Einflussfaktoren der ex vivo-Migration neutrophiler Granulozyten
←
→
Transkription von Seiteninhalten
Wenn Ihr Browser die Seite nicht korrekt rendert, bitte, lesen Sie den Inhalt der Seite unten
Aus der Klinik für Kinder- und Jugendmedizin
der Universität zu Lübeck
Direktor: Prof. Dr. E. Herting
_______________________________________________________
Einflussfaktoren der ex vivo-Migration
neutrophiler Granulozyten
Inauguraldissertation
Zur Erlangung der Doktorwürde der Universität zu Lübeck
- Aus der Sektion Medizin -
Vorgelegt von
Gyde Anna Simone Nissen
aus Hamburg
Lübeck 2019
1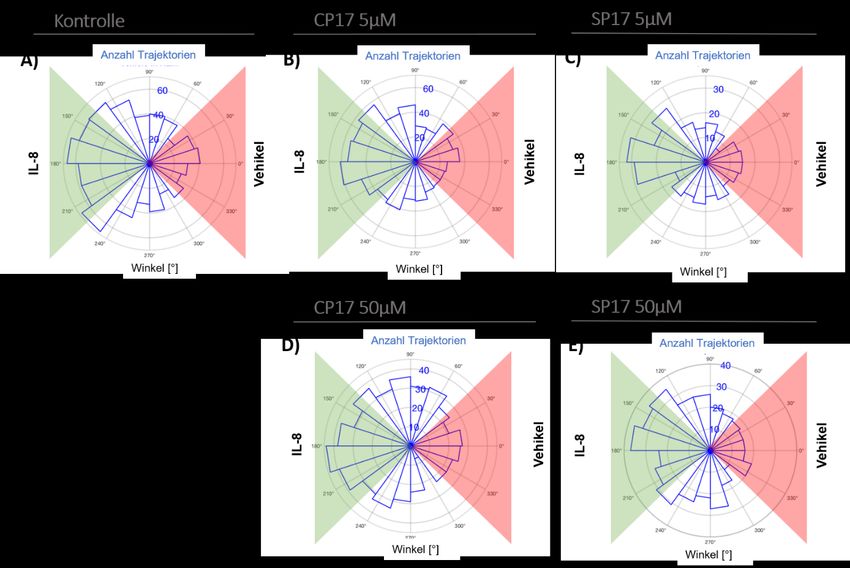
Erklärung
Ich versichere, die vorliegende Arbeit selbstständig und nur unter Verwendung der angegebenen
Hilfsmittel angefertigt zu haben.
Lübeck, 26.02.2019
_____________________________
Gyde Nissen
2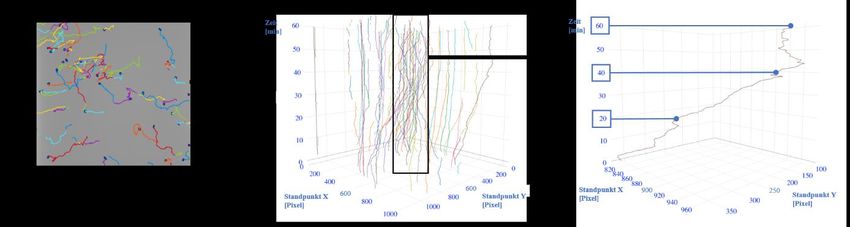
1. Berichterstatter: Prof. Dr. med. Matthias Kopp
2. Berichterstatter/In: Priv.-Doz. Dr. med. Lutz Welker
Tag der mündlichen Prüfung: 26.02.2019
Zum Druck genehmigt: Lübeck, den 26.02.2019
- Promotionskommission der Sektion Medizin -
3
Inhaltsverzeichnis
Abbildungsverzeichnis ................................................................................................................ 7
Tabellenverzeichnis ..................................................................................................................... 9
Abkürzungsverzeichnis ............................................................................................................. 10
Einführung................................................................................................................................. 12
1.1 Neutrophile Granulozyten ................................................................................................ 12
1.2 Migration neutrophiler Granulozyten ............................................................................... 12
1.2.1 Extravasation ............................................................................................................ 12
1.2.2 Migration im Interstitium .......................................................................................... 14
1.3 Einflussfaktoren neutrophiler Migration ........................................................................... 16
1.3.1 Chemokine ................................................................................................................ 16
1.3.2 Extrazelluläre Matrix ................................................................................................. 17
1.3.3 Matrikine .................................................................................................................. 18
1.3.4 Glukokortikoide ......................................................................................................... 20
1.4 Messinstrumente zellulärer Migration .............................................................................. 22
1.5 Fragestellung .................................................................................................................... 24
2 Material und Methoden................................................................................................... 26
2.1 Material ............................................................................................................................ 26
2.1.1 Puffer und Medien .................................................................................................... 26
2.2 Probandenrekrutierung und Probengewinnung ............................................................... 26
2.3 Isolation neutrophiler Granulozyten ................................................................................. 27
2.3.1 Bestimmung der Zellkonzentration ........................................................................... 28
2.3.2 Überprüfung der Reinheit und Zellvitalität ............................................................... 28
2.3.3 Überprüfung des Aktivierungsgrads ......................................................................... 29
2.4 Der µSlide-Migrationsassay .............................................................................................. 30
2.4.1 Vorbereitung der Matrix ........................................................................................... 30
2.4.2 Vorbereitung des Chemokins .................................................................................... 31
2.4.3 Vorbereitung der Wirkstoffe ..................................................................................... 31
2.4.4 Durchführung............................................................................................................ 32
2.4.5 Auswertung............................................................................................................... 34
2.4.6 A priori Definitionen zur Detektion technisch fehlerhafter Läufe ............................. 36
3 Ergebnisse ......................................................................................................................... 37
3.1 Analyse der Migration neutrophiler Granulozyten mit Timelapse-Mikroskopie und
