Entwicklung der quergestreiften Ösophagusmuskulatur bei Shh-Mutantmäusen - opus4.kobv.de
←
→
Transkription von Seiteninhalten
Wenn Ihr Browser die Seite nicht korrekt rendert, bitte, lesen Sie den Inhalt der Seite unten
Entwicklung der quergestreiften Ösophagusmuskulatur
bei Shh-Mutantmäusen
Aus dem Institut für Anatomie (Lehrstuhl für Anatomie und Zellbiologie)
der Friedrich-Alexander-Universität Erlangen-Nürnberg
Direktorin: Prof. Dr. S. Kürten
Dissertation
Der Medizinischen Fakultät
Der Friedrich-Alexander-Universität
Erlangen-Nürnberg
zur
Erlangung des Doktorgrades Dr. med. dent.
vorgelegt von
Julia Beckmann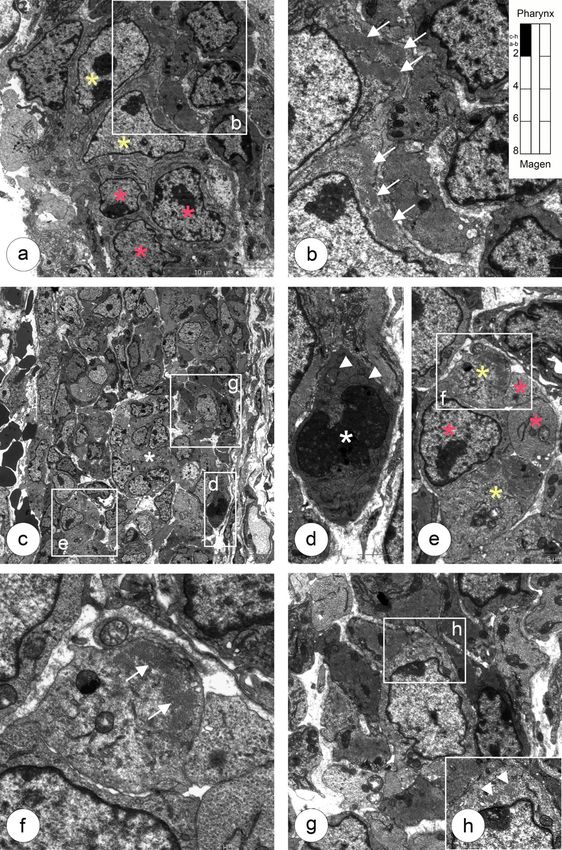
Als Dissertation genehmigt von der
Medizinischen Fakultät der Friedrich-Alexander-Universität
Erlangen-Nürnberg
Vorsitzender des Promotionsorgans: Prof. Dr. Markus F. Neurath
Gutachter: Prof. Dr. Jürgen Wörl
Gutachter: Prof. Dr. Winfried Neuhuber
Tag der mündlichen Prüfung: 16. März 2021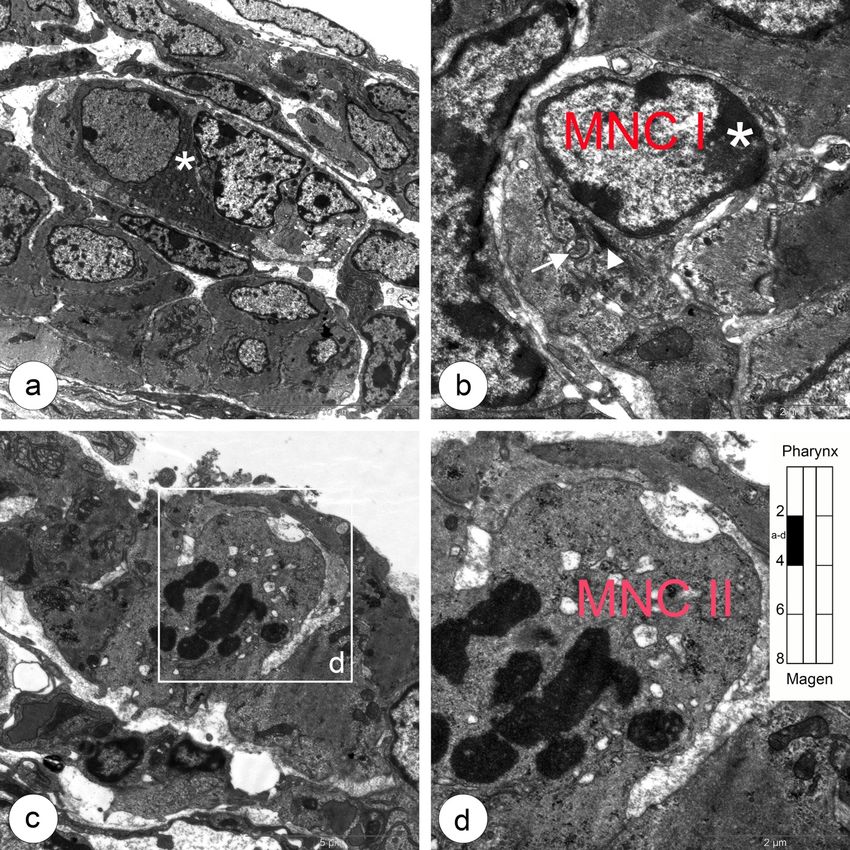
Inhaltsverzeichnis i
Inhaltsverzeichnis
1. Abstract ................................................................................................................1
1.1 Objectives ......................................................................................................1
1.2 Design & Methods..........................................................................................1
1.3 Observations & Results .................................................................................1
1.4 Conclusions ...................................................................................................2
2. Zusammenfassung ..............................................................................................3
2.1 Hintergrund und Ziele ....................................................................................3
2.2 Material und Untersuchungsmethoden ..........................................................3
2.3 Ergebnisse und Beobachtungen ....................................................................3
2.4 Praktische Schlussfolgerungen......................................................................4
3. Einleitung .............................................................................................................5
3.1 Aufbau und Funktion des Ösophagus............................................................5
3.2 Entwicklung des Ösophagus..........................................................................6
3.2.1 Embryonale Entwicklung des Ösophagus ..............................................6
3.2.2 Entwicklung der Ösophagusmuskulatur .................................................6
3.2.3 Pax7 – Funktion und Einfluss auf die Myogenese ................................10
3.3 Die enterische Co-Innervation der quergestreiften Ösophagusmuskulatur .10
3.4 Shh-Signalweg .............................................................................................11
3.5 Atresie ..........................................................................................................13
3.6 Ziele der Arbeit.............................................................................................15
4. Material und Methoden......................................................................................17
4.1 Herkunft, Zucht und Genotypisierung der Mäusemutanten .........................17
4.2 Gewebe, Präparation und Fixation ..............................................................18
4.3 Elektronenmikroskopie.................................................................................19
5. Ergebnisse .........................................................................................................20
5.1 Entwicklung der Ösophagusmuskulatur bei Shh(+/+) Kontrollmäusen ..........20
5.2 Entwicklung der Ösophagusmuskulatur bei Shh(+/-)-Mutantmäusen ............29
5.3 Entwicklung der Ösophagusmuskulatur bei Shh(-/-)-Mutantmäusen ............32
6. Diskussion..........................................................................................................40
6.1 Entwicklung der Ösophagusmuskulatur in E18 Kontrollmäusen .................40
6.2 Entwicklung der Ösophagusmuskulatur in Shh Mutantmäusen...................42
6.3 Ursprung der quergestreiften Ösophagusmuskulatur ..................................44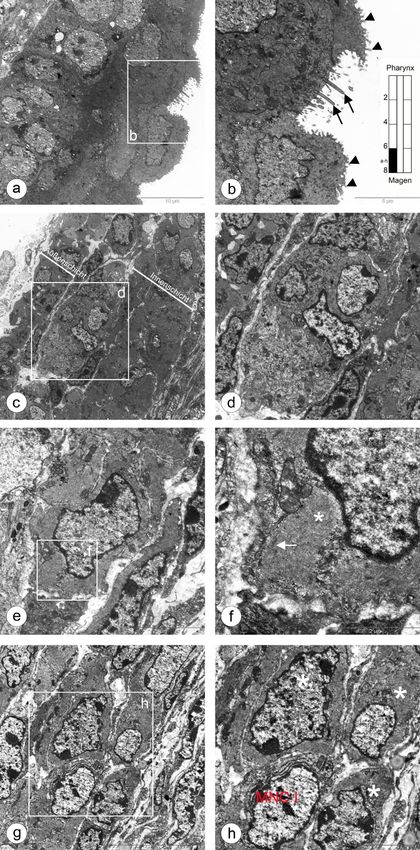
Inhaltsverzeichnis ii
6.4 Schlussfolgerung .........................................................................................44
7. Literaturverzeichnis...........................................................................................46
8. Danksagung .......................................................................................................51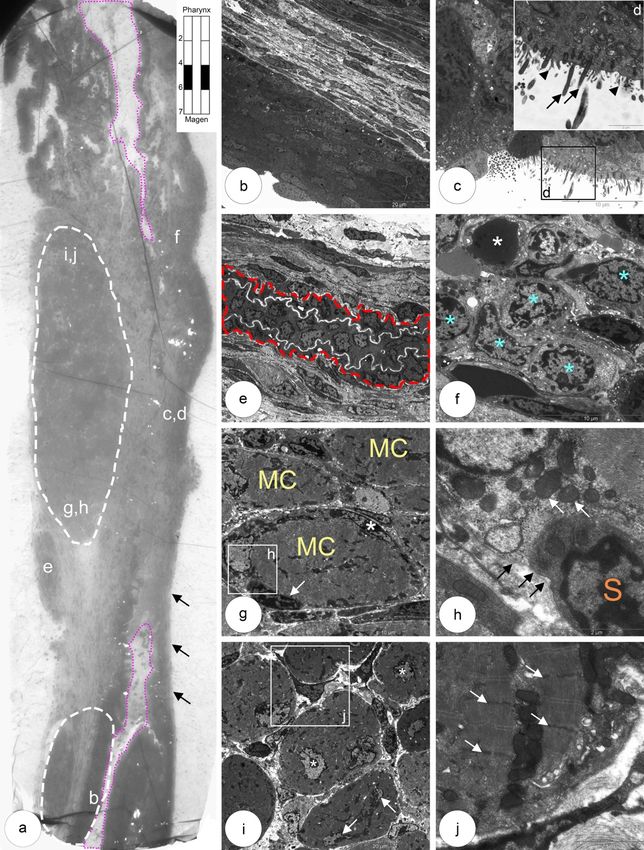
1 Abstract 1 1. Abstract 1.1 Objectives Sonic hedgehog (Shh) is a morphogen responsible for short- or long-term regulatory processes during embryonic development. Shh is expressed by the endodermal epi- thelium of the digestive and respiratory system, whereupon it is particularly active in the esophageal tissues. Here it induces different developmental processes depending on its concentration. Atresia is a condition in which there are motility and food transport disorders in the area of the esophagus. It is believed that there is a possible link be- tween a lack of Shh gene expression and malformations of the esophageal muscle tube, that cause the clinical symptoms. 1.2 Design & Methods For the present study, breeding pairs of mice were used in which the Shh gene was heterozygous changed in both sexes. Since homozygous mutant mice are not viable, embryos were analyzed on the eighteenth day of development just before birth. In total, three control mice (Shh+/+), three heterozygous (Shh+/-) and three homozygous (Shh-/-) mutant mice were included in the study, which were from four pregnant female mice. The nine esophagi of the embryos were divided into 2 mm long pieces from cra- nial to caudal, fixed and prepared for electron microscopy. 1.3 Observations & Results While Shh(+/+) mice showed a regular transformation of smooth muscle into striated muscle in a craniocaudal sequence, the analysis of Shh(-/-) mice showed that complete inhibition of gene activity leads to complex morphological changes in the mouse esoph- agus. Although mesenchymal cells differentiated into their further developmental stages, a regulated activation of proliferation and differentiation no longer took place in its conventional sequence. In addition, the amount of proliferating muscle precursor cells was significantly reduced, continuity interruptions of the esophagus occurred and the stratification of the organ had been abolished. In Shh(+/-) mice there were no or no detectable developmental changes.
