Vorsicht, Verwundung! Reizweiter lei- tung mit mechanosensitiven Proteinen
←
→
Transkription von Seiteninhalten
Wenn Ihr Browser die Seite nicht korrekt rendert, bitte, lesen Sie den Inhalt der Seite unten
601
Pflanzliche Wundsignalübertragung
Vorsicht, Verwundung! Reizweiterlei-
tung mit mechanosensitiven Proteinen
ren. Dadurch wird die Nahrungsaufnahme
MICHAEL M. WUDICK und das Wachstum der Raupen beeinträch-
INSTITUT FÜR MOLEKULARE PHYSIOLOGIE, UNIVERSITÄT DÜSSELDORF tigt und die Pflanze als Nahrungsquelle
unattraktiver [3]. Damit diese Abwehrstoffe
Being sessile, plants are exposed to adverse stresses, including nicht nur im verwundeten Blatt, sondern in
wounding by insects. Albeit lacking experimental evidence, one der ganzen Pflanze hergestellt werden, sen-
hypothesis predicted involvement of hydro-electric signals in wound det die Pflanze Signale bis in unverwundete
Gewebeteile. Diese wundinduzierte Signal-
signaling. Now, we could show that the mechanosensitive anion
weiterleitung kann dabei auf unterschied-
channel MSL10 is necessary for wound-induced long-distance signal- liche Weise erfolgen.
ing in plants. By linking mechano-sensing, ion fluxes, membrane depo-
larization and electrical signal propagation, MSL10 might integrate Signalweiterleitung in Pflanzen:
hydraulic and electric wound signals. elektrisch, chemisch oder
hydraulisch?
DOI: 10.1007/s12268-021-1658-5
Die Ausbreitung des Wundsignals erfolgt
© Der Autor 2021
über das Leitgewebe der Pflanze, das den
Transport von Wasser und darin gelöster
Nährstoffe ermöglicht. Die Weiterleitung
ó Pflanzen sind im Laufe ihres Lebens- gasförmigen Methyljasmonats kann es über kann in Form elektrischer, chemischer oder
zyklus einer Vielzahl äußerer Faktoren aus- Spaltöffnungen im Blatt ausströmen und hydraulischer Signale erfolgen, oder einer
gesetzt, wie Klimaschwankungen oder dem Nachbarpflanzen vor anstehendem Befall Kombination daraus. Die Blätter der Modell-
Befall von Insekten und Schädlingen. Schät- durch Fressfeinde „warnen“ [2]. Zudem kann pflanze Arabidopsis thaliana (Ackerschmal-
zungsweise gehen weltweit jährlich bis zu Jasmonsäure die Produktion von toxischen wand) wachsen in Rosettenform und sind
20 Prozent der Ernteerträge durch Schäd- Substanzen veranlassen. Wenn Pflanzen bei- durch ihr sequenzielles Wachstum von „alt“
lingsbefall verloren [1]. Ein besseres Ver- spielsweise von Raupen angefressen werden, nach „jung“ klassifizierbar (Abb. 1). Zudem
ständnis über pflanzliche Abwehrstrategien produzieren sie bestimmte Hemmstoffe, die kann man anhand der Blattstellung identi-
gegen Insektenfraß ist deshalb nicht nur aus die Verdauungsenzyme der Raupen blockie- fizieren, welche Blätter über dasselbe Leit-
ökonomischer Sicht, sondern auch mit Blick
auf eine gesicherte Lebensmittelversorgung
hoch relevant. ˘ Abb. 1: Die Blätter
der Ackerschmal-
Pflanzliche Abwehrstrategien: vom wand wachsen spiral-
förmig und lassen
mechanischen Schutzschild bis zur
sich entsprechend
„chemischen Keule“ des Zeitpunkts ihres
Zur Abwehr von Fressfeinden wie Insekten Erscheinens von alt
haben Pflanzen verschiedene Strategien ent- (Blatt 1) nach jung
(Blatt 20) klassi-
wickelt. Als erster Schutz dienen oft äußere
fizieren. Lokale Ver-
Barrieren, z. B. in Form von Borke, Stacheln wundung, z. B. durch
oder wachsbeschichteten Blättern. Wird die- Raupen (Blatt 8), initi-
se Barriere durchbrochen, kann die Pflanze iert elektrische, che-
durch gezielte Hemmstoffproduktion verhin- mische und hydrauli-
sche Signale, die sich
dern, dass Schädlinge sich auf andere Pflan-
in entfernte Pflanzen-
zenteile oder Nachbarpflanzen ausbreiten. teile (Blatt 13) aus-
Verwundung führt zur Produktion des breiten (Pfeile).
pflanzlichen Hormons Jasmonsäure, das die
Freisetzung von Abwehrstoffen und somit
die Immunabwehr der Pflanze ankurbelt.