Einzelzelltracking ......................................................................................................................... 37
3.1.1 Trajektoriendetektion und -verifikation ..................................................................... 37
3.1.2 Sektorenanalyse anhand des Migrationswinkels ...................................................... 37
4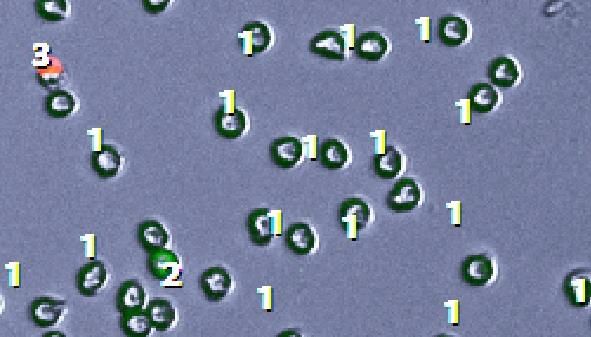
3.1.3 Qualitätskontrolle des µSlide-Migrationsassays ........................................................ 39
3.2 Neutrophile Migration in einfacher und komplexer extrazellulärer Matrix ....................... 39
3.2.1 Spenderabhängige Variabilität der PMN zur Testung von Auswirkungen durch Matrix
und Prednisolon ....................................................................................................................... 39
3.2.2 Migration in Fibronektin: Fokussierung im IL8-zentrierten Sektor ............................ 40
3.2.3 Migration in komplexer Plazentamatrix: Orientierung zum IL8-gerichteten Sektor .. 41
3.2.4 Einfache vs. komplexe Matrix: Abnahme IL8-fokussierter Migration, Geschwindigkeit
und Direktivität bei Zunahme IL8-neutraler Zellbewegungen in komplexer Matrix ................. 42
3.3 Veränderungen neutrophiler Migration durch Prednisolon .............................................. 43
3.3.1 Prednisolon in einfacher Matrix: Umkehr der Migrationsrichtung ........................... 43
3.3.2 Prednisolon in komplexer Matrix: Verminderte IL8-Fokussierung ............................ 44
3.4 Veränderungen neutrophiler Migration durch das Kollagen IV-Peptid CP17 .................... 45
3.4.1 Spenderabhängige Variabilität der PMN zur Testung von CP17 ................................ 45
3.4.2 CP17 in einfacher Matrix: Fokussierung im IL8-gerichteten Sektor mit Reduktion von
Geschwindigkeit und Direktivität ............................................................................................. 47
3.4.3 CP17 in komplexer Matrix: Defokussierung bei Abnahme der Geschwindigkeit ....... 49
4 Diskussion ......................................................................................................................... 51
4.1 Migration in einfacher Matrix ........................................................................................... 51
4.2 Migration in komplexer Matrix ......................................................................................... 53
4.3 Chemorepulsion durch Prednisolon.................................................................................. 55
4.4 Antiinflammatorisches Potential des Kollagen IV-Peptids CP17 ........................................ 58
4.5 Kritische Betrachtung des µSlide-Migrationsassays .......................................................... 61
4.6 Klinischer Ausblick zu neutrophiler Migration .................................................................. 62
5 Zusammenfassung............................................................................................................ 66
5 Literaturverzeichnis .......................................................................................................... 67
7 Anhang .............................................................................................................................. 78
7.1 Materialverzeichnis ........................................................................................................... 78
7.2 Ethikvotum ....................................................................................................................... 80
7.3 Probandenfragebogen ...................................................................................................... 81
7.4 Spenderinformationen...................................................................................................... 82
7.5 Abbildungen...................................................................................................................... 84
7.5.1 Differenzierung sich kreuzender Zellen ..................................................................... 84
7.5.2 Gegenüberstellung veröffentlichter und ergänzter Graphen .................................... 85
7.5.3 Geschwindigkeit und Direktivität unter Prednisolon in einfacher und komplexer Matrix
................................................................................................................................ 88
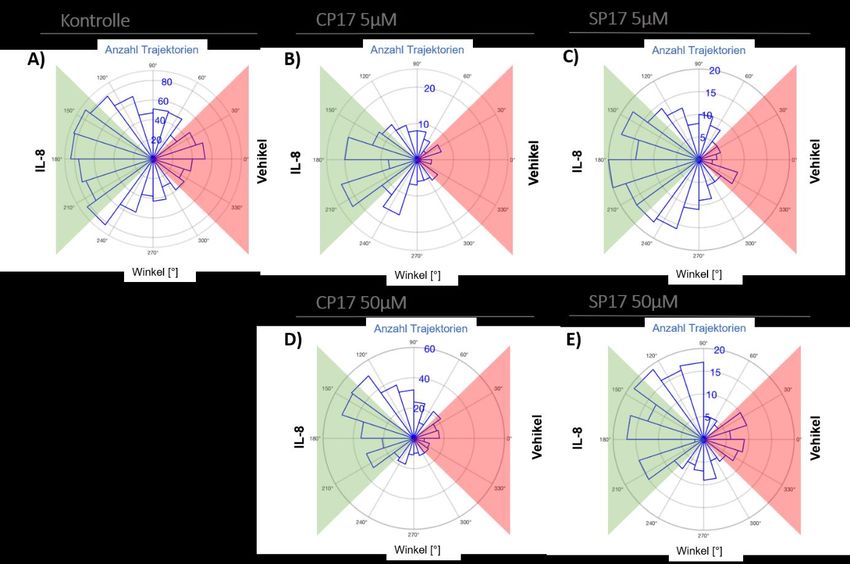
7.5.4 Winkelverteilung unter CP17/SP17 in komplexer Matrix .......................................... 89
7.5.5 Sektorenanalyse nach CP17/SP17 in komplexer Matrix ............................................ 89
Danksagung ............................................................................................................................... 90
5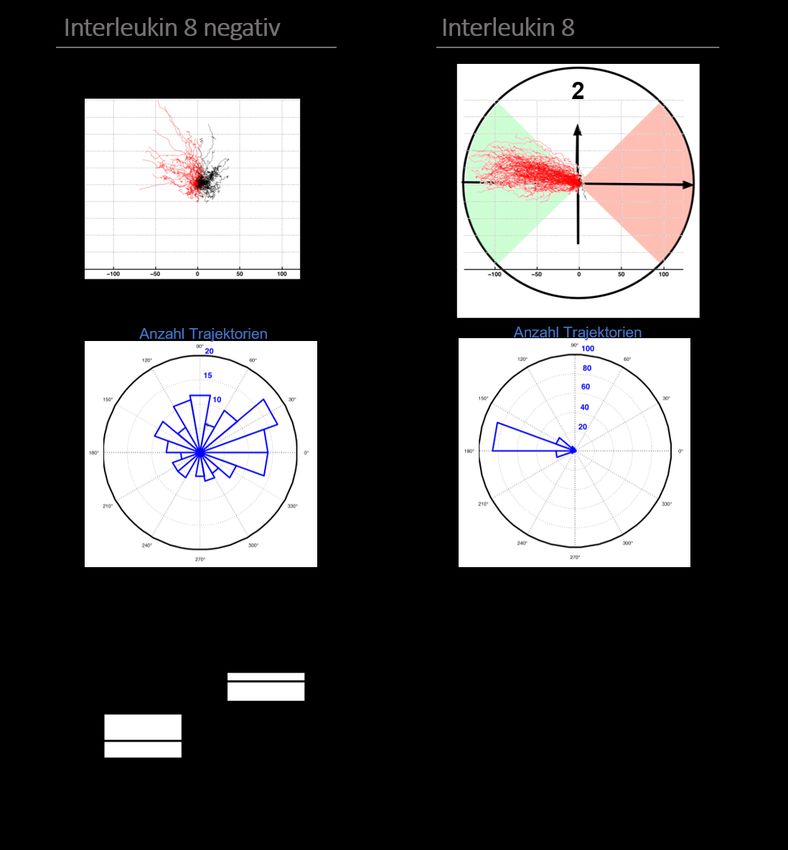
Curriculum vitae ....................................................................................................................... 91
6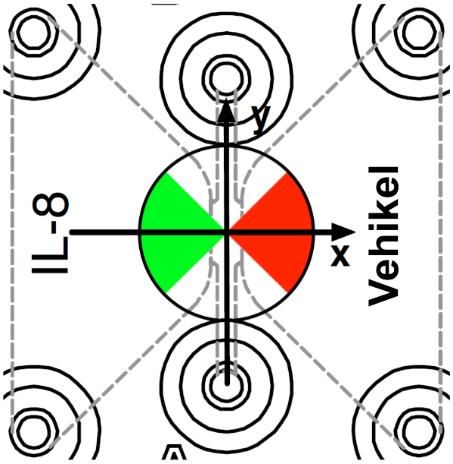
Abbildungsverzeichnis
Abbildung 1: Ablauf der Isolation von PMN mittels Dichtegradientenzentrifugation. .................... 27
Abbildung 2: Propidiumiodidfärbung des Isolats zur Bestimmung von Reinheits- und Vitalitätsgrad.
