3.1 Körpereigene, elektromagnetische Biosignale
←
→
Transkription von Seiteninhalten
Wenn Ihr Browser die Seite nicht korrekt rendert, bitte, lesen Sie den Inhalt der Seite unten
3.1 Körpereigene, elektromagnetische Biosignale 3.1.1 Funktionsweise von Nervenzellen, zum Thema „Nervenreizleitung“ Aufbau der Nervenzelle Eine Nervenzelle (Neuron) ist eine auf Erregungsleitung spezialisierte Zelle, vgl. Bild 3.1/1. Das menschliche Gehirn besteht schätzungsweise aus 1011 bis 1012 Nervenzellen. Eine typische Säugetier-Nerven- zelle ist aus Dendriten, dem Zell- körper und einem Axon (Zellfort- satz) aufgebaut. Dieser Zellfort- satz kann sehr lang sein und er- möglicht eine Erregungsleitung über weite Strecken (bis zu 1,5 m !!). Dabei läuft ein elektrisches Signal durch das Axon. Es wird erzeugt, indem bestimmte Ionen gezielt durch die Zellmembran durchgeschleust werden. Das Axonende steht über Synapsen, an denen das Signal chemisch (seltener elektrisch) weitergege- ben wird, mit anderen Nervenzel- len oder Empfängerzellen in Ver- bindung. Bild 3.1/1: Nervenzelle Der Zellkörper (Soma) enthält neben dem Zellkern diverse Organellen. Das Soma hat in Abhängigkeit vom Nervenzelltypus eine Größe zwischen 5 und mehr als 100 µm. Es produ- ziert die Neurotransmitter für die Reizweiterleitung. Die an Dendriten eintreffenden Signale werden im Soma weiterverarbeitet. Das geschieht durch räumliche und zeitliche Summation von Änderungen des Membranpotentials. Vom Soma wachsen Fortsätze aus, die Dendriten. Das sind feine plasmatische Verästelun- gen des Zellkörpers, die über Synapsen den Kontakt zu anderen Nervenzellen herstellen. Sie empfangen Aktionspotentiale von anderen Neuronen durch deren Axone. Der Dendriten- baum einer einzigen menschlichen Nervenzelle kann mit 100.000 bis 200.000 Fasern ande- rer Neuronen im Kontakt stehen (die Potenziale aller Eingänge werden summiert). Dies ermög- licht eine extrem hochgradige Parallelisierung der Signalisierung zwischen den Nervenzellen. Die elektrischen Potentiale zufolge der Reize aller „Eingänge“ werden summiert, sobald das Summenpotential im Zellinneren zu positiv wird, reagiert die Zelle mit einem elektrischen Spannungsimpuls („Spike“) An das Soma angesetzt ist der Axonhügel. Das Schwellenpotential des Axonhügels ist stark reduziert. Es wirkt dadurch als Initialsegment. Von hier aus werden die Aktionspotentiale an das Axon weitergeleitet. Durch das niedrige Schwellenpotential und die Nähe des Axon- hügels zum Zellkörper ist sichergestellt, dass bei einer Erregung der Zelle das Aktionspoten- tial nur an einem Ort entsteht