Jasmonsäure kann dabei in verschiedenen
chemischen Varianten auftreten. In Form des
BIOspektrum | 06.21 | 27. Jahrgang
602 W I S S EN S CH AFT
A B C D
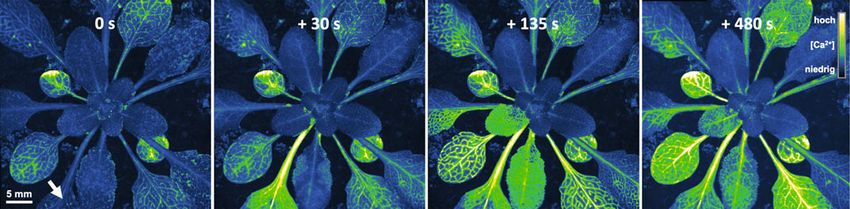
˚ Abb. 2: Visualisierung wundinduzierter Calciumsignale durch den fluoreszierenden Calciumsensor MCaMP6s. Mechanische Blattverwundung
(A, Pfeil) simuliert Insektenfraß und führt zum lokalen Anstieg der Calciumkonzentration [Ca2+] im Leitgewebe (B). Das Signal gelangt dann in
benachbarte, über gemeinsame Leitgewebe verbundene Blätter (C), bevor es sich auf die restliche Pflanze ausbreitet (D). © Bild: C. Sies, HHU.
Pflanzenteilen ein Unterdruck. Mechanis-
tische Modelle nahmen an, dass die Verwun-
dung des Leitgewebes zu einem rapiden
Druckanstieg in den Leitgewebezellen führt.
Die sich ausbreitende Druckdifferenz könnte
von mechanosensitiven Proteinen erfasst
und in elektrische Signale umgewandelt wer-
den. Daran beteiligte Proteine waren bislang
jedoch unbekannt.
Das mechanosensitive Protein MSL10
ist essenziell bei der
Wundsignalübertragung
Elektrische Signale lassen sich in Blättern als
Oberflächenpotenziale – die surface oder slow
wave potentials (SWPs) – detektieren. Die
Messung von SWPs erfolgt mithilfe von Elek-
troden, die auf Blattstielen von verwundeten
˚ Abb. 3: Repräsentative Spannungskurven von Kontrollpflanzen (grau) und msl10-Mutanten
(grün). Im abgebildeten Beispiel sind sowohl die Dauer (in Sekunden, s) als auch die Amplitude und unversehrten Blättern platziert werden
(in Millivolt, mV) des elektrischen Signals in der msl10-Mutante verändert, was z. B. zu einer ver- und die gesamte elektrische Aktivität des
minderten Produktion von Abwehrstoffen führen kann. darunterliegenden Gewebes erfassen. Basie-
rend auf diesen Informationen haben wir ein
Messverfahren optimiert, bei dem nach
gewebe zum direkten Signalaustausch befä- und führt zur Glutamat-abhängigen Aus- mechanischer Verwundung von Blatt 8 das
higt sind. Durch den Fressfeind oder die breitung des Ca2+-Signals [5, 6]. dabei generierte elektrische Wundsignal in
Verwundung freigesetzte Botenstoffe kön- Blatt 13 gemessen wird (Abb. 1). Kontroll-
nen an spezielle Rezeptoren im Leitgewebe Visualisierung wundinduzierter pflanzen sind durch SWPs mit charakteris-
binden. Einige dieser Rezeptoren fungieren Signalmoleküle tischer Dauer und Amplitude gekennzeich-
als Kanäle, die den Transport von Ionen Genetisch codierte, fluoreszenzbasierte Sen- net (Abb. 3, [6, 7]). Anhand systematischer
über biologische Membranen ermöglichen. soren für Glutamat und Ca2+-Ionen erlauben Screens von Mutanten, denen bestimmte
Ionenkanäle können das chemische Wund- es, die Entstehung und Ausbreitung des Gene fehlten, entdeckten wir, dass Pflanzen
signal in ein elektrisches Signal umwan- Wundsignals in Echtzeit quantitativ zu ana- ohne den mechanosensitiven Anionenkanal
deln, verstärken und an Nachbarzellen lysieren (Abb. 2). Vorangegangene Experi- MSL10 signifikant verkürzte SWPs aufwie-
übermitteln. Glutamat ist eines der am bes- mente zeigten, dass bestimmte GLRs essen- sen [10]. Zudem kam es in diesen Pflanzen
ten erforschten wundinduzierten Signalmo- ziell für die Ausbreitung wundinduzierter zu einer Verringerung des wundinduzierten