.......................................................................................................................................................... 29
Abbildung 3: Anti-CD62L-Färbung zur Bestimmung des Aktivierungsgrads. ................................... 30
Abbildung 4: Aufbaus eines µSlides Chemotaxis 3D (Ibidi GmbH, Deutschland). ........................... 33
Abbildung 5: Visualisierung von Zelldetektion und -tracking. .......................................................... 34
Abbildung 6: Einteilung der Migrationswinkel in Sektoren. ............................................................. 35
Abbildung 7: Technische Fehler einer Timelapse-Mikroskopie. ....................................................... 36
Abbildung 8: Migration in Ab- bzw. Anwesenheit von Interleukin 8................................................ 38
Abbildung 9: Spenderspezifische Direktivität und Geschwindigkeit der Migrationen von PMN der
Spender A1-6. ................................................................................................................................... 39
Abbildung 10: Spenderspezifische Direktivität und Geschwindigkeit der Migrationen von PMN der
Spender B1-7. ................................................................................................................................... 40
Abbildung 11: Sektorenanalyse der Migrationen unter IL8-Stimulation in FN. ............................... 40
Abbildung 12: Sektorenanalyse der Migrationen unter IL8-Stimulation in HEM. ............................ 41
Abbildung 13: Vergleich von Migrationen in einfacher und komplexer Matrix. .............................. 42
Abbildung 14: Sektorenanalyse der Migrationen nach Präinkubation mit Prednisolut® in FN. ...... 43
Abbildung 15: Sektorenanalyse der Migration nach Inkubation mit Prednisolut® in HEM. ............ 44
Abbildung 16: Spenderspezifische Direktivität und Geschwindigkeit der Migrationen von PMN der
Spender C1-4, D1-3 in FN bzw. E1-6 und F1-3 HEM. ........................................................................ 46
Abbildung 17: Migrationswinkelverteilung der Migrationen unter CP17/SP17 in FN...................... 47
Abbildung 18: Sektorenanalyse der Migrationen unter CP17/SP17 in FN. ...................................... 48
Abbildung 19: Änderung von Geschwindigkeit und Direktivität der Migrationen unter CP17/SP17 in
FN. .................................................................................................................................................... 49
Abbildung 20: Änderung von Geschwindigkeit und Direktivität unter CP17/SP17 in HEM. ............ 50
Abbildung 21: Fehlerhafte Zelldetektion bei Kreuzung zweier Zellen.............................................. 84
Abbildung 22: Gegenüberstellung veröffentlichter und ergänzter Graphen anhand der
7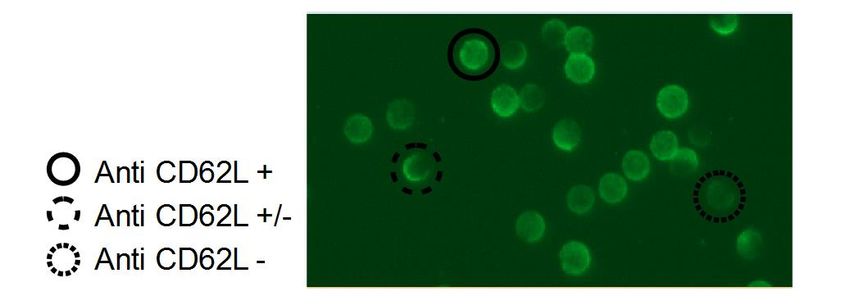
spenderspezifischen Direktivität und Geschwindigkeit der Migrationen in FN. .............................. 85
Abbildung 23: Gegenüberstellung veröffentlichter und ergänzter Graphen anhand der
spenderspezifischen Direktivität und Geschwindigkeit der Migrationen in HEM. ........................... 86
Abbildung 24: Sektorenanalyse der Migrationen in FN mit Gegenüberstellung publizierter und
korrigierter Daten. ............................................................................................................................ 87
Abbildung 25: Sektorspezifische Geschwindigkeit und Direktivität in FN und HEM nach
Präinkubation mit Predisolut®.......................................................................................................... 88
Abbildung 26: Migrationswinkelverteilung der Migrationen in HEM unter CP17/SP17. ................. 89
Abbildung 27: Sektorenanalyse der Migrationen unter CP17/SP17 in HEM.................................... 89
8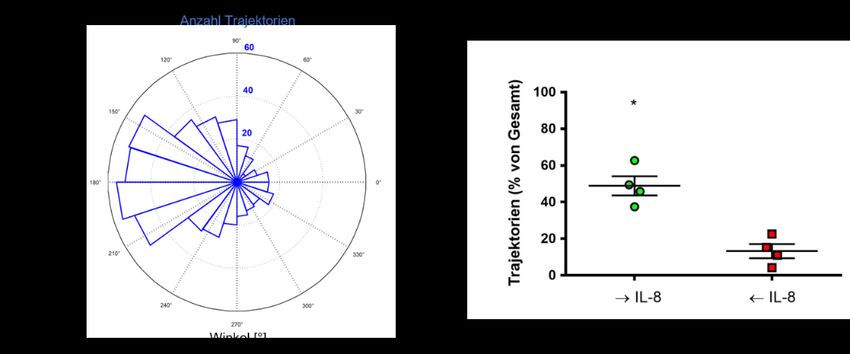
Tabellenverzeichnis
Tabelle 1: Wichtige Adhäsionsmoleküle der Rekrutierung von PMN in der Lunge und postkapillären
Venolen............................................................................................................................................. 13
Tabelle 2: Spenderdaten für Migrationsversuche in einfacher Matrix +/- Prednisolut®.................... 82
Tabelle 3: Spenderdaten für Migrationsversuche in komplexer Matrix +/- Prednisolut®. ................. 82
Tabelle 4: Spenderdaten für Migrationsversuche in einfacher Matrix +/- CP17 bzw. SP17. .............. 83
Tabelle 5: Spenderdaten für Migrationsversuche in komplexer Matrix +/- CP17 bzw. SP17. ............ 83
9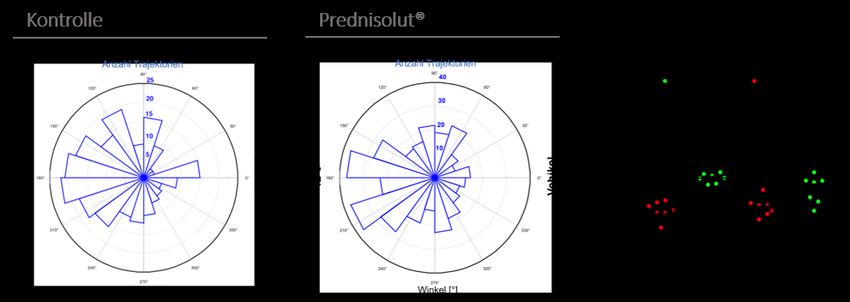
Abkürzungsverzeichnis
Abk. Abkürzung
BAL Bronchoalveoläre Lavage
Bzw. Beziehungsweise
°C Grad Celsius
COPD Chronisch obstruktive Lungenerkrankung, engl. chronic obstructive lung disease
C5a Komplementfaktor 5a
DAMP Damage associated molecular pattern
DNA Desoxyribonukleinsäure
ECM Extrazelluläre Matrix, engl. extracellular matrix
Engl. Englisch
fMLP Formyl-Methionyl-Leucyl-Phenylalanin
FN Fibronektin
GC Glukokortikoid, engl. glucocorticoid
GCR Glukokortikoidrezeptor, engl. glucocorticoid receptor
Geschl. Geschlecht
GFP Grün fluoreszierendes Protein
GM-CSF Granulocyte-monocyte colony-stimulating factor
Gr. Griechisch
HEM Humane extrazelluläre Matrix aus Plazentagewebe
ICAM 1/2 Intercellular adhesion molecules 1/2
IL8 Interleukin 8
KG Körpergewicht
KL Körperlänge
LFA-1 Lymphocyte function-associated antigen 1
LTB4 Leukotrien B4
Mac-1 Macrophage Antigen 1
10MAPK Mitogen-aktivierte Proteinkinase
MMP Matrixmetalloprotease
MT1-MMP Membran-Typ 1 Matrixmetalloprotease
NET Neutrophil extracellular trap
PAF Platelet activating factor
PAMP Pathogen associated molecular pattern
PBMC Mononukleäre Zellen im Blut, engl. peripheral blood mononuclear cell
PBS Phosphat gepufferte Salzlösung, engl. phosphate buffered saline
PGP Prolin-Glycin-Prolin
PI Propidiumiodid
PI3K Phosphoinosid-3-Kinase
PMN Polymorphonukleäre Leukozyten
PRR Pathogen recognition receptor
PSGL 1 P-selectin glycoprotein ligand 1
RFP Rot fluoreszierendes Protein
RNA Ribonukleinsäure, engl. ribonucleic acid
rpm Umdrehungen pro Minute, engl. rounds per minute
rcf Relative Zentrifugalkraft, engl. relative centrifugal force
ROS Reaktive Sauerstoffspezies, engl. reactive oxygen species
TIMP Gewebeinhibitor von Metalloproteasen, engl. tissue inhibitor of
metalloproteases
VCAM 1 Vascular cell adhesion molecule 1
11Einführung
Der Mensch ist in seiner Umwelt in ständigem Kontakt zu Fremdstoffen und potentiell pathogenen
Erregern. Zum Schutz vor Infektionen hat sich daher ein komplexes Wächter- und Abwehrsystem
aus angeborener und adaptiver Immunantwort ausgebildet. Überwindet ein Erreger die
mechanischen und chemischen Barrieren unseres Körpers, werden zu seiner raschen Erkennung
und Elimination zunächst die humoralen und zellulären Komponenten des angeborenen
Immunsystems aktiviert.
1.1 Neutrophile Granulozyten
Neben den Zellen des mononukleären Phagozytensystems, natürlichen Killerzellen, basophilen und
eosinophilen Granulozyten machen neutrophile Granulozyten die essentielle zelluläre Komponente
des angeborenen Immunsystems aus. Neutrophile Granulozyten sind dabei mit 50-70% die größte
Fraktion der Leukozyten im peripheren Blut eines Erwachsenen. Aufgrund ihres charakteristisch
segmentierten Zellkerns werden sie mit eosinophilen und basophilen Granulozyten zusammen auch
als polymorphnukleäre Leukozyten (PMN) bezeichnet. PMN differenzieren aus einer myeloischen
Vorläuferzelle im Knochenmark. Etwa 95% der PMN des Körpers ruhen hier, um über die Blutbahn
als eine der ersten Zellen an den Ort der Infektion rekrutiert zu werden [1]. Ein zweiter Pool existiert
endothelgebunden in Milz, Leber und Lunge [2]. Aktuell wird von einer mittleren Lebenszeit von bis
zu 5,4 Tagen ausgegangen [3]. Die Hauptaufgabe von PMN ist die Elimination von Pathogenen durch
Phagozytose, die Produktion reaktiver Sauerstoffspezies (engl. reactive oxygen species, ROS) und
die Sezernierung sogenannter Neutrophil extracellular traps (NETs). Zusätzlich steuern PMN durch
die Produktion von Chemokinen die Etablierung und Auflösung von Entzündungsherden und haben
auf diese Weise ebenfalls eine immunregulatorische Funktion [4].