1.4 Conclusions 2 1.4 Conclusions The results of the study suggest a link between defects in the Shh gene and morpho- logical aberrations in the esophagus which are already visible in embryonic develop- mental stages. The investigations indicate that Shh does not have any influence on the actual differentiation process of mesenchymal progenitor cells but has to be involved in signal cascades that allow and control a regulated developmental sequence. The available findings are of high clinical relevance, as they are the first to show morpho- logical changes on the electron microscopic level in the genetic mouse model, which contribute decisively to the further understanding of the pathogenesis of esophageal atresia.
2 Zusammenfassung 3 2. Zusammenfassung 2.1 Hintergrund und Ziele Sonic-Hedgehog (Shh) ist ein Morphogen, welches für kurz- oder langfristige Regula- tionsprozesse während der Embryonalentwicklung zuständig ist. Shh wird vom en- dodermalen Epithel der Speise- und Luftwege exprimiert, worauf es unter anderem besonders in den Geweben des Ösophagus aktiv wird. Hier induziert es in Abhängig- keit von seiner Konzentration verschiedene Entwicklungsvorgänge. Die Atresie stellt ein Krankheitsbild dar, bei welchem es zu Motilitäts- und Nahrungstransportstörungen im Bereich des Ösophagus kommt. Es wird vermutet, dass ein möglicher Zusammen- hang zwischen einer ausbleibenden Shh-Genexpression und Fehlbildungen des öso- phagealen Muskelschlauchs besteht, welche die klinischen Symptome verursachen. 2.2 Material und Untersuchungsmethoden Für die vorliegende Untersuchung wurden Zuchtpaare von Mäusen verwendet, bei de- nen das Shh-Gen in beiden Geschlechtern heterozygot verändert war. Da homozygote Mutantmäuse nicht überlebensfähig sind, wurden Embryonen am 18. Tag der Entwick- lung kurz vor der Geburt analysiert. Insgesamt wurden jeweils drei Kontrollmäuse (Shh+/+), drei heterozygote (Shh+/-) und drei homozygote (Shh-/-) Mutantmäuse in die Studie einbezogen, die von vier trächtigen weiblichen Mäusen stammten. Die neun Ösophagi der Embryonen wurden von kranial nach kaudal in 2 mm lange Stücke un- terteilt, fixiert und für die Elektronenmikroskopie aufbereitet. 2.3 Ergebnisse und Beobachtungen Während in Shh(+/+)-Mäusen eine regelmäßige Umwandlung der glatten in querge- streifte Muskulatur in einer kraniokaudalen Sequenz zu beobachten war, zeigte die Analyse von Shh(-/-)-Mäusen, dass eine vollständige Hemmung der Genaktivität zu komplexen morphologischen Veränderungen im Mäuseösophagus führt. Mesen- chymale Zellen differenzierten sich zwar in ihre weiteren Entwicklungsstufen, eine re- gulierte Aktivierung der Proliferation und Differenzierung fand jedoch nicht mehr in ih- rer herkömmlichen Abfolge statt. Hinzu kommt, dass die Menge an proliferierenden Muskelvorläuferzellen deutlich reduziert war, Kontinuitätsunterbrechungen des Öso- phagus auftraten und der typische Wandaufbau des Organs aufgehoben war. In
2.4 Praktische Schlussfolgerungen 4 Shh(+/-)-Mäusen zeigten sich keine bzw. keine nachweisbaren Entwicklungsverände- rungen. 2.4 Praktische Schlussfolgerungen Aufgrund der Untersuchungsergebnisse lässt sich auf einen Zusammenhang zwischen Defekten des Shh-Gens und morphologischen Aberationen im Bereich des Ösopha- gus schließen, die bereits in embryonalen Entwicklungsstadien sichtbar werden. Die Untersuchungen weisen darauf hin, dass Shh nicht auf die eigentliche Differenzierung der mesenchymalen Vorläuferzellen Einfluss hat, sondern an Signalkaskaden beteiligt sein muss, die eine regulierte Entwicklungsabfolge zulässt und kontrolliert. Die vorlie- genden Befunde haben hohe klinische Relevanz, da sie erstmalig morphologische Veränderungen auf elektronenmikroskopischer Ebene im genetischen Mäusemodell darstellen, die zum weiteren Verständnis der Pathogenese der Ösophagusatresie ent- scheidend beitragen.