und weitergeleitet wird. Das ist wichtig für die gerichtete Erre- gungsleitung, da eine Nervenzelle bei ausreichender Reizintensität an jeder Stelle erregt werden kann und Aktionspotentiale in jede Richtung leitet. Das Axon ist ein langer Fortsatz der Nervenzellen, der am Axonhügel entspringt. Es ist in der Regel mehr oder weniger stark verzweigt und mündet in synaptische Endigungen (Sy- napsen). Ein Axon kann je nach Typ der Nervenzelle von 1 µm bis 1 m und länger (!!) sein. (Umgangssprachlich „Nerven“). Es weist eine Dicke von 0,5 bis 10 μm auf. Das Axon ist von mehreren aufeinander folgenden Myelinscheiden umhüllt. (Schutzhülle, auch elektrische Dr. Garn, Ergänzungen Dr. Ehrlich-Schupita Elektromagnetische Felder und der Mensch Seite 3.1.1
Isolation). Das Axon ist zuständig für die Übertragung des Aktionspotentials einer Nervenzel- le und leitet dieses zu den Synapsen und damit an andere Nervenzellen weiter. Des Weite- ren wandern die Stoffe, die im Soma gebildet werden (Neurotransmitter, Enzyme), durch das Axon zur Synapse. Auch in umgekehrter Richtung, also von der Synapse in Richtung Zell- körper, findet Stofftransport statt. Die Synapse stellt eine Schnittstelle zwischen Neuronen dar, über die Informationen che- misch übertragen werden können, vgl. Bild 3.1/2. Am Ende des Axons befinden sich soge- nannte Endknöpfchen, welche die Kontaktstelle zwischen zwei Nervenzellen bilden. Die Sy- napse enthält die präsynaptische Membran der "Senderzelle", welche die Neurotransmitter nach einer vorausgegangenen Erregung in den synaptischen Spalt entleert. Ihr gegenüber liegt die postsynaptische Membran der "Empfängerzelle", welche mit Rezeptoren bestückt ist, an der die Neurotransmitter binden, um dort sogenannte Ionenkanäle zu öffnen. Zwi- schen den beiden Membranen liegt der etwa 30 nm breite synaptische Spalt, welcher mit einer plasmatischen Lösung aufgefüllt ist. Ein Neuron bildet 1 bis 200.000 Synap- sen mit anderen Neuronen. Das mensch- liche Gehirn weist insgesamt etwa eine Billiarde Synapsen auf. Synapsen von Nervenzellen verbinden sich auf diese Weise indirekt untereinander zu einem neuronalen Netzwerk. Bild 3.1/2: Chemische Synapse (be- stehend aus: Axon Terminal mit präsy- naptischer Membran der „Senderzelle“ = „Endknöpfchen“ eines Axons // synap- tischem Spalt // Dendritenfortsatz mit postsynaptischer Membran der „Emp- fängerzelle“) Elektrochemische Funktionsweise der Nervenzelle, Natrium-Kalium-Pumpe Im Inneren des Axons wie auch außerhalb der Membran existieren Anionen und Kationen. Sie bestimmen Ladung und elektrische Leitfähigkeit des