1.2 Migration neutrophiler Granulozyten
1.2.1 Extravasation
Im Falle einer Infektion erkennen ortständige Zellen wie Makrophagen, Mastzellen oder
dendritische Zellen über ihre Pattern Recognition Receptors (PRR) sogenannte damage bzw.
pathogen associated molecular patterns (DAMP/PAMP) zerstörter körpereigener bzw. -fremder
Strukturen [5]. Durch die konsekutive Freisetzung von Zytokinen kommt es zur lokalen Reaktion mit
Vasodilatation und Endothelaktivierung. Hierbei werden unter anderem Oberflächenrezeptoren
12wie P- und E-Selektine aus Weibel-Palade Körperchen in die Zellmembran der Endothelzellen
transloziert beziehungsweise neu synthetisiert. Frei zirkulierende PMN werden durch deren
Bindung mittels P-selectin glycoprotein ligand 1 (PSGL-1) eingefangen und beginnen die Phase des
Rollings, den ersten Schritt der Extravasation [6],[7]. Zusätzliche Bindungen zwischen intercellular
adhesion molecules 1 und 2 (ICAM-1, -2) der Endothelzellen und den lymphocyte function-
associated antigens 1 (LFA-1) der PMN führen schließlich zu einer ausreichenden Entschleunigung
der PMN mit Übergang in eine rollende Bewegung an der Endotheloberfläche [8]. Hierbei kommen
PMN in direkten Kontakt zu PAMPs, Chemokinen und Wachstumsfaktoren, die eine Aktivierung der
PMN bewirken [9]. Durch Konformationsänderungen der Oberflächenrezeptoren entsteht eine
insbesondere Integrin-basierte, feste Adhäsion der PMN an der Endotheloberfläche [7],[10].
Zeitgleich bewirkt die Aktivierung der PMN eine Zunahme der Bindungskapazität
membranständiger L-Selektine. Über Leukozyten-Leukozyten-Interaktion können auf diese Weise
zusätzlich PMN aus der Blutbahn abgefangen werden. Da ein kompletter Stillstand den weiteren
Transit hindern würde, kommt es nach Aktivierung der PMN zu einem schnellen Abwerfen von L-
Selektin (engl. shedding) [7],[11], [12],[13].
Für die Überquerung der Barriere aus Endothel, Basalmembran und Perizyten verfügen PMN über
die Fähigkeit der transzellulären und parazellulären Migration. Der parazelluläre Weg ist dabei
schneller und effizienter [7], [14], weswegen er vornehmlich gewählt [15], [16] und hier beschrieben
wird. Für den Prozess ist eine Vielzahl von Oberflächenrezeptoren notwendig, die in Tabelle 1 näher
beschrieben sind.
MOLEKÜL DER ENDOTHEL-OBERFLÄCHE MOLEKÜL DER PMN-OBERFLÄCHE
ROLLING P-Selektin (CD62P) PSGL-1 (CD162), PTX3
E-Selektin (CD62E) PSGL-1 (CD162), ESL-1, CD44
PSGL-1 (CD162), GlyCAM L-Selektin (CD62L)
ADHÄSION ICAM-1 (CD54), ICAM-2 (CD 102) LFA-1/Integrin αLβ2 (CD11a/CD18)
VCAM-1 (CD106) VLA-4/Integrin α4β1 (CD49d/CD29)
TRANSMIGRATION ICAM-1 (CD54) LFA-1/Integrin αLβ2 (CD11a/CD18)
ICAM-2
VCAM-1(CD 102)
(CD106) MAC-1/Integrin
VLA-4/Integrin αα4βM1β(CD49d/CD29)
2 (CD11b/CD18)
PECAM-1 (CD31) PECAM-1 (CD31)
JAM-A (CD321) LFA-1/Integrin αLβ2 (CD11a/CD18)
JAM-B (CD322)
VE-Cadherin (CD144) VLA-4/Integrin
Zwischen α4β1 (CD49d/CD29)
Endothelzellen
JAM-C (CD323)
Tabelle 1: Wichtige Adhäsionsmoleküle der Rekrutierung von PMN MAC-1/Integrin αMund
in der Lunge β2 (CD11b/CD18)
postkapillären
Venolen. CD Cluster of differentiation , ESL-1 E-selectin ligand 1, GlyCAM Glycosylation-dependent cell
adhesion molecule, ICAM-1 Intercellular adhesion molecule 1, JAM Junctional adhesion molecule, LFA-
1 Lymphocyte function-associated antigen 1, MAC-1 Macrophage 1 antigen, PECAM-1
Platelet/endothelial cell adhesion molecule 1, PSGL-1 P-Selectin glykoprotein ligand 1, VCAM-1
Vascular cell adhesion molecule 1, VE-Cadherin Vascular endothelial cadherin, VLA-4 Very late antigen
4 [5], [7].
13Auf der Suche nach dem Weg des geringsten Widerstands tasten adhärente PMN zunächst mit Hilfe
von Pseudopodien ihre Umgebung ab. In vitro finden sich derartige Orte vor allem an Kontaktstellen
dreier Endothelzellen, da diese weniger Junktionsproteine und eine geringere Ordnung aufweisen
[16], [17]. Mit Hilfe feiner Zellkernausstülpungen drängen sie endotheliale Aktinfilamente
auseinander, um diese zu passieren [18]. Zusätzlich sind PMN mit einer Vielzahl
matrixdegradierender Enzyme ausgestattet. Engstellen können somit durch Lyse lokaler Strukturen
überwunden werden. Für die Extravasation von PMN sind die Matrixmetalloproteasen 2 und 9
(MMP2, MMP9) relevant. Diese Kollagenasen werden im Rahmen der Aktivierung von PMN aus
ihren Granula an die Zelloberfläche transloziert [19]. Um die Extravasation möglichst effektiv und
gewebsschonend zu gestalten wählen PMN gezielt Regionen mit einer niedrigen Dichte
extrazellulärer Matrix, die u.a. arm an Kollagen IV sind [20], [21]. Zusätzlich besitzen PMN die
Fähigkeit einer ausgeprägten Nukleusdeformität. So konnten Wolf et al. anhand Modellen nicht-
degradierbarer Matrix zeigen, dass PMN Öffnungen von bis zu einem Zehntel Durchmesser ihres
Zellkerns passieren können [22]. Ist umliegendes Gewebe zu sehr verdichtet und die Kapazität der
Nukleusdeformität überschritten, nutzen PMN die Möglichkeit zur perizellulären Kollagenolyse, um
die weitere Passage sicherzustellen [22], [23]. Diese Orte entwickeln sich hierdurch im Verlauf zu
präferierten Austrittsstellen für weitere emigrierende Leukozyten [21]. Darüber hinaus
übernehmen MMPs die Modulierung von Oberflächenrezeptoren wie Integrinen und L-Selektin und
unterstützen die Formierung und Eliminierung chemotaktischer Gradienten, weswegen sie für die
Extravasation unabdingbar sind [24]–[26], reviewed in [27]. Nach Überquerung der Basalmembran
erfahren PMN schließlich weitere Führung durch Chemokinsezernierung der Kapillaren-
ummantelnden Perizyten. Diese werden in einem interzellulären Spalt überwunden [28], [29].
1.2.2 Migration im Interstitium
Aufgrund technischer Limitationen ist die Migration im extravasalen Raum bisher unzureichend
erklärt. Generell gilt es fibrilläres, v.a. Kollagen-haltiges, von zellreichem Gewebe zu unterscheiden.
Zellreiches Gewebe findet sich zum Beispiel in epithelialen Barrieren, lymphatischen Organen und
dem Gehirn. Fibrilläres Stroma ist unter anderem in der Lunge, Dermis oder Plazenta unterhalb
endothelialer und epithelialer Barrieren vorhanden. Im Gegensatz zur Extravasation verläuft die
Fortbewegung in fibrillärem Gewebe non-adhäsiv, das heißt unabhängig von fester
Rezeptorbindung mittels Integrinen [30]–[32]. Dieses Muster wird auch als Gleiten und Zwängen
(engl. flowing and squeezing) beschrieben. Hier beeinflusst die Umgebung maßgeblich die
Fortbewegung der Zelle. So konnte gezeigt werden, dass Leukoyten speziell Kollagenfibrillen als
Führungsschienen nutzen, um an diesen entlang zu gleiten [22], [33]. Unabhängig von der
Zelladhäsion ist so eine Polarisation und Polymerisation des Aktomyosinskeletts ausreichend, um
14eine schnelle, amöboid-artige Fortbewegung entlang eines Chemokingradientens zu gewährleisten
[31], [34]–[36]. Regionen hoher Matrixdichte werden von PMN registriert und wenn nötig unter
Verformung des Zellkerns überwunden [35]. Im Rahmen von Entzündungsreaktionen kommt es zu
strukturellen Veränderungen des Interstitiums, die mit einer Erhöhung der Matrixdichte
einhergehen. Zunächst führt das entzündlich bedingte Kapillarleck mit konsekutivem
Wassereinstrom ins Gewebe zu einer Volumenzunahme und Verdrillung von Kollagenfibrillen [35].