3 Einleitung 5 3. Einleitung 3.1 Aufbau und Funktion des Ösophagus Der menschliche Ösophagus ist ein ca. 25cm langer muskulärer Schlauch, dessen Funktion größtenteils darin besteht, durch peristaltische Muskelkontraktionen den Speisebolus vom Rachen zum Magen zu befördern (Benninghoff & Drenckhahn 2008). Die Speiseröhre lässt sich von oral nach abdominal in drei Abschnitte einteilen. Von oral nach abdominal sind das die Pars cervicalis (5-8cm), die Pars thoracica (16cm) und die Pars abdominalis (1-4cm). Darüber hinaus besitzt der Ösophagus in seinem Verlauf drei physiologische Engstellen. Diese befinden sich auf Höhe des Schildknor- pels (Angustia cricoidea, 6. HWK), des Aortenbogens (Angustia aortica, 4. BWK) und beim Durchtritt durch das Zwerchfell (Angustia diaphragmatica, 10. BWK; (Benninghoff & Drenckhahn 2008; Geboes & Desmet 1978). Die Ösophaguswand ist – entspre- chend der typischen Beschaffenheit des Verdauungstraktes – aus vier Schichten auf- gebaut. Von innen nach außen formieren sich diese aus einer Schleimhaut- und Mus- kelschicht (Tunica mucosa) – welche wiederum in drei Bereiche unterteilbar ist (La- mina epithelialis, Lamina propria, Lamina muscularis mucosae) –, einer verschiebli- chen Bindegewebsschicht (Tela submucosa), einer dickeren Muskelschicht (Tunica muscularis) und einer weiteren Bindegewebsschicht (Tunica adventitia). Aufgrund der intraperitonealen Lage der Pars abdominalis wird diese in ihrer äußersten Schicht durch Bauchfell bedeckt (Tunica serosa; (Mittal 2012). Die epitheliale Auskleidung des Ösophagus entstammt dem Endoderm, wohin gegen die Lamina propria mucosae, die Lamina muscularis mucosae, die Tela submucosa und die neuralen Elemente meso- dermaler Herkunft sind (Geboes & Desmet 1978). Die Mukosa des Ösophagus ist aus mehrschichtigem unkeratinisierten Plattenepithel aufgebaut, welches ab der soge- nannten „Z-Linie“ in das Zylinderepithel der Kardia des Magens übergeht (Al-Shraim et al. 2015; Patti et al. 1997). Das Epithel dient dem Schutz des Ösophagus während teils abrasive Nahrung das Organ passiert. Submuköse Drüsen erleichtern hierbei den Nahrungstransport (Zhang et al. 2018). Die Tunica muscularis des Menschen setzt sich aus einer äußeren Längsmuskulatur- und einer inneren zirkulär angeordneten Muskelschicht zusammen. Beide haben im proximalen Abschnitt einen spiralförmigen Verlauf. In kranialen Bereichen sind die äußere und innere Muskelschicht nicht voll-
3.2 Entwicklung des Ösophagus 6 ständig voneinander getrennt, wohingegen in kaudalen Abschnitten eine klare Tren- nung vorliegt. Hierbei befindet sich in der oberen Hälfte des Ösophagus neben über- wiegend quergestreifter Muskulatur (50-60 %) auch glatte Muskulatur. In der unteren Hälfte des Ösophagus wird die Tunica muscularis vollständig von glatter Muskulatur gebildet (Kallmünzer et al. 2008). Der obere Ösophagussphinkter setzt sich aus quer- gestreifter, der untere Sphinkter aus glatter Muskulatur zusammen (Mittal 2012). Im Unterschied zum Menschen weisen andere adulte Säugetiere, wie etwa die Maus oder der Hamster, eine ausschließlich aus quergestreiften Muskelzellen bestehende Öso- phaguswandung auf, während diese bei Vögeln aus rein glatter Muskulatur besteht (Kallmünzer et al. 2008; Neuhuber & Wörl 2016; Shiina et al. 2005). 3.2 Entwicklung des Ösophagus 3.2.1 Embryonale Entwicklung des Ösophagus Der Respirationstrakt sowie der Verdauungstrakt entwickeln sich aus dem Vor- derdarm, welcher sich während der Entwicklung zwischen der Herzschlaufe und dem Neuralrohr befindet. Diese embryonale Struktur ist endodermalen Ursprungs und ent- steht am achten embryonalen Tag im Rahmen der Gastrulation. Das Vorderdarmrohr unterliegt einem Separationsprozess, welcher durch einen reziproken Signalaustausch zwischen Endoderm und Mesoderm gesteuert wird. Während sich der dorsale Anteil der Röhre zum Ösophagus differenziert, bildet der ventrale Anteil die Anlage für Trachea und Lunge. Diese dorso-ventrale Strukturierung verläuft von kaudal nach kra- nial und wird durch Signale aus der Chorda dorsalis sowie verschiedene Transkripti- onsfaktoren und Signalmoleküle (Sox2, Nkx 2.1) reguliert. Die genauen zellulären Vor- gänge, die für die Trennung von Ösophagus und Trachea sorgen, sind bisher noch ungeklärt. Unterbrechungen der zuvor genannten Regulationsprozesse führen zu un- vollständigen Separationsvorgängen und Fehlbildungen von Trachea und Ösophagus. Die beiden häufigsten Defekte stellen hierbei die Atresie und die Ausbildung einer tracheoösophagealen Fistelung dar (Billmyre et al. 2015; Jacobs et al. 2012). 3.2.2 Entwicklung der Ösophagusmuskulatur Der Ursprung der quergestreiften Muskulatur des Mäuseösophagus ist noch nicht end- gültig geklärt. In der Literatur finden sich unterschiedliche Herkunftstheorien. Zunächst wurde der Prozess der sogenannten „Transdifferenzierung“ beschrieben. Hierbei soll
3.2 Entwicklung des Ösophagus 7 es zur Umwandlung glatter in quergestreifte Muskelzellen im Mäuseösophagus kom- men (Patapoutian et al. 1995). Dieser Entstehungsansatz konnte mittlerweile jedoch durch Nachweis zweier unterschiedlicher Genotypen für glatte und quergestreifte Mus- kelzellen widerlegt werden (Rishniw et al. 2003). Ein anderes Konzept wurde im Jahr 2015 verfolgt. Laut dieser Studie sind die quergestreiften Muskelzellen des Ösophagus neben der Herz- und Kopfmuskulatur ein drittes Derivat des kardiopharyngealen Mes- oderms. Hiermit unterscheiden sie sich zur Skelettmuskulatur des Körpers, welche sich aus Stammzellen der Somiten entwickelt. Die Studie erläutert im weiteren Verlauf, dass es zu Migrationsprozessen muskulärer Vorläuferzellen aus dem Mesoderm des Kopfes zu ihrem Bestimmungsort im Mäuseösophagus kommt (Gopalakrishnan et al. 2015). Eine weitere Theorie beschreibt eine autochthone Entwicklung quergestreifter Muskelzellen des Ösophagus. Diese beruht darauf, dass Vorläuferzellen der späteren Muskulatur im Mäuseösophagus nachgewiesen werden konnten, welche sich nach entsprechender Stimulierung zur quergestreiften Muskulatur differenzierten (Wörl & Neuhuber 2005b). Die Differenzierung von quergestreiften Muskelzellen im Ösophagus setzt im Vergleich zur Extremitätenmuskulatur spät ein. Erste Vorläuferzellen der quergestreiften Musku- latur lassen sich zum Zeitpunkt der zwölften embryonalen Woche (E12.5) im proxima- len Abschnitt des Ösophagus nachweisen. Diese befinden sich zwischen den beiden glatten Muskelschichten der Tunica muscularis. Die Differenzierung der Vorläuferzel- len beginnt zunächst in der äußeren Muskelschicht und setzt sich dann in der inneren Schicht fort (Chihara et al. 2015; Gopalakrishnan et al. 2015; Kablar et al. 2000; Zhao & Dhoot 2000). Der Ersatz von glatter durch quergestreifte Muskulatur findet in der sogenannten „Übergangszone“ statt und verläuft in kraniokaudaler Richtung. Während der Myogenese durchlaufen die quergestreiften Muskelzellen verschiedene Reifesta- dien (Tabelle 3.1; Wörl & Neuhuber 2005b). Unreifere Entwicklungsstadien befinden sich kaudal, reifere weiter kranial (Wörl & Neuhuber 2005b). Im Unterschied zur Ske- lettmuskulatur lässt sich die ösophageale Myogenese nicht in zwei Entwicklungspha- sen – die primäre und sekundäre Myogenese – einteilen (Su et al. 2011; Wörl & Neuhuber 2005b). Die Ausreifung der Muskulatur des Ösophagus endet in der zweiten postnatalen Woche (P14). Je nach Mäusestamm variiert der Anteil an glatter und quer- gestreifter Muskulatur im ausgereiften Zustand. Glatte Muskulatur befindet sich bei- spielsweise bei 129Sv-Mäusen nur noch im distalen Drittel des Muskelschlauchs
3.2 Entwicklung des Ösophagus 8 (Chihara et al. 2015), wohin gegen bei C57/BI6-Mäusen quergestreifte Muskulatur bis zum unteren Ösophagussphinkter ausgereift ist (Breuer et al. 2004). Kontroverse Ansichten bestehen in der Literatur in Bezug auf die tatsächliche Genese der quergestreiften Muskulatur. In den 1990er-Jahren wurden der Nachweis einer Koexistenz zellspezifischer Marker für beide Muskelzelltypen – glatt und quergestreift – in einer individuellen Zellpopulation der Ösophagusmuskulatur als Beleg für die Um- wandlung von glatten zu quergestreiften Muskelzellen durch Transdifferenzierung ge- nommen (Patapoutian et al. 1995). In darauf folgenden Studien konnten weitere An- haltspunkte gefunden werden, die diese Meinung unterstützen (Kablar et al. 2000; Sang & Young 1997; Stratton et al. 2000). Im Gegensatz dazu halten andere Autoren die Abstammung von glatter und quergestreifter Muskulatur aus zwei unterschiedli- chen Entwicklungslinien für wahrscheinlicher, da für beide Muskelarten zum einen mit molekularbiologischen Methoden getrennte Entwicklungswege und zum anderen elektronenmikroskopisch unterschiedliche Vorläuferzellen nachgewiesen werden konnten (Rishniw et al. 2003; Wörl & Neuhuber 2005b; Zhao & Dhoot 2000). Ebenso umstritten wird die Frage diskutiert, ob eine Apoptose glatter Muskelzellen stattfindet oder nicht. Während Wörl und Neuhuber zahlreiche apoptotische glatte Muskelzellen in der Übergangszone von glatter zu quergestreifter Muskulatur dokumentieren konn- ten (Wörl & Neuhuber 2005b), sprechen andere Autoren diesem Vorgang eine unter- geordnete Rolle zu, da in ihren Untersuchungen der Großteil an glatten Muskelzellen nach kaudal wandert, um sich hier an der Bildung des unteren Ösophagussphinkters zu beteiligen (Rishniw et al. 2007).
