während der Einnistung und im ersten bis zweiten Trimenon exprimiert werden, um sich mit Zellen des angeborenen Immunsystems zu verständigen. Die HLA-Gruppen -E bis -G interagieren dabei mit bestimmten Rezeptoren der NK- Zellen (KIR, NKG/NKG2, KLR) und Lymphozyten (LILR / ILT). Die rezeptorvermittelte Interaktion spielt eine wichtige Rolle bei der Bildung der Plazenta und der Einstellung eines immunsupressiven Milieus zur Erhaltung der Schwangerschaft (Würfel, Würfel, & Seelbach-Göbel, 2019). Die relevanten nicht-klassischen HLA-Klasse-Ib-Gruppen wirken rezeptorvermittelnd inhibierend auf die uNK und andere Immunzellen des Endometriums, wodurch eine Abstoßungsreaktion gegen den genetisch und immunologisch fremden Embryo aus- bleibt. 2.5.2 Immunologische Interaktion an der embryo-maternalen Grenzfläche Der semiallogene Trophoblast bildet je nach Gestationsalter und anatomischer Lage mehrere Schnittstellen zum mütterlichen Immunsystem (Abbildung 9). Im frühen Sta- dium der Implantation ist die Blastozyste von Trophektoderm umgeben, das sich so- wohl zur endgültigen Zottenplazenta (villösen Plazenta), bestehend aus Zytotro- phoblasten (CT) und Syncytiotrophoblasten (ST), als auch zu dem eindringenden extravillösen Trophoblasten (EVT) entwickelt, der für den Austausch der Gase und die Bereitstellung von Nährstoffen über den mütterlichen Blutkreislauf verantwortlich ist (Pollheimer, 2018). Die früheste Schnittstelle (Nummer 1) umfasst den ST des sich implantierenden Embryos und die Dezidua. Das ist das einzige Mal, dass der ST an der Gewebegrenzfläche anliegt. Die Chorionzotten werden direkt von Sekret der Uterusdrüsen umspült, was die histotrophische Ernährung ermöglicht bis sich der intervillöse Kreislauf nach 8 Wochen durchgesetzt hat. Im zweiten Trimester ersetzen drei neue Schnittstellen die Erstgenannte: Schnittstelle Nummer 2 - zwischen ein- dringenden EVT und mütterlichen Immunzellen in der Dezidua, Schnittstelle Nummer 3 - zwischen Decidua parietalis und Amnionmembran und Schnittstelle Nummer 4 - zwischen ST und mütterlichen Blut. Der Syncytiotrophoblast gibt kontinuierlich extrazelluläre Vesikel (EV) in den intervil- lösen Raum und somit in den Blutkreislauf der Mutter ab (Levine, 2020; Giacomini, 2017). Die EVs sind membrangebundende, von Zellen stammende Partikel und spie- 26
Sie können auch lesen

















