Etwas verzögert beginnt die Augmentation der Kollagensynthese durch Fibroblasten und glatte
Muskelzellen [37]. Untersuchungen mit intravitaler Multiphotonenmikroskopie an Mäuseohren
bestätigten die Hypothese aus in vitro-Versuchen, dass PMN gezielt Regionen niedriger
Kollagenfibrillendichte aufsuchen, um möglichst rasch ihre Destination zu erreichen [35], [38]. Mit
dieser Form der Fortbewegung unterscheiden sich PMN gänzlich von mesenchymalen Zellen. Diese
sind während der interstitiellen Migration durchgehend auf eine feste Bindung an Matrixproteine
angewiesen und unterliegen bei Verlust des Matrixkontakts sogar einem als Anoikis bezeichnetem
Zelltod (gr. für Heimatlosigkeit) [39]. Auf diese Weise erreichen PMN Geschwindigkeiten von
maximal 20µm/min, womit sie bis zu 100-fach schneller sind als Fibroblasten [40], [41]. Interessant
ist zudem, dass die interstitielle Migration im Gegensatz zur Extravasation nicht-proteolytisch
verläuft [38], [42], [43]. Dennoch besteht eine Abhängigkeit von der Aktivität PMN-eigener
Matrixmetalloproteasen ohne die unter anderem die Polarisation Neutrophiler gestört ist [35]. Ob
spezielle perivaskuläre Gewebestraßen entlang von Perizyten von einem Teil der PMN für eine
zügige Migration bevorzugt werden, bleibt zu klären [31].
In zellreicher Umgebung werden Integrine für die Rekrutierung weiterer Leukozyten und Formation
strukturierter Zellaggregate am Entzündungsherd benötigt [41]. Speziell Mac-1- und LFA-1-
defiziente PMN zeigten nach ungehinderter Migration in fibrillärem Gewebe einen Migrationsstop
am Übergang zum zellreichen Entzündungsherd [41], [44].
151.3 Einflussfaktoren neutrophiler Migration
Zahlreiche Umgebungssignale beeinflussen über Modulation von Aktivierungsgrad, Adhäsion und
Zytoskelettkonformation fortwährend die Migration von PMN. Eine Auswahl der Einflussfaktoren ist
im Folgenden beschrieben.
1.3.1 Chemokine
Chemokine sind eine Gruppe strukturell verwandter, körpereigener Signalproteine, die eine erhöhte
Motilität und gerichtete Zellbewegung auslösen. Aktuell sind etwa 50 dieser 8-12kDa großen
Proteine bekannt [45]. Abhängig von der Position ihrer endständigen Cysteinreste (C) werden sie in
vier Gruppen unterteilt: XC, CC, CXC und CX3C, wobei X jede andere Aminosäure beschreibt.
Chemokine werden von Immunzellen, aber auch Mesenchymzellen wie beispielsweise
Endothelzellen oder Pneumozyten, in den Extrazellularraum sezerniert. Dort liegen sie zunächst frei
oder an extrazelluläre Matrixkomponenten gebunden vor. Die Gruppe der CXC-Chemokine kann in
Abhängigkeit von der An- bzw. Abwesenheit eines den beiden ersten Cysteinen vorangestellten ELR-
Motivs (Glutamat-Leucin-Arginin) in zwei Subgruppen unterteilt werden. Die sogenannten CXC-ELR-
Liganden sind besonders relevant für die Attraktion von PMN [46]. Einen wichtigen Vertreter dieser
Gruppe stellt Interleukin 8 (IL8/CXCL2) dar. Neben dem Lipidmediator Leukotrien B4 (LTB4) , dem
Komplementfaktor 5a (C5a) und dem Bakterien-entstammenden Formylpeptid Formyl-Methionyl-
Leucyl-Phenylalanin (fMLP) ist IL8 einer der physiologisch wichtigsten Attraktoren Neutrophiler
Migration [31], [47]. IL8 wird unter anderem von Monozyten und Makrophagen, Endothel- und
Epithelzellen, glatten Muskelzellen und aktivierten PMN selber synthetisiert [48], [49]. Die
Produktion unterliegt transkriptorischer Regulation, weswegen die Sekretion im Vergleich zur
Freisetzung von Lipidmediatoren frühestens eine Stunde nach Stimulation beginnt, jedoch auch bis
zu 24 Stunden anhalten kann [49], [50]. In der Kaskade der Rekrutierung spielt IL8 dementsprechend
erst im späteren Verlauf einer Entzündungsreaktion eine Rolle [50]. Durch die Fähigkeit der Bindung
von Glykosaminoglykanen und Heparansulfaten sind Chemokine vor direkter Proteolyse geschützt,
was ihre Verweildauer im Gewebe erhöht [51]–[53]. Hauptaufgaben sind die Rekrutierung von PMN
an den Entzündungsort und die Mobilisierung weiterer PMN aus dem Knochenmark [19], [50].
Darüber hinaus initiiert IL8 über eine Aktivierung von gewebsständigen dendritischen Zellen die
Mobilisierung von T- und B-Lymphozyten und damit den Beginn der adaptiven Immunantwort [45].
Wahrgenommen wird IL8 über die Chemokinrezeptoren CXCR1 und CXCR2 an der Zelloberfläche
aktivierter PMN [54]. Beide stellen G-Protein-gekoppelte Rezeptoren dar, die über den
Phosphoinosit-3-Kinaseweg (PI3K) zu einer sich rasch amplifizierenden, intrazellulären Reaktion und
damit schnellen Umsetzung von Aktinpolymerisation in Migration führen [47], [55]. Um
16sicherzustellen, dass PMN in einem Potpourri aus Stimulanzien adäquat reagieren, sind diese
hierarchisch geordnet [56]. Die Chemoattraktantien C5a und fMLP sind dabei LTB4 und IL8 höher
gestellt und können über simultane Inhibition des PI3K-Signalwegs und Aktivierung der Mitogen-
aktivierten-Proteinkinase Kaskade (MAPK) die finale Zellrichtung beeinflussen [57]–[59]. Auch die
zeitliche Präsentation der Stimulantien scheint eine Rolle zu spielen. So zeigten Foxman et al., dass
PMN eine Art Aktivierungsgedächtnis besitzen und speziell auf neue Reize reagieren [58], [60].
1.3.2 Extrazelluläre Matrix
Der Begriff der extrazellulären Matrix (ECM) bezeichnet ein dreidimensionales Netzwerk, das unter
anderem Blut- und Lymphgefäße, Nerven und Drüsen in seine Umgebung einbettet und dem Organ
seine Struktur und Elastizität verleiht. Die verschiedenen Komponenten der ECM werden unter dem
Begriff des „Matrisoms“ zusammengefasst. Dabei stellen Glykoproteine, Kollagene und
Proteoglykane die Hauptbestandteile des Matrisoms dar, die sich in jeweils unterschiedlicher
Zusammensetzung in den Organen wiederfinden [61]–[63]. Darüber hinaus beinhaltet der Begriff
des Matrisoms inzwischen auch alle mit der ECM-interagierenden Proteine. Dazu gehören ECM-
Regulatoren, wie Proteasen und Verbindungsproteine und matrixgebundene Faktoren wie Zytokine.