Mediums. Die Biomembran des Axons mit einer Dicke von nur etwa 10 nm !!!!! sorgt nun dafür, dass zwischen Innen und Außen verschiedene Konzentrationen der Ionen bestehen. Das bedeutet, dass an der Au- ßenwand des Axons eine andere elektrische Ladung anliegt als innen („Polarisation“, Ruhe- potential). Zwischen den beiden Oberflächen der Membran besteht also eine elektri- sche Potentialdifferenz. Die ungleiche Verteilung der Ionen innerhalb und außerhalb der Zelle sowie selektive Per- meabilitäten der Membrankanäle erzeugen das Membranpotential: Während die Zelle viele K+-Ionen und wenige Na+- und Cl--Ionen enthält, sind außerhalb der Zelle die Na+- und Cl-- Konzentrationen hoch und die K+-Konzentrationen niedrig. Aufgrund der unterschiedlichen Ionenkonzentrationen diffundieren die in der Zelle hoch konzentrierten K+-Ionen durch die K+- Kanäle hinaus. Dabei entfernen sie positive Ladungen aus der Zelle, das Zellinnere wird negativ aufgeladen. Dies wirkt der Diffusion der K+-Ionen entlang ihres Konzentrationsgra- dienten entgegen. Die negative Aufladung des Zellinneren kommt zum Stillstand, wenn sie den „Diffusionsdruck“ des Konzentrationsgradienten für K+ gerade aufhebt. Im Zustand des Fließgleichgewichts passieren auswärts und einwärts gleich viele K+-Ionen die Kanäle. Die- ses Fließgleichgewicht ist ein „dynamisches Gleichgewicht“ und benötigt ständig Energiezu- fuhr zum Organismus, z.B. durch die Aufspaltung von Zucker. Dr. Garn, Ergänzungen Dr. Ehrlich-Schupita Elektromagnetische Felder und der Mensch Seite 3.1.2
Für Körpertemperatur (T = 310 K) und ein Verhältnis der Ionenkonzentrationen von [K+]i/[K+]a = 39 wird daraus z. B. das K+-Gleichgewichtspotential von -97 mV. Die Herstellung und Aufrechterhaltung dieses Potentials erfolgt durch die Natrium-Kalium-Pumpe. Eine Nervenzelle erhält ein Signal, indem Neurotransmitter an spezielle Rezeptoren in der postsynaptischen Membran in den Dendriten oder auch des Somas der zu erregenden Zelle anbinden. Ist die Erregung auf diese Weise übertragen, wird sie über die Dendriten an das Soma der Nervenzelle und von dort zum Axonhügel weitergeleitet. Jede der eingehenden Depolarisationen an den verschiedenen Synapsen der Nervenzelle verändert dabei das Membranpotential an der axonalen Membran. Je näher eine Synapse am Soma ansetzt, desto stärker ist ihr Einfluss auf die Nerven- zelle, je länger der Weg, den die Erregung zurücklegen muss, desto schwächer wird der Einfluss. Eine stärkere Reizung eines Dendriten (Fortsatzes vom Soma, der Ner- venzelle) resultiert also in einer stärkeren Depolarisierung. Nahezu