leküle. Ähnlich wie im menschlichen Ca2+- und elektrischer Signale über das Leit- Ca2+-Signals und einer verminderten Produk-
Gehirn, wo Glutamat als Neurotransmitter gewebe sind [7]. Über die Bedeutung des tion von Abwehrstoffen [10]. Die Ausbrei-
die neuronale Zell-Zell-Kommunikation Leitgewebes für die Ausbreitung eines tung des Botenstoffs Glutamat war hingegen
ermöglicht, verfügt das Leitgewebe von hydraulischen Wundsignals und die Kopp- nicht beeinträchtigt. Da das gleichzeitige
Pflanzen über Glutamatrezeptoren (GLRs), lung hydroelektrischer Signale wurde lange Entfernen der ebenfalls zur Wundsignalüber-
an die der Botenstoff binden kann [4]. Die spekuliert [8, 9]. Im Leitgewebe herrscht tragung benötigten GLRs zu keiner weiteren
Interaktion mit Glutamat erhöht die durch den kontinuierlichen Transpirations- Veränderung des SWPs führte, ist unsere
Calcium(Ca2+)-Transportaktivität der GLRs sog zwischen der Wurzel und oberirdischen Hypothese, dass entweder zwei parallele Sig-
BIOspektrum | 06.21 | 27. Jahrgang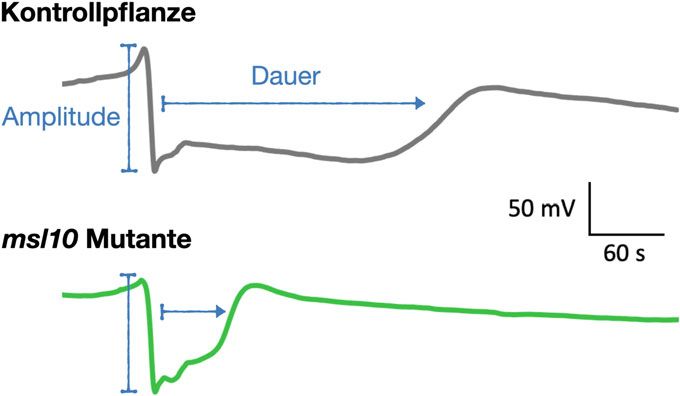
nalwege existieren oder dass sich MSL10 nummer 267205415 – SFB 1208 gefördert
und die GLRs in einem gemeinsamen Sig- und ich danke allen daran Beteiligten! ó
nalweg gegenseitig regulieren können
[10]. Literatur
[1] Deutsch CA, Tewksbury JJ, Tigchelaar M et al. (2018)
Increase in crop losses to insect pests in a warming cli-
Glutamat als Wundsignal oder mate. Science 361: 916–919
Nährstoff: Wie unterscheidet die [2] Farmer EE, Ryan CA (1990) Interplant communica-
tion: airborne methyl jasmonate induces synthesis of pro-
Pflanze? teinase inhibitors in plant leaves. Proc Natl Acad Sci USA
Anders als im tierischen System fungiert 87: 7713–7716
[3] Green TR, Ryan CA (1972) Wound-induced protein-
Glutamat in Pflanzen nicht nur als Boten- ase inhibitor in plant leaves: a possible defense mecha-
stoff, sondern auch als Metabolit, dessen nism against insects. Science 175: 776–777
[4] Alfieri A, Doccula FG, Pederzoli R et al. (2020) The
Konzentration ständigen Schwankungen structural bases for agonist diversity in an Arabidopsis
unterliegt. Um zwischen metabolischen thaliana glutamate receptor-like channel. Proc Natl Acad
Sci USA 117: 752–760
Schwankungen und verwundungsbeding- [5] Wudick MM, Portes MT, Michard E et al. (2018)
tem Anstieg der Glutamatkonzentration CORNICHON sorting and regulation of GLR channels
underlie pollen tube Ca2+ homeostasis. Science 360:
zu unterscheiden, bedarf es eventuell 533–536
einer weiteren Stellschraube. Diese Funk- [6] Toyota M, Spencer D, Sawai-Toyota S et al. (2018)
Glutamate triggers long-distance, calcium-based plant
tion übernimmt möglicherweise MSL10. defense signaling. Science 361: 1112–1115
In unserem Modell führt wundinduzierter [7] Mousavi SAR, Chauvin A, Pascaud F et al. (2013)
GLUTAMATE RECEPTOR-LIKE genes mediate leaf-to-leaf
Druckabfall im Blatt-Leitgewebe zur wound signalling. Nature 500: 422–426
MSL10-Aktivierung und dem Transport [8] Malone M (1996) Rapid, long-distance signal trans-
mission in higher plants. Adv Bot Res 22: 163–228