Um Übersicht und Verständnis der ECM zu fördern gründeten Naba et al. 2016 das „Matrisom-
Projekt“. Der hieraus entstandene Matrix-Atlas umfasst in seiner aktuellen Version 1026 ECM-
codierende Gene [61]. Die ECM ist indes ein dynamischer Raum, der sich stetig neuen
Anforderungen anpasst [64]. Beispielsweise werden in der Lunge täglich etwa 10% des Kollagens
durch Mesenchymzellen restrukturiert oder abgebaut und neu synthetisiert [65], [66]. Dabei
entstehen unter anderem sogenannte „Super-Highways“ auf denen eine besonders effektive Form
der Zellmigration ermöglicht wird [33]. Weiterhin ist die ECM durch Präsentation von Chemokinen
und Wachstumsfaktoren und direkter Kraftübertragung mittels Zell-ECM Interaktion an der
Modulation von zellulärem Migrationsverhalten beteiligt [67], [68]. Mit der Fähigkeit der Bindung
von Migrationsmodulatoren hat ECM zudem eine Reservoirfunktion und kann bei Bedarf eine
Gradientenbildung für den Ablauf gerichteter Migration stabilisieren. Die Liste weiterer Funktionen
der ECM wächst stetig. Aktuelle Zusammenfassungen finden sich in [61], [69].
Ein in nahezu allen Organen exprimiertes Matrixmolekül ist das Glykoprotein Fibronektin (FN). Es
besteht aus zwei etwa 230kDa großen Untereinheiten, die über zwei Disulfidbrücken miteinander
verknüpft sind. Ein einziges Gen codiert für FN, welches durch Alternatives Splicing die zwei
Hauptformen, zelluläres und plasmatisches FN, ergibt [70]. Zelluläres FN wird beispielsweise von
Fibroblasten und Epithelzellen in unlöslicher Form sezerniert und an der Zelloberfläche bereits
dimerisiert direkt in die ECM eingebaut. Plasmatisches FN dahingegen ist löslich und wird von
17Hepatozyten in die Blutbahn freigesetzt von wo es in umliegendes Gewebe diffundiert und dort
eingebaut wird [71]. Plasmatisches FN ist besonders leicht für experimentelle Zwecke zu
extrahieren, was es für die Forschung zu einem beliebten und viel verwendeten Matrixmolekül
macht [72]. Zur Kommunikation mit seiner Umgebung besitzt FN verschiedene funktionelle
Abschnitte. Neben Domänen zur Verbindung mit anderen FN-Molekülen und Einheiten zur Bindung
von Matrixmolekülen, wie z.B. Kollagenen, dient die sogenannte RGD-Sequenz (Arginin-Glycin-
Asparaginsäure) der Herstellung von Integrin-vermitteltem Zellkontakt. Die Sequenz ist
charakteristisch für viele Glykoproteine der ECM und ist insbesondere in den ersten Schritten der
Matrix-Formierung mit Fibrillenbildung unabdingbar [69], [73], [74].
Die tatsächliche Zusammensetzung verschiedener Gewebearten umfasst eine große Zahl an
Mitspielern, deren Zusammenspiel aufgrund ihrer Komplexität noch nicht vollständig verstanden
ist. So konnten massenspektrometische Untersuchungen von dezellularisiertem, humanen
Lungengewebe eine Vielfalt von über 127 Matrisom-Proteinen in den unteren Atemwegen abbilden
[75]. Parallel dazu führten weitere Arbeitsgruppen Proteom-Analysen unterschiedlicher
Gewebeproben durch, die zusätzlich Aussagen zur Quantität der Einzelkomponenten ermöglichten
[69]. Exemplarisch zu erwähnen seien Analysen muriner Lungen, in denen Schiller et al. eine Vielfalt
von mindestens 150 Matrisom-Proteinen nachweisen konnten [76]. Kollagene machten dabei den
größten Anteil der Matrixproteine aus. Man geht davon aus, dass speziell die fibrillären Kollagene I,
II, III, V und XI zusammen mit Proteoglykanen das Grundgerüst der Lunge bilden. Dieses Interstitium
verbindet Parenchymzellen miteinander und verleiht dem Organ seine biomechanischen
Charakteristika [69], [77]. Darüber liegt eine dünne Schicht aus Glykoproteinen und
netzwerkartigem Kollagen IV, welche die Struktur von Basalmembranen bildet und so Epithel und
Endothel basalwärts begrenzt. Das Lungengerüst ist dabei durch die Interaktion von Matrix und
PMN in stetigem Wandel. So konnten Lämmermann et al. durch intravitale 2-Photonenmikroskopie
eindrucksvoll zeigen, wie PMN eine Neuordnung von Kollagenfibrillen vornehmen, um ein
kollagenfreies, inflammatorisches Zentrum zu generieren [41]. Welche Auswirkungen sowohl die
physiologische Gewebezusammensetzung als auch deren pathologische Umstrukturierung auf die
Interaktion von Zellen und Matrix hat gilt es weiter zu untersuchen.
1.3.3 Matrikine
Durch Speicherung und gezielte Präsentation von Zytokinen und Wachstumsfaktoren ist ECM ein
wichtiger Regulator von Zellverhalten. Darüber hinaus kann sie durch die Liberation sogenannter
Matrikine biologische Aktivität kontrollieren. Matrikine sind Fragmente extrazellulärer
Matrixproteine, die durch enzymatische oder chemische Spaltung von diesen abgetrennt werden
18und so eine von ihrem Mutterprotein unabhängige Wirkung auf Zellaktivität entfalten. [78]–[81].
Die Wirkung von Matrikinen ist vielseitig. Während einige Matrikine die Zellproliferation,
Angiogenese und Tumorentstehung beeinflussen, zeigen andere Matrixfragmente zytotoxische
oder migrationsmodulierende Wirkung [81], [82]. Freigesetzt werden Matrikine durch die Aktivität
Matrix-prozessierender Enzyme. Wichtige Vertreter sind Matrixmetalloproteasen (MMP) und die
Serinprotease Elastase, die von einem Großteil differenzierter Zellen exprimiert werden. PMN
besitzen in ihrem Zytosol verschiedene Arten von Vesikeln in denen MMPs und Elastasen jeweils
separat als Proenzym gespeichert und bei Aktivierung in die Zellmembran transloziert werden [19],
[83]. Dort sind sie fest verankert, weswegen sie ihre katalytische Wirkung speziell auf Proteine der
Zellmembran, sekretierte Proteine und Proteine im direkten extrazellulären Umfeld ausüben [84].
Reguliert wird die Aktivität der multifunktionellen Enzyme auf Ebene der Genexpression, durch
Angebot und Affinität von Substrat sowie einem Gleichgewicht aus Proenzym-Aktivierung und
Enzym-Inaktivierung [85]. Die Aktivierung erfolgt unter anderem durch Abspaltung der Pro-Domäne
des Zymogens durch weitere Proteasen, durch Bindung von Matrix oder durch oxidative Prozesse.
Sezernieren PMN reaktive Sauerstoffspezies können diese durch Oxidierung der Pro-Domäne MMPs
aktivieren und durch Modifikation von Aminosäuren der katalytischen Domäne das Enzym
schlussendlich inaktivieren. PMN besitzen damit ein Instrument zur impulsiven Kontrolle
proteolytischer Aktivität [86], [87]. Inaktiviert werden MMPs zudem zum Großteil durch
unselektives α2-Makroglobulin, ein Akute-Phase-Protein, welches von aktivierten Makrophagen in
die Blutbahn sezerniert systemisch schnell verfügbar ist [88]. Spezifische Inaktivierung von MMPs
erfolgt nicht zuletzt durch eine Gruppe von vier sogenannten Gewebeinhibitoren (engl. tissue
inhibitors of metalloproteases, TIMP) [89]. Physiologischerweise tritt ein erhöhter Matrixabbau im
Rahmen inflammatorischer Prozesse und Reparaturvorgänge auf, wobei das Vorkommen Matrix-
prozessierender Enzyme zwischen den einzelnen Gewebeformen variiert. Bei mehreren mit
Lungengerüstumbauten einhergehenden Erkrankungen wie dem Asthma bronchiale, der Zystischen
Fibrose oder der Bronchopulmonalen Dysplasie konnte ein Ungleichgewicht von MMPs und TIMPs
im Sinne eines pathologisch erhöhten MMP-Anteils nachgewiesen werden. Eine Störung der Matrix-
Homöostase wird daher als wichtiger Faktor ihrer Pathogenese angenommen [85], [90]–[93].