gleichzeitig einlau- fende Reize addieren sich in ihrer Wirkung, was bedeutet, dass sich innerhalb der Zelle und am Axonhügel ein Erregungspotential aufbaut (Summation). Wird am Axonhügel das Schwellenpotential erreicht und überschritten, so wird durch Depo- larisation ein Aktionspotential entlang des Axons ausgelöst. Bild 3.1/3 zeigt den Verlauf der Ionenverteilung. Bild 3.1/3: Verlauf der Ionenverteilung während des Aktionspotentials, Schritte 1) bis 6) Blau: Na-Ionen, Rot: K-Ionen Schritt 1) Ruhepotential, Ionenverteilung hält das Potential im Zellinneren auf -70mV Schritt 2) Eine Potentialänderung pflanzt sich über ein Axion fort und gelangt zum Synapsenspalt, Neurotransmit- ter veranlassen eine Öffnung von Na-Kanälen, Na-Ionen strömen in die Zelle, die Innenseite der Zellmembran wird positiver in Vergleich zum Ruhepotential Schritt 3) eine Schwelle ist überschritten, die Na-K-Pumpe kann den Na-Ionenüberschuss nicht schnell genug ausgleichen, das Potential im Zellinneren wird positiv, die sogenannte DEPOLARISATION beginnt, es gibt einen elektrischen Spannungspuls Dr. Garn, Ergänzungen Dr. Ehrlich-Schupita Elektromagnetische Felder und der Mensch Seite 3.1.3
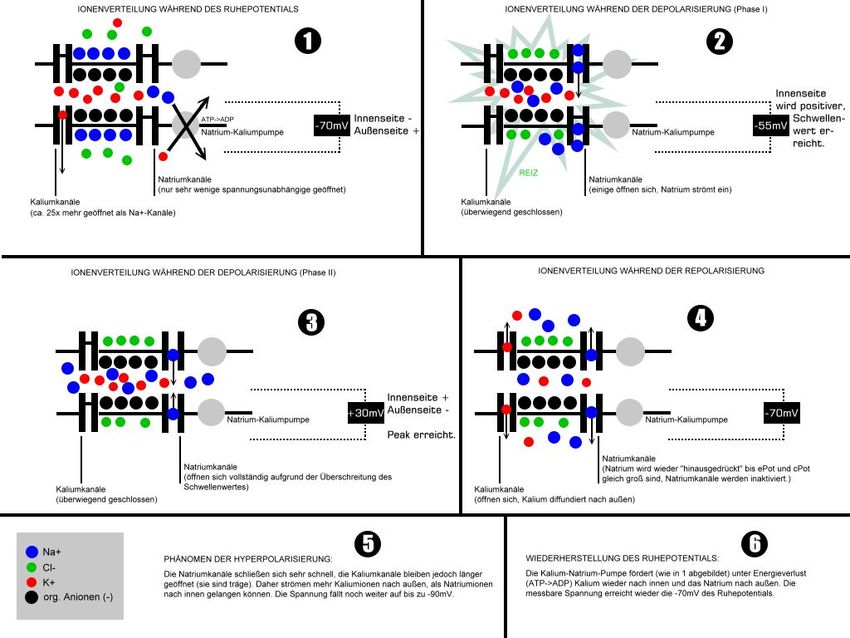
Schritt 4) bis 6) die K-Kanäle öffnen sich und nach etwa 10 ms ist das Ruhepotential wieder hergestellt Die spezifischen Ionenkanäle für Natrium- und Kalium-Ionen öffnen sich in Abhängigkeit vom Membranpotential, d. h. sie sind spannungsaktiviert. (Vergleiche mit der Funktionsweise eines Feldeffekttransistors). So ist beispielsweise ein spannungsabhängiger Natriumkanal beim Ru- hemembranpotential geschlossen und aktivierbar. Bei Depolarisation über einen kanalspezi- fischen Wert wird der Kanal durchlässig für Ionen und geht in den Zustand offen über. Der Kanal bleibt aber trotz anhaltender Depolarisation nicht offen, sondern wird innerhalb weni- ger Millisekunden unabhängig vom Membranpotential wieder geschlossen. Das geschieht meist durch einen im Zytoplasma liegenden Teil des Kanalproteins, die Inaktivierungsdomä- ne, die sich gleich einem „Stöpsel“ in den Kanal setzt und diesen verstopft. Nicht alle Kanäle öffnen sich gleichzeitig bei ein und demselben Wert des Membranpotentials. Vielmehr ist die Wahrscheinlichkeit eines Kanals, in einen bestimmten Zustand überzugehen, spannungsab- hängig. Aus der rein statistischen Verteilung stellt sich ein Gleichgewicht ein, so dass eine größere Zahl von Kanälen in der Summe sehr gut das beschriebene Modell erfüllt. Auch ist der Zeitaufwand, um von einem Zustand in den anderen überzugehen, kanalspezi- fisch. Im