3.2 Entwicklung des Ösophagus 9
Zelltyp Abkürzung Morphologische Kriterien
1. Glatte Muskelzellen
Helle glatte Muskelzellen L-SMC Zentral liegender heterochromatischer Nucleus,
Zellorganelle an beiden Polen des Nucleus an-
gesiedelt, Hauptteil des hellen Zytoplasmas mit
dünnen Filamenten gefüllt, dense bands
(Stratton et al. 2000)
Dunkle glatte Muskelzellen D-SMC Gleiche Eigenschaften wie L-SMC, aber dunk-
les Zytoplasma und Nucleus
Apoptotische glatte Muskel- A-SMC Gleiche Eigenschaften wie D-SMC, aber dunk-
zellen les kondensiertes Chromatin und Zytoplasma
2. sich entwickelnde quergestreifte Muskelzellen
a. Abwesenheit von Myo-
fibrillen
Mononukleäre Zellen Typ I MNC-I Undifferenzierte runde oder spindelförmige
(Muskelvorläuferzellen) mesenchymale Zellen mit wenig Zytoplasma
und wenigen Zellorganellen
Mononukleäre Zellen Typ II MNC-II Undifferenzierte spindelförmige Zellen mit grö-
(Myoblasten) ßerem Anteil an Zytoplasma mit Zellorganellen,
v.a. rER, freie Ribosomen und Mitochondrien
(Zhao & Dhoot 2000)
Mononukleäre Zellen Typ MNC-III Undifferenzierte Zellen mit den gleichen Eigen-
III schaften wie MNC-II, zusätzlich deutlich hell-ge-
(Myoblasten) färbtes Zytoplasma an einem Pol des Nucleus
Satellitenzellen SC Undifferenzierte mononukleäre Zellen, einge-
schlossen in die Basallamina einer Muskelfaser,
aber abgetrennt durch eine Plasmamembran
(Zhao & Dhoot 2000)
b. Vorkommen von Myo-
fibrillen
Myozyten MC Differenzierte mononukleäre Zellen mit zentral-
ständigem Kern und – abhängig vom Enwick-
lungsstatus – Bündel von Myofibrillen oder ein-
zelnen Sarkomeren
Myotuben MT Differenzierte, verlängerte, aus mononukleären
Zellen fusionierte multinukleäre Zellen, zentral-
ständige Kerne, mehrreihige Sarkomere
Myofasern MF Differenzierte multinukleäre Zellen mit randstän-
digen Kernen, mehrreihige Sarkomere
Begriffe und Definitionen modifiziert nach Wörl und Neuhuber (2005)
Tabelle 3.1: Zusammengefasste morphologische Kriterien verschiedener Typen glatter und sich entwickelnder querge-
streifter Muskelzellen in der Tunica muscularis eines P4 Ösophagus (Wörl & Neuhuber 2005b).3.3 Die enterische Co-Innervation der quergestreiften Ösophagusmuskulatur 10
3.2.3 Pax7 – Funktion und Einfluss auf die Myogenese
Pax7 ist ein Protein, welches zu der Gruppe der Paired-Box-Gene (Pax-Gene) gehört
und unter anderem für die strukturelle postnatale Entwicklung der Tunica muscularis
des Ösophagus verantwortlich ist (Chihara et al. 2015; Jostes et al. 1990; Wörl et al.
2009). Hier kodiert Pax7 für gewebsspezifische Transkriptionsfaktoren, welche die
Zellproliferation in der Übergangszone und damit die Entwicklung der quergestreiften
Ösophagusmuskulatur von proximal nach distal unterstützen. Ein Verlust von Pax7
beeinflusst die Myogenese im Bereich des Ösophagus in mehrfacher Hinsicht: Neben
einer verzögert einsetzenden Entwicklung quergestreifter Muskelzellen zeigt sich auch
eine geringere Mitoserate in der Übergangszone. Alle zellulären Entwicklungsstufen
(siehe Tabelle 3.1; Wörl & Neuhuber 2005b) sind vorhanden, liegen jedoch in redu-
zierter Anzahl vor. Neben den organspezifischen Störungen des Ösophagus zeigen
sich auch allgemeine Wachstumsdefizite an homozygot veränderten Mutantmäusen
(Pax7-/-), welche mit einer verminderten Lebensdauer von maximal drei bis vier Wo-
chen einhergehen (Chihara et al. 2015; Wörl et al. 2009). Eine weitere Aufgabe von
Pax7 besteht darin, die Entwicklung, Aktivierung und Regulation von Satellitenzellen
zu koordinieren. Satellitenzellen sind mononukleäre Zellen, welche sich zwischen der
Basallamina und der Plasmamembran ausgereifter Muskelfasern befinden. Sie spielen
eine entscheidende Rolle für das Muskelwachstum und deren Regeneration. Mit zu-
nehmendem Entwicklungsstadium des ösophagealen Muskelschlauchs nimmt die An-
zahl der Satellitenzellen bei Mäusen ab. Ein Fehlen von Pax7 äußert sich hier durch
ein späteres Auftreten von Satellitenzellen, gefolgt von einer signifikant stärkeren Ab-
nahme selbiger im reiferen Entwicklungsstatus. Die Veränderungen bei den Pax7-ab-
hängigen Muskelvorläuferzellen führen zu einem verzögertem Wachstum der querge-
streiften Muskulatur mit Ausbildung eines gemischt-muskulären Ösophagus (Wörl et
al. 2009).
3.3 Die enterische Co-Innervation der quergestreiften Ösophagusmusku-
latur
Das klassische Konzept der Innervation quergestreifter Muskelzellen des Ösophagus
nimmt an, dass cholinerge Motorneurone des N. vagus an den motorischen Endplatten
inserieren. Diese finden ihren Ursprung im Nucleus ambiguus des Hirnstammes, wel-
cher zugleich als Kontrollzentrum für die Peristaltik dient. Die Aufgabe des zwischen3.4 Shh-Signalweg 11 der äußeren und inneren Muskelschicht gelegenen Plexus myentericus bleibt in dieser Ansicht unklar (Kallmünzer et al. 2008; Shiina et al. 2010; Wörl & Neuhuber 2005a). Im Jahr 1994 ließ sich erstmals an Ratten neben der vagalen auch eine enterische Innervation der motorischen Endplatten belegen. Dieser duale Innervationsweg konnte weder in der Skelettmuskulatur, noch in einem anderen Abschnitt des Verdauungs- trakts nachgewiesen werden und wird seither mit dem Begriff der „enterischen Co- Innervation“ beschrieben. Zwei morphologisch und chemisch unterschiedliche Arten von Neuronen konnten differenziert werden: Während dicke Axone aus dem N. ambi- guus stammen, inserieren zusätzlich dünne variköse Axone an den motorischen End- platten, deren Ursprung mit hoher Wahrscheinlichkeit im Plexus myentericus des au- tonomen Nervensystems liegt (Neuhuber & Wörl 2016; Neuhuber et al. 1994; Wörl & Neuhuber 2005a). Die Funktion der enterischen Co-Innervation scheint darin zu liegen, durch einen loka- len Reflexbogen des autonomen Nervensystems, Efferenzen des N. vagus zu inhibie- ren und somit Einfluss auf die Regulation der Ösophagusmotilität zu haben (Boudaka et al. 2007; Izumi et al. 2003; Neuhuber & Wörl 2016; Shiina et al. 2010). Das Prinzip der ösophagealen Co-Innervation konnte neben der Ratte auch am Menschen sowie in anderen Säugetiere und Nicht-Säugetiere nachgewiesen werden (Hempfling et al. 2009; Kallmünzer et al. 2008; Neuhuber & Wörl 2016; Wörl & Neuhuber 2005a). 3.4 Shh-Signalweg Sonic-Hedgehog (Shh) stellt neben Indian-Hedgehog (Ihh) und Desert-Hedgehog (Dhh) einen der drei Hedgehog-Signalwege bei Wirbeltieren dar, welche von Nüsslein und Vollhard im Jahr 1980 erstmals beschrieben wurden (Echelard et al. 1993; Nüsslein-Volhard & Wieschaus 1980). Sonic-Hedgehog wird als sogenanntes Morpho- gen bezeichnet, da es in der Embryonalentwicklung die weitere Morphogenese diver- ser Zellen steuert. Die Genexpression wird zunächst aus dem Neuralrohr unter Ein- fluss durch die Chorda dorsalis reguliert (Choudhry et al. 2014; Ingham & McMahon 2001). Hierbei hat vor allem die Konzentration des Gens im vorliegenden Gewebe ent- scheidende Auswirkung auf die weitere Morphogenese. In geringer Konzentration wird z.B. die Entwicklung vertebraler Neuronen, in hoher die von Motoneuronen und in sehr hoher Konzentration die der Bodenplatte induziert (Choudhry et al. 2014; Ericson et al. 1997; Jessell 2000; Roelink et al. 1995). Damit kommt dem Shh-Gen eine wichtige
3.4 Shh-Signalweg 12 Rolle bei der kurz- und langfristigen Regulierung unterschiedlicher Entwicklungspro- zesse, wie z.B. dem Wachstum oder der axialen Strukturierung von Geweben sowie der Zelldifferenzierung zu (Choudhry et al. 2014; Ingham & McMahon 2001; Simpson et al. 2009). Der Signalweg des Shh-Gens stellt einen komplexen Prozess dar, welcher auf mehreren Stufen reguliert wird. Der Transport vom Sekretions- zum Wirkort wird zuletzt durch das Enzym „Tout-Velu“ gesteuert. Daraufhin bindet Shh an seinen Memb- ranrezeptoren Ptc („Patched“) und Smo („Smoothened“), um eine Signalkaskade im Zellinneren auszulösen und so den Zellkern zur Genexpression anzuregen (Choudhry et al. 2014; Mann & Beachy 2004). Sonic-Hedgehog wird v.a. im endodermalen Epithel von Mund, Trachea, Lunge und Ösophagus exprimiert, während Indian-Hedgehog in kaudal gelegeneren Abschnitten des Verdauungssystems seine Wirkung erzielt (Mao et al. 2010; Tsukiji et al. 2014). Shh ist an der anteroposterioren Anordnung der Gliedmaßenknospen sowie dem dor- soventralen Aufbau neuraler Strukturen beteiligt (Ioannides et al. 2003; Riddle et al. 1993; Roelink et al. 1995). Durch Studien an Mäusen konnte nachgewiesen werden, dass Shh im endodermalen Epithel entscheidenden Einfluss auf die Zellproliferation im Mesenchym und dessen Differenzierung zu glatter Muskulatur hat (Tsukiji et al. 2014). Im Gegensatz dazu beobachteten Ioannides et al., dass Shh aufgrund seiner klaren Expressionsgrenzen keinen Einfluss auf die eigentliche Differenzierung von re- spiratorischem und gastrointestinalem Gewebe an sich hat, sondern auf die physiolo- gische Trennung dieser Gewebe (Ioannides et al. 2003). Außerdem scheint Shh an der Morphogenese unterschiedlicher Hautanhangsgebilde, wie z.B. Haare, Zähne o- der auch den respiratorischem System beteiligt zu sein (Chuong et al. 2000). Liegt eine Störung im Bereich der Genexpression von Shh vor, hat dies Einfluss auf die embryonale Weiterentwicklung unterschiedlicher Strukturen und Gewebe. Liting- tung et al. (Litingtung et al. 1998) konnten in ihrer Studie von 1998 nachweisen, dass ein Fehlen von Shh die Proliferation mesenchymaler Zellen negativ beeinflusst. Bei Untersuchungen an Shh(-/-) Mäusen wurde ein erhöhter Zelltod bei gleichzeitig vermin- derter Zellproliferation festgestellt. Darüber hinaus fand im Vergleich zu einer Kontroll- gruppe keine Trennung von Ösophagus und Trachea statt und die Entwicklung der Lungenknospen unterlag einer Verzögerung. Vergleichbare Ergebnisse konnten auch