Mit einem Anteil von etwa 85% machen Kollagene das Gros unserer Proteinmasse aus und sind
damit eine wichtige Quelle von Matrikinen [62]. Insgesamt zählt man in Vertebraten mindestens 28
verschiedene Arten an Kollagenen, die sich aus je drei Polypeptidketten über
Wasserstoffbrückenbindungen zu einer alpha-Helix zusammensetzen [94]. Diese Tripelhelices
formen wiederum Strukturen höherer Ordnung, die fibrilläre von nicht-fibrillären Kollagenen
unterscheidet [95]. Während fibrilläre Kollagene vorwiegend im Interstitium zu finden sind, bilden
nicht-fibrilläre Kollagene natürliche Barrieren wie die Basalmembran [96]. Ein wesentlicher
19Bestandteil von Basalmembranen ist Kollagen IV, welches in sechs genetisch eindeutigen Isoformen
(α1-6) existiert [97]. Die Verteilung der Kollagen IV-Isoformen ist organspezifisch, wobei in der Lunge
alle Phänotypen vertreten sind [98], [99]. Jede Isoform besitzt eine distinkte Kopfgruppe, die als
nicht-kollagene (NC)1 Domäne bezeichnet wird. Ihre Freisetzung aus Basalmembranen scheint
durch verschiedene Mechanismen Einfluss auf Organentwicklung und die Entstehung onkologischer
Erkrankungen zu haben [63], [100], [101]. Ein bereits seit den 1990er Jahren untersuchtes Matrikin
ist Tumstatin, die NC1-Domäne der Kollagen IV-Isoform α3. Diesem wird eine Hemmung der
Angiogenese und Inhibition der Zellproliferation von Pankreas-, Magen- und Mammakarzinom-
sowie Melanom- und Fibrosarkomzellen zugesprochen [99], [102]–[104]. Zudem gibt es Hinweise
eines Effektes auf akute inflammatorische Reaktionen. Monboisse et al. isolierten eine 18
Aminosäuren-lange Sequenz aus Tumstatin (Aminosäure 185-203), welches eine Inhibition der
neutrophilen ROS-Produktion bewirkte [105]. Nur wenige Jahre später zeigten Ziaie et al., dass eine
Exposition zu selbigem Peptid eine artifizielle Basalmembran aus Kollagen I und Endothelzellen vor
Schaden durch fMLP-stimulierte PMN bewahrte [106]. Verantwortlich für die Abspaltung von
Tumstatin ist ein MMP2- und MMP9-involvierender Mechanismus [99], [104]. Beide stellen Enzyme
dar, die auch von PMN sezerniert werden. Ein weniger bekanntes Matrikin ist Lamstatin, die NC1-
Domäne der Kollagen IV Isoform-α5, die eine inhibierende Wirkung auf Lymphangiogenese ausübt.
Unter der Annahme einer gemeinsamen aktiven Region führten Weckmann et al. daher eine
Sequenzanalyse der beiden NC1-Domänen Tumstatin und Lamstatin durch und identifizierten einen
homologen Abschnitt, Aminosäure 66-82 von Lamstatin entsprechend. Das Peptid mit dem Namen
CP17 zeigte ebenfalls einen inhibitorischen Effekt auf Lymphangiogenese [107].
1.3.4 Glukokortikoide
Glukokortikoide (GC) bezeichnen eine Klasse von Steroidhormonen, die hauptsächlich im Cortex
unserer Nebennieren gebildet werden. Weitere Syntheseorte finden sich im Thymus, der Haut, im
Darm und Gehirn, wo GC durch auto- und parakrine Mechanismen die lokale Zellentwicklung und
Immunzellaktivierung beeinflussen [108]. Wie alle Steroidhormone werden GC in einem
mehrschrittigen Prozess aus Cholesterin, einem in Plasmamembranen vorkommenden Lipid,
gebildet. Der Hauptvertreter der menschlichen GC ist Cortison. Dessen Freisetzung beruht auf der
Stimulation durch die Hypothalamus-Hypophysen-Nebennierenrinden-Achse [109]. Natürliche
Auslöser sind unter anderem physischer und psychischer Stress sowie entzündliche Reaktionen. Als
lipophiles Molekül kann Cortison durch Zellmembranen diffundieren und so nach Freisetzung durch
Bindung des zytosolischen GC-Rezeptors seine Wirkung rasch ausüben. GC-Rezeptoren finden sich
ubiquitär in allen kernhaltigen Zellen als Monomer im Zytoplasma. So auch in Zellen von
Hypothalamus und Hypophyse, worüber die weitere GC-Produktion durch negatives Feedback
20reguliert wird. Die Wirkung von GC ist spezifisch für die jeweilige Zellart, was aktuell auf die Vielfalt
von Expressionsmustern verschiedener GC-Rezeptor-Isoformen zurückgeführt wird [110]. Bei
Bindung von Cortison dimerisiert der GC-Rezeptor und transloziert als Rezeptor-Ligand-Komplex in
den Nukleus, wo er durch Kontakt zu GC-empfindlichen Bereichen der DNA (engl. glucocorticoid
responsive elements) als Induktor oder Suppressor einer Vielzahl von Genen fungiert. Wesentliche
Effekte von GC sind zum einen die namengebende Regulation des Glukosestoffwechsels [111] und
zum anderen eine immunmodulatorische Wirkung. Deren Entdeckung im Rahmen der Behandlung
einer Patientin mit Rheumatoider Arthritis revolutionierte 1949 die Medizin [112]. GC werden
seither vielseitig therapeutisch angewendet, so auch in der Behandlung chronischer
Lungenerkrankungen wie dem Asthma bronchiale, der primären ziliären Dyskinesie und der
Zystischen Fibrose. Diese Erkrankungen sind durch eine markante neutrophile Inflammation geprägt
über die eine Korrelation mit der Schwere der Erkrankung berichtet wird [113]–[116]. Inzwischen
existiert eine Vielzahl synthetischer GC-Präparate, die durch verbesserte Bioverfügbarkeit und
Halbwertzeit die Anwendung erleichtern und aufgrund verstärkter Affinität zum Glukokortikoid- und
schwächerer Affinität zum Mineralokortikoid-Rezeptor ein geringeres Nebenwirkungspotential
aufweisen. Eines dieser Medikamente ist das oral nahezu vollständig resorbierte Prednisolon, was
gegenüber Cortison eine 4-fache glukokortikoide sowie 0.8-fache mineralokortikoide Wirksamkeit
entfaltet. Mit einer biologischen Halbwertzeit von bis zu 36 Stunden übersteigt Prednisolon die des
Cortisons etwa um das Dreifache [117]. Die immunmodulatorische Wirkung von GC beruht auf
verschiedenen Mechanismen. Wie Cain et al. zusammenfassen beginnt die antiinflammatorische
Wirkung von GC bereits in der frühen Alarmierungphase durch die Suppression intrazellulärer
Signalkaskaden abwärts von Pathogensensoren wie den PRRs. Durch die Suppression von
Adhäsionsrezeptoren wie ICAM1 und VCAM1 und Chemokinen wie IL8 hemmen GC zudem die
Mobilisierung von Leukozyten aus Blutbahn und naher Umgebung. Gleichwohl fördern GC auch die
Auflösung von Entzündungsherden indem die Phagozytose apoptotischer Zellen initiiert wird und
eine Sensibilisierung für anti-entzündliche Botenstoffe wie Lipoxin A4 erfolgt [118]. Überdies üben
GC eine akute Wirkung, die nicht über transkriptorische Veränderungen erklärt werden kann, aus.
Unter anderem wird eine Veränderung transmembranärer Kationenströme durch Interkalation von
GC in die Plasmamembran diskutiert. Andere vermuten hingegen die Bindung
zellmembranständiger GC-Rezeptoren oder eine direkte Interaktion von GC-Rezeptor und
zytoplasmatischen Signalproteinen als alternative Wirkwege [119]. Es ist bekannt, das GC durch eine
Förderung der Freisetzung von PMN aus dem Knochenmark und eine Inhibition der Apoptose von
PMN, die angeborene Immunreaktion stärken [120]. Gleichzeitig hemmen GC jedoch die Expression
von gewebedegradierenden Kollagenasen und elementaren Enzymen der Lipidmediatorsynthese,
was dem Erhalt einer akuten inflammatorischen Reaktion entgegenwirkt [121]. Die genaue
21Funktion von GC in der akuten neutrophilen Inflammation gilt es weiter zu evaluieren.
1.4 Messinstrumente zellulärer Migration
Die Fortbewegung von Zellen ist grundlegend für die Embryogenese und spätere Umstrukturierung
von Organen. Zudem ist sie unerlässlich für unsere Immunantwort und Wundheilung nach Trauma
oder Infektion. Ein umfassendes Verständnis der Migration neutrophiler Granulozyten und ihrer
Modulatoren ist für die Ergründung der Pathogenese von Erkrankungen mit ausgeprägter
neutrophiler Inflammation wie dem Asthma bronchiale, chronisch entzündlichen
Darmerkrankungen oder der rheumatoiden Arthritis fundamental [122]. Man unterscheidet
verschiedene Arten der Migration. Chemokinese bezeichnet eine zufällige, ungerichtete Bewegung
als Zellantwort auf einen Umweltreiz. Dahingegen beschreibt Chemotaxis eine gerichtete Migration
als Reaktion auf ansteigende Konzentrationen eines gelösten Stimulus und kann entsprechend der
Hauptmigrationsrichtung hin oder weg von einem Chemokin in Chemoattraktion und
Chemorepulsion unterteilt werden [123]. Die Entwicklung von Assays zur Untersuchung des
Migrationsverhaltens von Immunzellen hat in den letzten Jahren großen Fortschritt erfahren,
weswegen die gängigen Versuchsaufbauten hier vorgestellt werden sollen.