geschilderten Natriumkanal läuft die Konformationsänderung von geschlossen nach offen in weniger als einer Millisekunde ab, während ein vergleichbarer Kaliumkanal Zeit in der Größenordnung von 10 ms benötigt. (Elektrisches Aktionspotential pflanzt sich entlang des Axons fort) Wandert nun ein Aktions- potential durch die Änderung des Konzentrationsgefälles der Ionen innerhalb des Axons am Axon entlang bis zum Endknöpfchen, so stößt dieser elektrische Impuls am Ende des Axons an eine Grenze, da eine Übertragung des elektrischen Signals durch den synaptischen Spalt zwischen den beiden Zellen nicht möglich ist. Der Reiz wird chemisch über die Synapsen weitergeleitet und mittels Neurotransmitter auf eine andere Zelle übertragen. Sobald ein Ak- tionspotential ausgelöst wurde, braucht die Zelle Zeit (Refraktärphase), um das Membranpo- tential wiederaufzubauen (Repolarisation), vgl. Bild 3.1/4. Während der Refraktärphase (Repolarisations- bzw. Erholungsphase) kann kein neues Aktionspo- tential ausgelöst werden. Wenn also von nachei- nander einlaufenden Reizen einer so stark ist, dass die Zelle ein Aktionspotential bildet und der nach- folgende Reiz während der Refraktärzeit einläuft, bildet die Zelle dafür kein neues Aktionspotential aus. Je mehr Aktionspotentiale die Zelle pro Sekunde abfeuert (je nach Zelltyp bis zu 500 Mal), desto stärker ist der Reiz. Bild 3.1/4: Zeitverlauf des Aktionspotentials, die Pulsdauer ist hier einige ms, kann bei an- deren Nervenzellen deutlich länger sein (z.B. für die Steuerung des Herzmuskels) Der Spannungsimpuls (Aktionspotential) ist die Antwort der Zelle auf den Empfang eines Reizes, diese Antwort wird an andere verknüpfte Zellen weiter gegeben, diese wiederum „überlegen“ sich gewissermaßen was sie damit anfangen sollen, sie „lernen“. Signalcodierung („Puls-Zahl / Pulsfrequenz-Codierung“) Alle Informationen, die die Nerven übertragen sollen, müssen als Frequenz von Aktionspo- tentialen „codiert“ werden. Langdauernde Depolarisationen lösen Serien von Aktionspoten- tialen aus, in denen die Frequenz der Impulse zur Stärke der Depolarisation proportional ist. Dr. Garn, Ergänzungen Dr. Ehrlich-Schupita Elektromagnetische Felder und der Mensch Seite 3.1.4
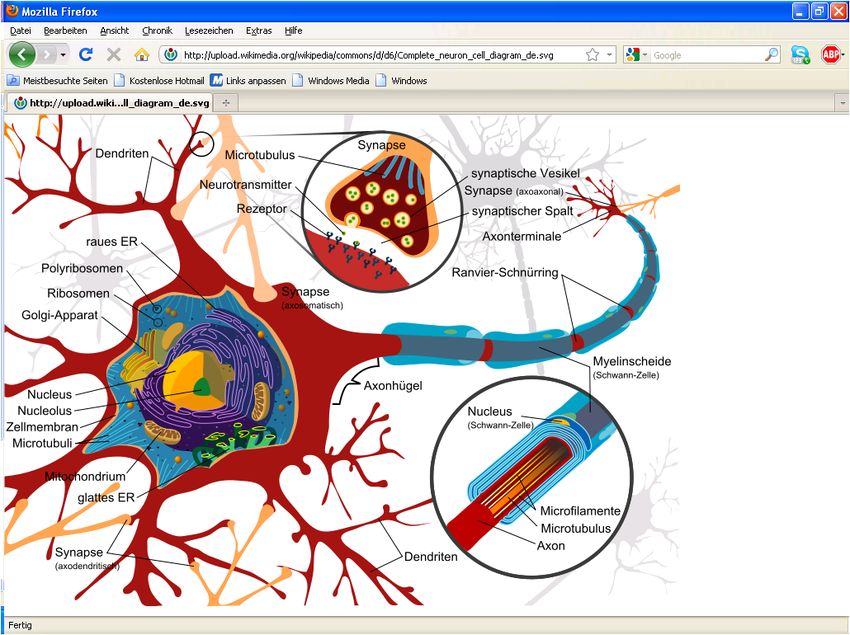
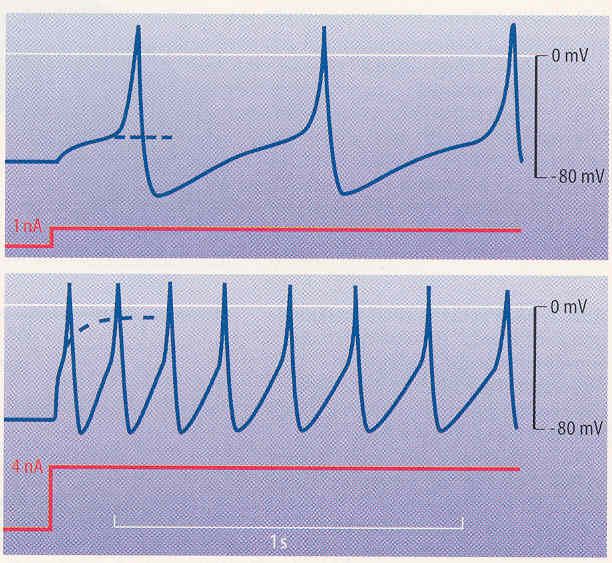