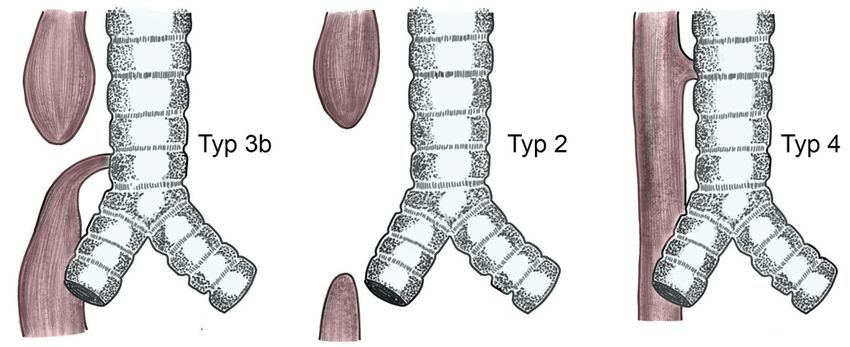
3.5 Atresie 13 an mit Adriamycin behandelten Mäuseembryos bestätigt werden und legen den Ver- dacht nahe, dass eine fehlerhafte Shh-Expression zur Entstehung von Atresie und tracheoösophagealer Fistelung beiträgt (Ioannides et al. 2003). Weitere entscheidende Defekte bei Verlust der Shh-Funktion konnten im Bereich des Vorderhirns und dessen Derivaten (Augenbläschen, Telenzephalon, Nasenhöhle etc.) diagnostiziert werden (Chiang et al. 1996). Korrelationen bestehen zudem zwischen einer Überexpression oder Reexprimierung von Shh und der Entstehung des Barrett-Ösophagus bei Re- fluxkrankheiten (Wang et al. 2014). Zuletzt konnte ein Zusammenhang zwischen Ent- wicklungsstörungen der Ösophagusmuskulatur sowie der Entstehung eines Megaöso- phagus bei Verlust des Shh-Gens nachgewiesen werden (Jia et al. 2018). 3.5 Atresie Mit dem Begriff „ösophageale Atresie“ wird eine Gruppe von Anomalien im Bereich der Speiseröhre zusammengefasst, bei denen es zu einer Kontinuitätsunterbrechung des Ösophagus kommt. Hierbei können unterschiedliche Erscheinungsformen auftreten, welche 1929 durch die Klassifikation nach Vogt in fünf Typen zusammengefasst wur- den (Typ 1, Typ 2, Typ 3a, 3b und 3c). Diese Einteilung nach Vogt wurde später mo- difiziert und erweitert (Spitz 2007). Die drei häufigsten Anomalien sind eine Atresie mit Ausbildung einer distalen tracheoösophagealen Fistel (Typ 3b nach Vogt; 86 %), eine Atresie ohne Entstehung einer tracheoösophagealen Fistel (Typ 2 nach Vogt; 7 %) und die Ausbildung einer tracheoösophagealen Fistel ohne eine Ösophagusatresie ( in der Originalarbeit von Vogt 1929 noch nicht beschrieben, später teilweise als Typ 4 nach Vogt bezeichnet; 4 %; siehe Abbildung 3.1; (Spitz 2007; Vogt 1929). Die Inzidenz der Erkrankung liegt zwischen 1:2500 und 1:3500 aller Lebendgeburten (Dutta et al. 2000; Spitz 2007). Abbildung 3.1: Häufigste Arten der Atresie (modifiziert nach (Spitz 2007)).
3.5 Atresie 14 Bei den betroffenen Patienten kommt es bereits vor operativen Eingriffen zu Motilitäts- störungen des Ösophagus, welche sich in den ersten Lebenswochen in Form von gast- roösophagealem Reflux, fehlgeleitetem Nahrungstransport und respiratorischen Prob- lemen zeigen können (Aspirot & Faure 2013; Faure & Righini Grunder 2017; Qi et al. 1999). Auch nach erfolgter Behandlung der Atresie können die Motilitätsstörungen per- sistieren oder verstärkt auftreten (Aspirot & Faure 2013; Faure & Righini Grunder 2017). Die häufigsten Komplikationen stellen hierbei der Reflux und die Dysphagie dar, welche im weiteren Verlauf zu Entzündungen des Ösophagus, einer eingeschränkten Verdauung und Metaplasien der Mucosa führen können. Zwischen 21-84 % der be- handelten Atresie-Patienten weisen Dysphagien auf, wobei die Symptome in Abhän- gigkeit des Patientenalters einer hohen Variabilität unterliegen. Fast jeder an Atresie erkrankte Mensch weist eine abnormale Peristaltik des Ösophagus auf. Hierbei lassen sich mit Hilfe der Manometrie drei Typen unterscheiden: Typ 1 zeigt keine peristalti- sche Aktivität, bei Typ 2 lassen sich isolierte distale Kontraktionen nachweisen und Typ 3 zeichnet sich durch einen Druckaufbau im Ösophagus aus (Biller et al. 1987; Faure & Righini Grunder 2017; Lemoine et al. 2013). Die Ätiologie der ösophagealen Atresie ist weitgehend unbekannt, wobei drei mögliche Hypothesen in Betracht gezogen werden. Zum einen kommen neuronale und musku- läre Defekte während der embryonalen Entwicklung in Frage, eine zweite Ursache stellen operative Eingriffe dar und zuletzt wird die Ösophagitis als Auslöser vermutet (Aspirot & Faure 2013; Boleken et al. 2007; Dutta et al. 2000; Faure & Righini Grunder 2017; Li et al. 2007). Darüber hinaus wird eine Assoziation mit einer fehlerhaften Genexpression des Shh-Gens vermutet (Litingtung et al. 1998; Spitz 2007). Um den Auslöser der Anomalien zu identifizieren, wurden bereits mehrere Studien durchgeführt. Dennoch konnte keine eindeutige Ursache ermittelt werden. Beispiels- weise wurden histologische Untersuchungen am atretischen Ösophagus des Men- schen vor und nach operativen Eingriffen vorgenommen. Hierbei konnte unter ande- rem festgestellt werden, dass es zu einer fehlerhaft angeordneten Muskulatur sowie Fibrosen des Ösophagus gekommen war. Außerdem zeigte sich eine Verzögerung der neuronalen Differenzierung sowie der Organisation des Plexus myentericus (Zuccarello et al. 2009). Bereits 1986 konnte an unbehandelten Patienten eine vermin- derte Ausprägung des Plexus nachgewiesen werden, welche vor allem im distalen Ösophagus durch eine stark reduzierte Menge an neuralem Gewebe zu sehen war
3.6 Ziele der Arbeit 15 (Nakazato et al. 1986). Des Weiteren ließen sich in postoperativ Biopsien verstärkt bindegewebige Ablagerungen zwischen glatten Muskelfasern des Ösophagus nach- weisen. Ein zunehmender Verlust der Zell-zu-Zell-Kontakte glatter Muskelzellen sowie die Ausbildung von Vakuolen, Fetteinlagerungen und knollenförmigen Ausstülpungen im Zytoplasma konnte dokumentiert werden (Al-Shraim et al. 2015). Weitere Studien fanden an fetalen Ratten statt, bei welchen durch Injektion von Adriamycin eine öso- phageale Atresie bzw. tracheoösophageale Fistelungen ausgelöst wurden. In Folge dessen konnten diffuse Gruppierungen von Nervengewebe sowie eine verminderte Nervendichte und ausbleibende Plexusbildung festgestellt werden (Cheng et al. 1999; Qi et al. 1999). Die Ergebnisse der Tierversuche lassen sich sehr gut mit den entspre- chenden Krankheitsbildern des Menschen vergleichen. Sie zeigen, dass tracheoöso- phageale Anomalien als Folge einer retardierten embryonalen Entwicklung resultieren, da alle Entwicklungsstadien der verschiedenen Gewebe vorhanden sind, jedoch erst zu einem verzögerten Zeitpunkt in Erscheinung treten (Merei et al. 1997). Als therapeutische Maßnahmen der Atresie wird ein chirurgisches Vorgehen gewählt (Spitz 2007). Derzeit werden postoperativ primär die Folgen der ösophagealen Motili- tätsstörungen, wie etwa die Ösophagitis therapiert (Faure & Righini Grunder 2017). Neue Untersuchungsmethoden, wie die „Pressure-Flow-Analysis“ sollen Aufschluss über die Ösophagusmotilität geben (Faure & Righini Grunder 2017; Rayyan et al. 2015) und damit bessere Behandlungsmethoden ermöglichen. 3.6 Ziele der Arbeit In dieser Arbeit wird mit Hilfe von Elektronenmikroskopie an Ösophagi von embryona- len Mäusen des Stadiums E18 die Entwicklung der quergestreiften Muskulatur der Tu- nica muscularis untersucht. Bei den Mäusen wurde zuvor das Shh-Gen gezielt verän- dert, so dass in die Untersuchung neben Kontrollmäusen heterozygote (Shh+/-) und homozygote (Shh-/-) Mutantmäuse einbezogen werden. Das Stadium E18 wurde aus- gewählt, da homozygote (Shh-/-) Mutantmäuse nicht überlebensfähig sind. Die Ösophagi der Kontroll- und Mutantmäuse werden den Embryonen entnommen und für die Elektronenmikroskopie systematisch aufgearbeitet. Die Untersuchung der Öso- phagi erfolgt schrittweise von kaudal nach kranial an 2 mm Längsschnitten. Im Stadium
3.6 Ziele der Arbeit 16 E18 ist neben glatter Muskulatur im kaudalen Bereich bereits quergestreifte Muskula- tur im kranialen Bereich ausgebildet, so dass alle Entwicklungsstufen der Ösophagus- myogenese in diesem Stadium beurteilt werden können. Ziel der vorliegenden Arbeit ist es, durch Vergleich von Kontroll- und Shh Mutantmäu- sen, einen möglichen Einfluss von Shh auf die schrittweise Umwandlung von glatter zu quergestreifter Muskulatur in der Tunica muscularis des Ösophagus herauszufin- den. Dazu gibt es bisher keine Untersuchungen. Die Ergebnisse dieser Untersuchung könnten die Hypothese erstmalig auf morphologischer Ebene untermauern, dass De- fekte im Shh-Gen an der Ausbildung von Ösophagusatresien bei menschlichen Föten mitbeteiligt sind.