von Anionen über die Zellmembran. Die [9] Farmer EE, Gasperini D, Acosta IF (2014) The
so hervorgerufene Membrandepolarisie- squeeze cell hypothesis for the activation of jasmonate
synthesis in response to wounding. New Phytol 204:
rung könnte der Pflanze helfen, den 282–288
gleichzeitigen Anstieg der Glutamatkon- [10] Moe-Lange J, Gappel NM, Machado M et al. (2021)
Interdependence of a mechanosensitive anion channel
zentration als Wundsignal zu erkennen and glutamate receptors in distal wound signaling. Sci
und GLR-vermittelte Abwehrreaktionen Adv 7: eabg4298
[11] Hansen KB, Yi F, Perszyk RE et al. (2018) Structure,
einzuleiten (z. B. durch Jasmonsäure-Pro- function, and allosteric modulation of NMDA receptors. J
duktion). In der Tat existiert eine ähnliche Gen Physiol 150: 1081–1105
Form der simultanen Ko-Aktivierung bei
Funding note: Open Access funding enabled and organized by Projekt
einigen tierischen Glutamatrezeptoren, DEAL.
Open Access: Dieser Artikel wird unter der Creative Commons
bei denen die Interaktion der Rezeptoren Namensnennung 4.0 International Lizenz veröffentlicht, welche die
Nutzung, Vervielfältigung, Bearbeitung, Verbreitung und Wiedergabe in
mit Glutamat nur bei gleichzeitiger Mem- jeglichem Medium und Format erlaubt, sofern Sie den/die ursprünglichen
Autor(en) und die Quelle ordnungsgemäß nennen, einen Link zur Creative
brandepolarisierung zu einer Signalwei- Commons Lizenz beifügen und angeben, ob Änderungen vorgenommen
wurden. Die in diesem Artikel enthaltenen Bilder und sonstiges
terleitung führt [11]. Indem wir die an der Drittmaterial unterliegen ebenfalls der genannten Creative Commons
Lizenz, sofern sich aus der Abbildungslegende nichts anderes ergibt.
pflanzlichen Wundsignalweiterleitung Sofern das betreffende Material nicht unter der genannten Creative
Commons Lizenz steht und die betreffende Handlung nicht nach
beteiligten Prozesse und Komponenten gesetzlichen Vorschriften erlaubt ist, ist für die oben aufgeführten
Weiterverwendungen des Materials die Einwilligung des jeweiligen
besser verstehen, können wir den Pflan- Rechteinhabers einzuholen. Weitere Details zur Lizenz entnehmen Sie bitte
der Lizenzinformation auf http://creativecommons.org/licenses/by/4.0/
zen möglicherweise helfen, Bedrohungen deed.de.
schneller zu erkennen und abzuwehren.
Zukünftig kann so eventuell Ernteausfäl-
len durch Schädlingsbefall gezielter ent- Korrespondenzadresse:
gegengewirkt werden. Dr. Michael M. Wudick
Institut für molekulare Physiologie
Heinrich-Heine-Universität Düsseldorf
Danksagung
Universitätsstraße 1
Dieses Projekt wurde durch die Deutsche D-40225 Düsseldorf
Forschungsgemeinschaft (DFG) – Projekt- wudick@hhu.de
AUTOR
Michael M. Wudick
2001–2006 Biochemiestudium an der Universität Potsdam. 2007–2010 Pro-
motion am INRA/SupAgro/CNRS/Université de Montpellier 2, Montpellier,
Frankreich. 2011–2014 Postdoc am Instituto Gulbenkian de Ciência (IGC),
Oeiras, Portugal. 2014–2019 Research Associate an der University of Mary-
land, College Park, USA. Seit 2019 Akademischer Rat und Gruppenleiter am
Institut für molekulare Physiologie der Universität Düsseldorf.
BIOspektrum | 06.21 | 27. JahrgangSie können auch lesen