Bild 3.1/5 zeigt, wie eine Zelle auf das Einschalten eines Reizstromes von 1 nA oder von 4 nA antwortet. Der geringe Strom von 1 nA führt zu einer langsam ansteigenden, elektrotoni- schen Depolarisation, die in Fortsetzung der strichlierten Kurve ihren Endwert finden würde. Vor dem Einstellen des Endwertes erreicht jedoch die Depolarisation die Schwelle und löst ein Aktionspotential aus. Dieses hyperpolarisiert nach der Repolarisation über das Ruhepo- tential hinaus, dann folgt eine langsame Depolarisation, nach etwa 0,5 s wird die Schwelle erreicht und ein weiteres Aktionspotential ausgelöst. Dieser Zyklus kann sich wiederholen, solange der depolarisierende Strom fließt. Die Dauerdepolarisation wird somit in eine rhyth- mische Aktionspotentialauslösung mit etwa 2 Hz umgesetzt. Beim stärkeren Strom von etwa 4 nA erfolgt grundsätzlich das gleiche wie bei 1 nA, nur die Steilheit und die Amplitude der (strichlierten) Dauerdepolarisation sind größer und entspre- chend die Frequenz der erzeugten Aktionspotentiale höher: Sie liegt anfänglich bei 7 Hz und nimmt auf 4 Hz ab. Diese langsame Abnahme einer Frequenz bei gleichbleibendem Reiz- strom wird „Adaption“ genannt. Insgesamt ist also die Amplitude des Reizstromes bzw. der Depolarisation in entsprechende Aktionspotentialfrequenzen umcodiert worden. a) Der depolarisierende Strom von 1 nA in ein Neuron erzeugt ein elektrotonisches (durch gleichblei- benden Stromfluss ausgelöstes) Potential, das in eine Dauerdepolarisation von etwa – 20 mV münden würde (strichliert), wenn nicht die Schwelle zur Auslösung eines Aktionspotentials über- schritten würde. Die Aktionspotentiale wiederholen sich rhythmisch, solange der Stromfluss an- hält. b) Der Strom von 4 nA erzeugt ein elektrotonisches Potential, das fast 0 mV erreichen würde (strich- liert). Es wird jedoch eine Serie von Aktionspotentialen mit einer höheren Wiederholrate ausgelöst als bei 1 nA. Bild 3.1/5: Rhythmische Impulsbildung, ausgelöst durch einen andauernden Reizstrom Dr. Garn, Ergänzungen Dr. Ehrlich-Schupita Elektromagnetische Felder und der Mensch Seite 3.1.5
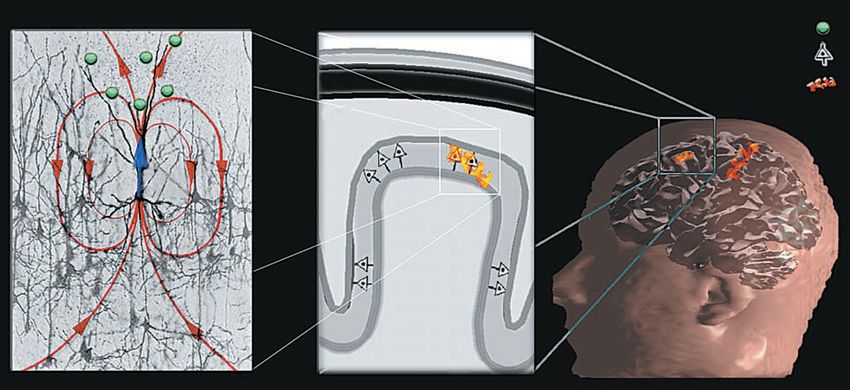
3.1.2 Elektrische und magnetische Felder im Gehirn Entstehung makroskopisch messbarer Felder In der Großhirnrinde (graue Masse) sitzen kortikale Pyramidenzellen (besonders große Ner- venzellen, die beim Menschen 85 % aller Nervenzellen ausmachen). Sie haben verlängerte Dendriten, die systematisch säulenförmig aneinander ausgerichtet sind und im rechten Win- kel zur Kortexschichte stehen, vgl. Bild 3.1/6 (Mitte). Diese räumliche Anordnung und die simultane Aktivierung großer Populationen dieser Zellen erzeugen die räumliche und zeitli- che Superposition der elementaren Aktivität jeder Zelle. Das Feld eines einzelnen Neurons ist zu schwach, um messbar zu sein. Die im EEG detektierten Signale stammen vielmehr aus Volumenbereichen mit untereinander