4 Material und Methoden 17 4. Material und Methoden 4.1 Herkunft, Zucht und Genotypisierung der Mäusemutanten Heterozygote Zuchtpaare von Mäusen mit einer gezielten Zerstörung des Sonic hedgehog (Shh) Gens wurden von Dr. Chin Chiang (Department of Cell Biology, Van- derbilt University Medical Center, Nashvill, Tennessee, USA) zur Verfügung gestellt (Chiang et al. 1996). Der genetische Hintergrund des Mäusestammes war C57/Bl6. Die Weiterzucht im Präklinischen Experimentellen Tierzentrum (PETZ) der Universität Erlangen-Nürnberg erfolgte durch heterozyote Männchen der Shh-Mutantlinie mit weiblichen C57/BL6 Wildtyp-Mäusen. Für die vorliegende Untersuchung wurden Mäuse des embryonalen Entwicklungsta- ges 18 (E18) verwendet, die folgende Genotypisierung aufwiesen: Wildtyp-Kontrollen (Shh+/+), heterozygote (Shh+/-) und homozygote (Shh-/-) Shh-Mutanten. Das Stadium E18 wurde verwendet, da homozygote (Shh-/-) Mäuse nicht überlebensfähig sind. Pro Genotyp wurden jeweils drei Embryonen verwendet, die von mindestens zwei trächti- gen Weibchen stammten. Zur Gewinnung der Kontroll- und Mutantmäuse wurden zeu- gungsfähige Shh(+/-) Männchen mit zeugungsfähigen Shh(+/-) Weibchen am Abend zu- sammengesetzt und am nachfolgenden Morgen auf die Existenz eines vaginalen Pfropfens untersucht. Beim Vorhandensein eines vaginalen Pfropfens wurde der Tag E0 festgelegt und die Embryonen am 18. Entwicklungstag entnommen. Genomische DNA wurde über eine Schwanzbiopsie und Inkubation des Gewebes bei 55 °C in einem Puffer gewonnen, der folgende Substanzen enthielt: 5 mM Ethylendia- mintetraessigsäure (EDTA), 0,1 M TRIS Hydrochlorid (pH 7,5), 0,2 % Natriumdode- cylsulfat (SDS), 0,2 M Natriumchlorid und 0,2 mg/ml Proteinase K (Carl Roth, Karls- ruhe, Deutschland). Nach der Proteinase K Verdauung des Gewebes erfolgte eine Phenol/Chloroform/Isoamylalkohol (25:24:1)-Extraktion mit anschließender Isopropa- nol Fällung der DNA. Der Shh-Genotyp wurde über die Polymerase Kettenreaktion (PCR; 94 °C, 1 min; 60 °C, 1 min; 72 °C, 2 min für 35 Zyklen) unter Verwendung von vier Primern bestimmt: P1, 5´-GACCATGTCTGACACTTAGGTTCC-3´; P2, 5´-GAAGGCCAGGAGGAGAAGGCTCAC-3´; P3, 5´-CTGTGCTCGACGTTGTCACTG-3´, P4, 5´-GATCCCCTCAGAAGAACTCGT-3´.
4.2 Gewebe, Präparation und Fixation 18 Die amplifizierten PCR-Produkte wurden mit einem 1,5 %igen Agarose Gel analysiert und mit Ethidiumbromid angefärbt. Ein 329 Basenpaare (bp) Produkt entsprechend dem Shh Gen wurde in Shh(+/+) und ein ca. 600 bp Produkt in Shh(-/-) Mäusen nachge- wiesen; beide Produkte waren in Shh(+/-) Mäusen vorhanden (Abbildung 4.1). Abbildung 4.1: Analyse der Polymerasekettenreaktion (PCR) von Shh-Kontrollmäusen (Shh+/+) im Vergleich zu Mutantmäu- sen (Shh+/-, Shh-/-). In Wildtyp-Mäusen (Shh+/+) ist nur ein 329 bp Produkt zu sehen, in homozygoten (Shh-/-) nur ein 600 bp Produkt und in heterozygoten (Shh+/-) Mutantmäusen sind beide Banden zu sehen. Auf der linken Seite ist eine 50 bp DNA-Leiter mit einer helleren Bande bei 350 und 800 bp mitgelaufen. 4.2 Gewebe, Präparation und Fixation Trächtige Weibchen, einschließlich der Embryonen, wurden mit einer intraperitonealen Überdosis Thiopental (250 mg/kg Körpergewicht) getötet und anschließend die Emb- ryonen entnommen. Zuerst wurden an den Embryonen Schwanzbiopsien für die Ge- notypisierung gewonnen, danach Thorax und Abdomen geöffnet und unter einem Prä- parationsmikroskop die Ösophagi mit Kehlkopf und Magen entnommen. Der Kehlkopf und Magen wurde vom Ösophagus abgetrennt, die Ösophaguslänge gemessen und der Ösophagus in einer kraniokaudalen Sequenz in 2 mm lange Stücke geteilt und für 12 Stunden in 2,5 %iger Phosphat-gepufferter Glutaraldehyd-Lösung (Carl Roth, Karlsruhe, Deutschland; pH 7,4) bei 4 °C fixiert. Nach der Fixierung erfolgte eine Spü- lung in Phosphatpuffer über Nacht bei 4 °C und eine weitere Fixierung für vier Stunden bei 4 °C in 1 %iger Osmiumtetroxid-Lösung, die mit 1,5 %iger Kaliumhexacyanoferrat-
4.3 Elektronenmikroskopie 19 Lösung versetzt war (Karnovsky 1971). Nach einer weiteren Spülung in Phosphatpuf- fer für zwei Stunden wurden die Gewebestücke in einer aufsteigenden Ethanol-Serie dehydriert (Carl Roth; 70 %, 60 min; 80 %, 30 min; 90 %, 30 min; 100 %, 2x30 min) und mit folgenden Lösungen in reine EPON 812-Ersatzlösung überführt: (1) Etha- nol/Aceton-Mischung (1:1; 30 min), (2) reines Aceton (Carl Roth; 30 min), (3 und 4) Aceton/EPON 812-Ersatzlösung-Mischung (2:1; 30 min; 1:2; 30 min) und (5 bis 7) reine EPON 812-Ersatzlösung (2x 60 min und über Nacht). Als Epon 812 Ersatzlösung wurde Glycidether 100 (Carl Roth) verwendet. Nach Zusatz des Beschleunigers DMP- 30 (Carl Roth) zur letzten Epon 812-Ersatzlösung wurden die Gewebeproben in mit Harzlösung gefüllten BEEM®-Kapseln gelegt und bei 60 °C und 80 °C für ca. 24 Stun- den polymerisiert. Einzelne Ultradünnschnitte von Epon-eingebetteten Gewebe wur- den auf Pioloform-beschichteten Kupfernetzchen (ovales Loch; Plano, Wetzlar, Deutschland) aufgespannt und mit Bleizitrat (2,8 %, in 0,2 M NaOH-Lösung) und Uranylacetat (10 %, in 40 %iger wässriger Ethanollösung) angefärbt und luftgetrock- net. Die Anfärbung erfolgte auf einer mit Parafin überzogenen Glasplatte, auf der die beiden Färbelösungen nacheinander aufgetropft wurden. Pro Tropfen wurde ein Kup- fernetz aufgelegt. Die jeweiligen Spülungen nach den Färbelösungen erfolgten mit destilliertem Wasser. 4.3 Elektronenmikroskopie Die kontrastierten Ultradünnschnitte wurden mit einem Transmissions-Elektronenmik- roskop Zeiss 906 (Carl Zeiss NTS GmbH, Oberkochen, Deutschland) ausgewertet, das mit einer digitalen Kamera (TRS 2048, Albert Tröndle Dünzelbach, Moorenweis) ausgestattet war. Die Bildbearbeitung bezüglich Helligkeit und Kontrast sowie die Ge- staltung der Bildtafeln erfolgte mit dem Adobe Photoshop CS6 (Adobe Systems, San José, CA, USA).