Eines der am längsten verwendeten in vitro-Systeme ist die sogenannte „Boyden-Kammer“. Sie
besteht aus einem unteren, Chemokin-haltigen Reservoir, das durch eine Membran von einer
oberen, Zell-haltigen Kammer getrennt ist. Nach einer definierten Inkubationszeit werden die Zellen
entweder in der Membran fixiert und ausgezählt oder ihr Anteil in der unteren Kammer, meist
photometrisch, bestimmt [124]. Mit der Zigmond- und der Dunn-Kammer wurde das Zwei-Kammer-
System weiterentwickelt. Chemokin- und Zellreservoir sind hier entweder nebeneinander oder
zirkulär umeinander angeordnet und über eine schmale Brücke miteinander verbunden [125],
[126]. Dieser Aufbau ermöglicht die Beobachtung des Migrationsprozesses an sich. Dies ist in der
Boyden-Kammer nicht möglich, die zwar aufgrund ihrer einfachen Handhabung, schnellen
Umsetzung, guten Reproduzierbarkeit und niedrigen Kosten gerne als Screeninginstrument
verwendet wird, jedoch einen Endpunktassay darstellt. Mit der traditionellen Boyden-Kammer ist
daher eine Differenzierung zwischen Chemokinese und Chemotaxis nicht möglich [127]. In der
zweiten Hauptgruppe der Migrationsversuche basierend auf Agar-Platten ist dies aufwändig, aber
prinzipiell machbar. Der wichtigste Vertreter dieser Gruppe ist der Unter-Agarose-Assay. Dieser
besteht in seinem Grundaufbau aus einer abgedeckten Agarosegel-Schicht, in die drei Löcher
gestanzt und je eines mit Zellen, Chemokin-Medium und Kontroll-Medium befüllt wird. Entweder
wird nach Ablauf eines bestimmten Zeitintervalls der Anteil migrierter Zellen in den verschiedenen
22Gelabschnitten ausgezählt oder durch Auswertung sequenzieller lichtmikroskopischer Aufnahmen
der Migrationspfad einzelner Zellen rekonstruiert [58], [128]. Neben technisch anspruchsvollen
Experimenten, wie beispielsweise der zellspezifischen Chemokinstimulation mit Beobachtung
morphologischer Veränderungen im Mikropipetten-Versuch, hat vor allen Dingen die Entwicklung
mikrofluider Assays die in vitro-Migrationsforschung voran getrieben [129]. Filigrane
Versuchsaufbauten ermöglichen die simultane Beobachtung von mehreren hundert
Einzelzellmigrationen unter örtlicher und zeitlicher Variation von Chemokin-Stimulation, in
verschiedenen 2D- und 3D-Matrizes und sogar die Integration von Zellisolation und -migration aus
wenigen Mikroliter Blut in einem Device [130]–[133]. Die hierfür benötigten Objektträger sind meist
Spezialanfertigungen aus Polydimethylsiloxan, einem auch als Dimeticon bekannten Silikonöl. Die
Herstellung ist kostenintensiv und erfordert spezielles Equipment und Handling, um stabile
Gradientenbildung und Matrixhomogenität zu gewährleisten, was ihren Einsatz limitiert [134]. Der
Informationsgehalt in Einzelzellpfaden ist hoch, birgt gleichwohl die Herausforderung der
Auswertung. Diese erfolgt nach wie vor langwierig manuell oder mit Hilfe teurer Hard- und
Software. Die Verbesserung der internen Validierung hat das automatisierte Zelltracking in den
letzten Jahren wiederum verbessert [135], [136]. Die Nachbildung interstitieller Matrix zur
Verbesserung des Verständnisses der Leukozyten-ECM-Interaktion war eine weitere Aufgabe der in
vitro-Migrationsforschung. Anwendung fanden zunächst dichte 3D-Matrizen aus Hydrogel, Kollagen
I oder ein von Sarkomzellen sezerniertes Matrixgemisch namens „Matrigel“ [31]. Vereinzelt wurden
auch am Zelldurchmesser orientierte Glaskanäle oder nanofibröse Kunststoffmembranen zur
Imitation der ECM gewählt [137]. Die tatsächliche Interaktion von PMN mit umgebendem Gewebe
und Leukozyten ist in vivo weiterhin am besten beobachtbar. Der Fortschritt der
Multiphotonenmikroskopie erlaubt inzwischen fokussierte Aufnahmen nicht nur am murinen
Cremaster-Muskel sondern auch an freipräparierten Organen lebender Mäuse in etwa 300µm Tiefe.
Durch Fluoreszenzfärbung von Zellen und Matrixbestandteilen geben diese Bilder zusätzlich
Aufschlüsse über ihre stete Interaktion [4], [138].
231.5 Fragestellung
Der folgende Abschnitt fasst den thematischen Hintergrund dieser Arbeit zusammen und umreist
die daraus abgeleiteten Fragestellungen.
Neutrophile Granulozyten bilden die größte Fraktion der Leukozyten und gehören im Falle einer
akuten inflammatorischen Reaktion zu den ersten Zellen am Ort der Entzündung. Für die
Rekrutierung von PMN aus der Blutbahn bedarf es zunächst einer Selektin-vermittelten
Entschleunigung mit nachfolgender Integrin-abhängiger Adhäsion und Transmigration über
Endothel und Basalmembran hinweg. Die weitere Migration verläuft in zellreichem Gewebe
ebenfalls adhäsiv, in fibrillärem Interstitium hingegen nicht-proteolytisch und non-adhäsiv ab, was
PMN eine schnelle Fortbewegung ermöglicht. Eine rasche Rekrutierung im Falle einer akuten
Infektion oder Trauma ist lebensnotwendig. Bei unzureichender Kontrolle bewirkt eine
überschießende Inflammation jedoch erheblichen Gewebeschaden. Die Migration von PMN bedarf
daher multipler Regulatoren, wovon eine Auswahl in dieser Arbeit untersucht werden soll. Der
Fokus liegt hierbei auf der modulierenden Wirkung von extrazellulärer Matrix, die bereits durch ihre
physikalischen Eigenschaften Einfluss auf neutrophiles Migrationsverhalten ausübt. Darüber hinaus
besteht eine stete Interaktion von Matrix und PMN. Wir gehen daher von einer Veränderung
neutrophilen Migrationsverhaltens bei Variation der Matrix aus, deren Ausprägung und
Mechanismen unzureichend geklärt sind. Des Weiteren gibt es Anhalt für immunmodulatorische
Eigenschaften des Kollagen IV-Peptids CP17, welche bisher noch nicht untersucht wurden. Aus
klinischer Sicht interessiert des Weiteren der Effekt von Glukokortikoiden, die häufig unspezifisch
antiinflammatorisch eingesetzt werden, der genaue Mechanismus dieser Wirkung jedoch
unbekannt ist. Vor dem Hintergrund der umfassenden transkriptorischen Regulation durch
Glukokortikoide nehmen wir eine Modulation der Migration neutrophiler Granulozyten an. Um den
eigentlichen Prozess der Migration auf Einzelzellebene darstellen zu können soll ein bisher für
adhäsive Zellen wie Endothel- und Fibrosarkomzellen etablierter und kommerziell verfügbarer
µSlide-Migrationsassay auf die Migration von PMN angepasst werden [139]. Ziel dieser Arbeit ist
die Darstellung und Auswertung der neutrophilen interstitiellen Migration in vitro, um folgende
Fragestellungen zu bearbeiten:
- Wie stellt sich die Migration neutrophiler Granulozyten in einfacher Matrix dar und was
verändert die Zunahme der Matrixkomplexität?
- Welchen Einfluss hat das Glukokortikoid Prednisolon auf neutrophile Migration?
- Wie verändert das Kollagen IV-Fragment CP17 die Migration neutrophiler Granulozyten?
Zum einen soll diese Arbeit durch die Bereitstellung eines kommerziell erwerbbaren Assays für non-
adhäsive Zellen zur Analyse des Migrationsprozesses unter Variation von Matrix und Stimulanz die
24Sie können auch lesen

















