verbundenen Neuronen, deren synaptische Strö- me zeitlich korreliert sind. Dies resultiert in einem Stromfluss und entsprechenden Potentia- len, die mit der Elektro-Enzephalographie (EEG) und der Magneto-Enzephalographie (MEG) gemessen werden können. Auch die im EEG auf der Kopfhaut messbaren Potentialun- terschiede zufolge der Superposition von Einzelsignalen in den Makro-Volumsbereichen sind sehr klein und liegen in der Größenordnung von 5µV bis 100 µV! Die kortikalen Nervenzellen bilden funktionale Netzwerke, die über verschiedene Gehirnregi- onen verteilt sein können. Sie sind die mutmaßlichen Generatoren der EEG u. MEG Signale. Stimulierende postsynaptische Potentiale entstehen am Scheitel des Dendritenbaums einer kortikalen Pyramidenzelle und triggern die Erzeugung eines Stromflusses durch das Gehirn- volumen von der nicht erregten Membran des Somas zu den Enden des Dendritenbaums. Der Strom fließt durch den Dendriten (Primärstrom im Bild 3.1/6 links, in blau) und verteilt durch den Kortex zurück (Sekundärströme, in rot). Bild 3.1/6: Netzwerke kortikaler Neuronengruppen als Erzeuger der EEG-Signale Näherungsweise Berechnung elektrischer und magnetischer Felder im Kopf Die gesamte Stromdichte im Kopf besteht aus zwei Komponenten: Einem primären (oder treibenden) Stromfluss JP(r´), der der tatsächlichen Aktivität der Neuronen entspricht, und dem im Volumen fließenden Strom JV(r´), der von der Wirkung des elektrischen Feldes auf die extrazellulären Ladungsträger herrührt. J(r´) = JP(r´) + JV(r´) = JP(r´) + s(r´).E(r´) = JP(r´) - s(r´).ÑV(r´), (3.1-1) wobei s … Leitfähigkeit des Gewebes im Kopf V … elektrisches Potential E = - grad V Die Divergenz des Gesamtstromes muss (im quasistatischen Fall, der hier gegeben ist) null sein (1. Maxwellgleichung mit ¶D/¶t = 0, div rot = 0 à div J = 0). Also DV = ÑJP/s (die zweite räumliche Ableitung des Potentials) (3.1-2) Dr. Garn, Ergänzungen Dr. Ehrlich-Schupita Elektromagnetische Felder und der Mensch Seite 3.1.6
Die Lösung dieser Poissongleichung führt auf das elektrische Potential am Aufpunkt r. Inte-
griert wird über das Volumen G, das die Quelle(n) enthält. Für den unendlich ausgedehnten,
homogenen Raum würde (3.1-3) gelten:
1 r - r´
4ps ò
V (r ) = J P
(r´) 3
dr´ (Lösungsansatz für das Potential) (3.1-3)
G r - r´
Betrachten wir die Quellen, die sich in einem vergleichsweise kleinen Volumenbereich am
Ort r0 befinden. Der Beobachtungspunkt r ist ein Stück davon entfernt. Die primäre Strom-
dichte JP kann in diesem Fall gut durch einen äquivalenten „Stromdipol“ als Punktquelle re-
präsentiert werden.