5 Ergebnisse 20
5. Ergebnisse
In die Untersuchung wurden Längsschnitte von insgesamt neun Ösophagi einbezo-
gen, von denen drei homozygot (Shh-/-) und drei heterozygot (Shh+/-) veränderte Mäu-
semutanten waren, sowie drei als Kontrollgruppe (Shh+/+) dienten. Die Mäuse befan-
den sich zum Zeitpunkt der Analyse im Entwicklungsstadium E18. Unter dem Elektro-
nenmikroskop (EM) konnte so auf der gesamten Länge der Ösophagi eine verglei-
chende Beurteilung der sich entwickelnden Muskulatur und deren umliegenden Ge-
webe stattfinden.
5.1 Entwicklung der Ösophagusmuskulatur bei Shh(+/+) Kontrollmäusen
Maus 1
Maus 2
Maus 3
0 2 4 6 8 10 12 14
Länge E18 Ösophagus in mm
quergestreifte Muskulatur Übergangszone glatte Muskulatur
Abbildung 5.1: Aufbau der Tunica muscularis der E18-Kontrollmäuse (Shh+/+)
Die absolute Länge der Ösophagi in der Kontrollgruppe variierte zwischen 8 und
12 mm. Das durchschnittliche Gewicht der Mäuse betrug 1,0±0,4 g. Die Tunica mus-
cularis ließ sich in zwei bzw. drei Abschnitte, entsprechend ihres Entwicklungsstandes
einteilen: einen kranial gelegenen, hauptsächlich aus quergestreifter Muskulatur be-
stehenden Anteil, eine Übergangszone und einen kaudalen Bereich, welcher aus-
schließlich glatte Muskelzellen beinhaltete (Abbildung 5.1). Da die Entwicklung in den
Ösophagi der Kontrollmäuse weitestgehend identisch war, wurden exemplarisch die
EM-Bilder von Maus 1 verwendet und in kaudokranialer Richtung beschrieben. Ledig-
lich die Bilder aus Abbildung 5.8 sind Aufnahmen des Ösophagus von Maus 3, da hier5.1 Entwicklung der Ösophagusmuskulatur bei Shh(+/+) Kontrollmäusen 21 kranial bereits eine ausgeprägtere Entwicklung der quergestreiften Muskulatur stattge- funden hatte. Die hinzugefügte Skala pro Abbildung zeigt, in welchem Abschnitt des Ösophagus die Bilder aufgenommen wurden. Das Lumen des Ösophagus war in seiner gesamten Länge überwiegend von einem mehrschichtigen kubischen Epithel ausgekleidet, dessen Oberfläche durchgehend mit Mikrovilli und teilweise mit Kinozilien bedeckt war (Abbildung 5.2a,b). Der abdominale Ösophagusabschnitt wurde ausschließlich von glatter Muskulatur aus- gekleidet. Diese zeigte im proximalen mehr Zytoplasma als im distalen Bereich. Der Anteil an spindelförmigen, parallel angeordneten glatten Muskelzellen nahm in kaudo- kranialer Richtung ab. Neben vielen hellen glatten Muskelzellen (L-SMC) befanden sich wenige dunklere glatte Muskelzellen (D-SMC) in der Tunica muscularis. Während sich die Zellorganellen einiger glatten Zellen bereits an einem Pol des Zellkerns kon- zentrierten, lagen diese bei einer Vielzahl von glatten Muskelzellen noch in der Peri- pherie verteilt (Abbildung 5.2e; Abbildung 5.3f). Myofilamente sowie dense bands wa- ren eindeutig sichtbar (Abbildung 5.2f). Die Differenzierung in eine äußere schmalere und eine innere dickere Muskelschicht verlor sich nach distal. Die Orientierung der Muskelschichten verlief vorwiegend in einer inneren längsorientierten und einer äuße- ren zirkulär angeordneten Muskelschicht, wobei diese Anordnung nicht fortlaufend er- kennbar war. Mesenchymale Zellen kamen vermehrt v.a. im kranialen Bereich des ab- dominalen Segments zwischen den glatten Muskelzellen der inneren Schicht vor (Abbildung 5.2g,h). Weitere mesenchymale Gruppen lagen zwischen beiden Muskel- schichten. In diesem Bereich ließ sich vereinzelt auch bereits differenziertes Nerven- gewebe erkennen (Abbildung 5.2c,d). Im Randbereich des Muskelschlauchs lagen viele mit Erythrozyten gefüllte Kapillaren, welche durch ihr dunkles Erscheinungsbild apoptotischen Zellen ähnelten (Abbildung 5.3d). In der sich nach kranial anschließenden Übergangszone war ebenfalls eine Aufteilung in eine äußere und eine innere Muskulaturschicht erkennbar. Die ab hier einsetzende Myogenese quergestreifter Muskulatur ließ sich durch eine sich teilende Zelle in der Außenschicht kennzeichnen (Abbildung 5.4c,d). Vorläuferzellen der quergestreiften Muskulatur ordneten sich in kleinen Gruppen – umgeben von glatter Muskulatur und einer gemeinsamen Basallamina – primär in der äußeren Schicht an. Vereinzelt waren diese auch in der inneren Schicht zu finden. Sie zeichneten sich durch einen erhöhten
5.1 Entwicklung der Ösophagusmuskulatur bei Shh(+/+) Kontrollmäusen 22 Anteil an Zellorganellen (Mitochondrien, Golgi-Apparat) aus (Abbildung 5.4b). Teil- weise wiesen sie schon erste quergestreifte Myofilamente auf, während der Zellkern noch mittelständig lag (Abbildung 5.5). Die innere Muskelschicht setzte sich überwie- gend aus glatten Muskelzellen zusammen, deren Quantität nach kranial abnahm. Die glatten Muskelzellen beinhalteten zahlreiche Zellorganellen (Mitochondrien, rER, Golgi-Apparat), welche sich im gesamten Zytoplasma verteilten. Myofilamente waren bereits deutlich ausgeprägt. Die Zellkörper waren teils rund, teils schon spindelförmig und parallel zueinander angeordnet. Als weiteres Element waren vereinzelte Ansamm- lungen mesenchymaler Zellen zwischen den muskulären Schichten zu erkennen. Dar- über hinaus traten Mitosen überwiegend in der äußeren Muskelschicht und Apoptosen in beiden Schichten auf (Abbildung 5.6). Im zervikalen Abschnitt verlief die Muskulatur im Vergleich zum abdominalen Ösopha- gus überwiegend in umgekehrter Weise: Außen in einer Längsmuskel- und innen in einer zirkulären Muskelschicht (Abbildung 5.8c). Die vorhandene Muskulatur setzte sich primär aus quergestreiften Muskelzellen zusammen, welche in ihrer Entwicklung meist noch unreif waren. Dies zeichnete sich v.a. dadurch aus, dass die Zellkerne nur teilweise vollkommen randständig lagen und ein Zusammenschmelzen zu multinukle- ären Zellen (MF) nur partiell stattgefunden hatte (Abbildung 5.8b,d). Sarkomere hatten sich bereits deutlich in Ketten angeordnet und zeigten ausgeprägte Z-Streifen (Abbildung 5.8a,d). Satellitenzellen konnten vereinzelt dokumentiert werden. Diese waren von einer eigenen Basallamina umschlossen (Abbildung 5.7a,b). Weiterhin be- fanden sich Gruppen von Vorläuferzellen (MNC-I) zwischen den multinukleären Mus- kelzellen. (Abbildung 5.8b,d). Zwischen der äußeren und inneren Muskelschicht lagen Bindegewebszellen, die sich durch ein vermehrtes Vorkommen von rauem endoplas- matischem Retikulum (rER) und Mitochondrien auszeichneten.
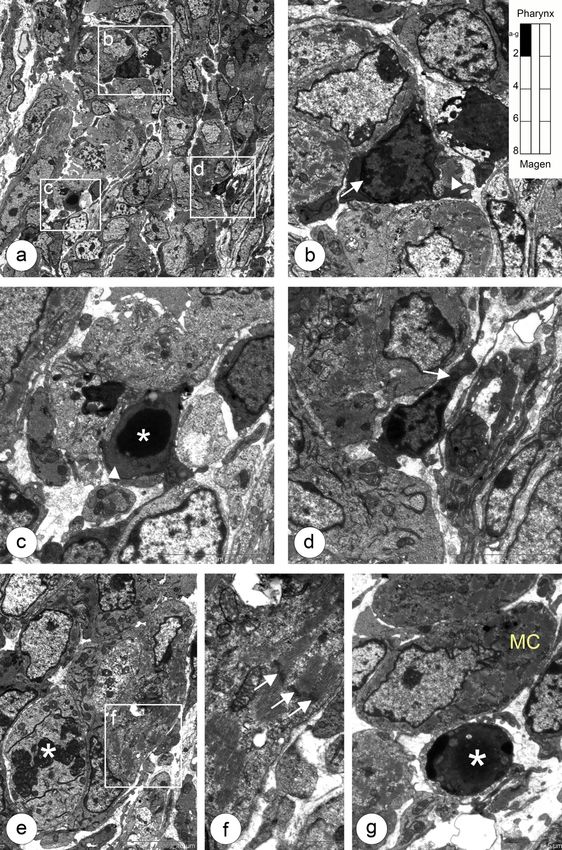
5.1 Entwicklung der Ösophagusmuskulatur bei Shh(+/+) Kontrollmäusen 23 Abbildung 5.2: Abdominaler Abschnitt mit glatter Muskulatur und undifferenzierten mononukleären Muskelvorläuferzellen a,b: Der Ösophagus ist von einem mehrschichtigen kubischen Epithel mit Mikrovilli (b, Pfeilspitzen) und wenigen Kinozilien (b, Pfeile) lumenwärts ausgekleidet. c,d: In der Übersicht zeigt sich eine schmale Außen- und eine breite Innenschicht der Muskulatur, dazwischen ein Ganglion des Plexus myentericus. e,f: Glatte Muskelzellen mit typischen morphologischen Kennzeichen: Myofila- mente (f, Stern) mit dense bands (f, Pfeil) im Zytoplasma. g,h: Zwischen den glatten Muskelzellen (h, Sterne) befinden sich mononukleäre Zellen vom Typ I (h, MNC I).
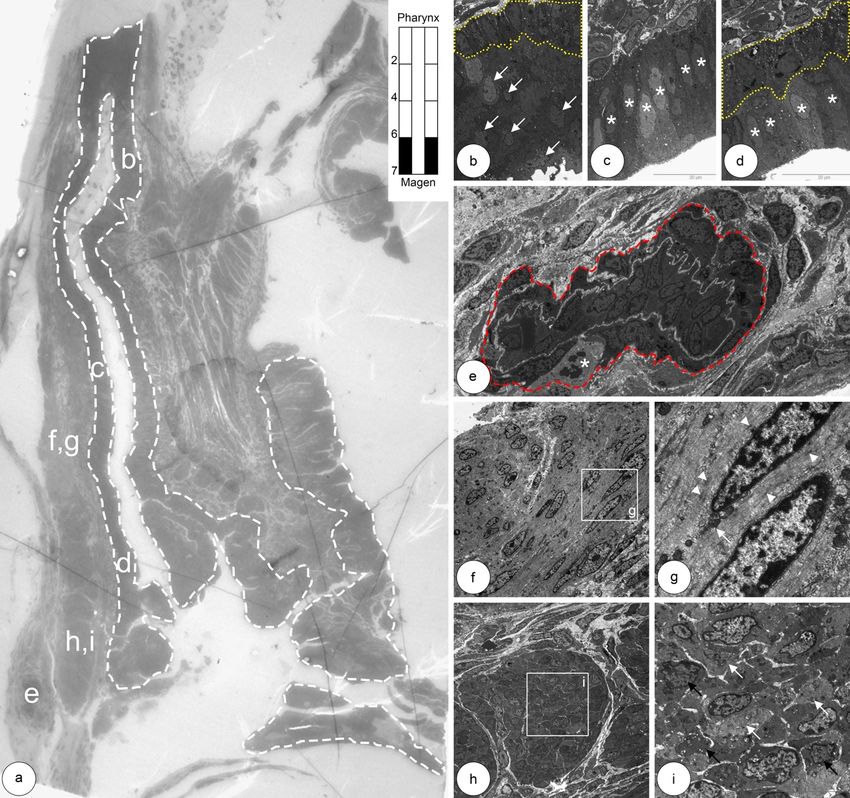
5.1 Entwicklung der Ösophagusmuskulatur bei Shh(+/+) Kontrollmäusen 24 Abbildung 5.3: Glatte Muskelzellen im unteren thorakalen Abschnitt des Ösophagus. a,b: In der Übersicht sieht man eine dünne äußere, neben einer dicken inneren Muskelschicht. Die Vergrößerung (b) zeigt ein zwischen beiden Muskelschichten lie- gendes Ganglion c: Neben überwiegend hellen glatten Muskelzellen (L-SMC) sind wenige dunklere glatte Muskelzellen (D-SMC) zu erkennen (Pfeil). d: In der äußeren Muskelschicht kommen Kapillaren vor. Die Erythrozyten ähneln mit ihrem dunklen Zellleib apoptotischen Zellkernen (Stern). e,f: Die Zellorganellen (f, Pfeile) der glatten Muskelzellen ordnen sich überwiegend in der Peri- pherie des Zytoplasmas an.
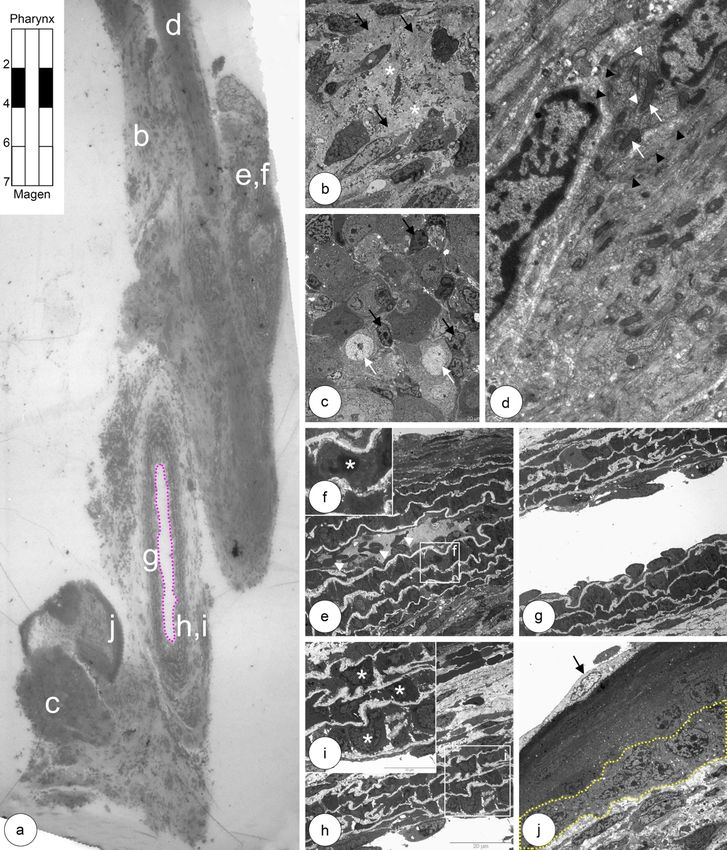
5.1 Entwicklung der Ösophagusmuskulatur bei Shh(+/+) Kontrollmäusen 25 Abbildung 5.4: Beginn der Myogenese und mononukleäre Muskelvorläuferzellen im oberen thorakalen Ösophagus. a: Zwi- schen einer dünnen Außen- und einer breiten Innenschicht der Tunica muscularis liegt ein Ganglion (Stern). b: Muskelvorläufer- zelle (MNC I) mit heterochromatischem Zellkern (Stern), geringer Zytoplasmaanteil mit enthaltenen Mitochondrien (Pfeil) und Golgi-Apparat (Pfeilspitze) sind deutlich erkennbar. c,d: Die erste sich teilende Muskelvorläuferzelle (MNC II) in der Außenschicht kennzeichnet den Beginn der einsetzenden Myogenese quergestreifter Muskelzellen.
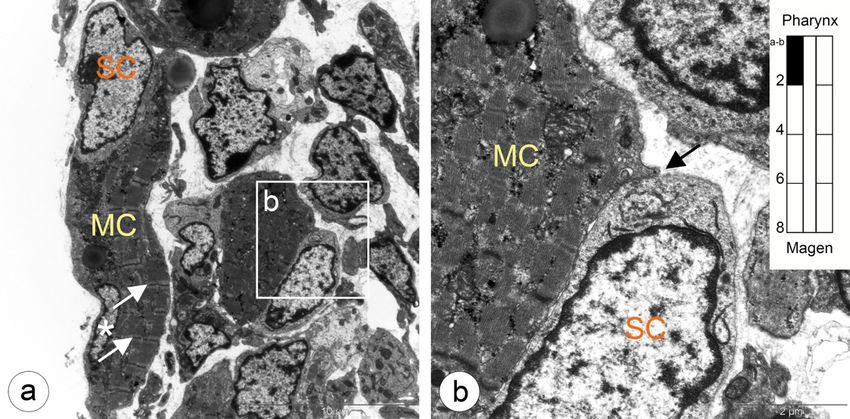
5.1 Entwicklung der Ösophagusmuskulatur bei Shh(+/+) Kontrollmäusen 26 Abbildung 5.5: Übergangszone im zervikalen Ösophagus. Mononukleäre Muskelzellen und apoptotische glatte Muskelzel- len. a: Erste auftretende Gruppe mononukleärer Zellen vom Typ II (rote Sterne) und Myozyten (gelbe Sterne) zwischen glatten Muskelzellen in der äußeren Muskelschicht. b: Ausschnitt aus a, mit bereits deutlich ausgeprägte Myofilamentbündel (weiße Pfeile) in den Myozyten. c: Übersichtsaufnahme der äußeren (links) und inneren (rechts) Muskulaturschicht. Zwischen beiden Schichten entwickelt sich Nervengewebe des Plexus myentericus (weißer Stern). d: Apoptotische glatte Muskelzelle mit konden- siertem Chromatin (Stern) und noch vorhandene Mitochondrien (Pfeilspitzen). e: Eine Gruppe mononukleärer Zellen Typ II (rote Sterne) und Myozyten mit Myofilamentbündeln (gelbe Sterne) in der Außenschicht aus c. f: Vergrößerte Darstellung der Myofila- mentbündel (Pfeile) aus e. g,h: Myozyt mit Myofilamenten (h, Pfeilspitzen) umgeben von glatten Muskelzellen in der Außenschicht.
Sie können auch lesen

















