JP(r´) = m.d(r´-r0), (3.1-4)
wobei d(r) … Diracfunktion
ò
Moment m º J (r´)dr´ .
P
(3.1-5)
Damit wird
1 r - r0
V (r ) = m (VGL.: PRECHTL KAP. 3.1)
4ps r - r0
3
Das inverse Problem führt auf die Berechnung der Dipolstromquellen, die die Potentiale an
der Kopfoberfläche erzeugen. Diese Potentiale werden im EEG gemessen.
Die Berechnung beinhaltet selbstverständlich Ungenauigkeiten:
Ø Die gemessenen Signale sind von Rauschen überlagert.
Ø Das Modell für die Kopplung zwischen den Stromquellen im Kortex und den gemessenen
Potentialen ist zu grob.
Ø Unterbestimmtheit bzw. Mehrdeutigkeit des inversen Problems bewirkt begrenzte Auflö-
sung in der Schätzung.
Dieses Verfahren wird zur Quellenanalyse eingesetzt. Es liefert sowohl in der Neurologie als
auch in der Gehirnforschung wertvolle Aufschlüsse über die Topographie der Gehirnaktivität.
Quellenanalyse (Low resolution brain electromagnetic tomography, LORETA)
Die primären Generatoren der im EEG registrierten Wellenvorgänge und Potentialschwan-
kungen sind synaptische Ströme. Diese Stromflüsse über die neuronalen Membranen fun-
gieren als winzige Stromquellen und –senken. Für jedes Neuron muss der gesamte Fluss
von Ladungen (hinein und hinaus) in Summe null ausmachen.
Bezogen auf die Schichtdicke haben die EEG-Messpunkte großen Abstand. Daher können
die Stromverteilungen innerhalb eines Stückes Kortex durch kleine, dipolförmige Stromquel-
len repräsentiert werden, die normal zur jeweiligen Kortexfläche ausgerichtet sind und deren
Stärke zeitlich variiert. Die Kopplung zwischen den Dipolstärken und den an der Kopfoberflä-
che gemessenen elektrischen Feldern kann damit einfach als Summe bzw. Integral über alle
Volumenbereiche berechnet werden. Mittels Quellenanalyse kann man aus den am Kortex
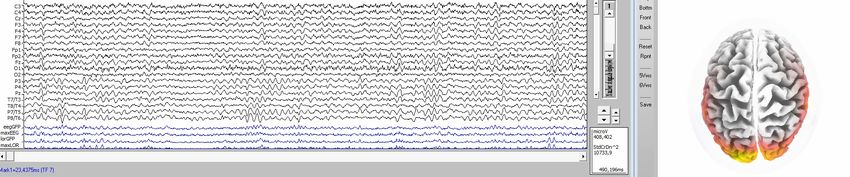
gemessenen Oberflächenpotentialen auf die Quellen zurückrechnen, Bild 3.1/7.
Bild 3.1/7: EEG-Aufnahme nach dem 10-20 System (Koordinatensystem der EEG-
Messelektroden-Punkte auf der Schädeldecke), LORETA-Quellenanalyse
Dr. Garn, Ergänzungen Dr. Ehrlich-Schupita Elektromagnetische Felder und der Mensch Seite 3.1.7Sie können auch lesen

















































