8Folgen des Klimawandels für die - Biodiversität in Wald und Forst - Callistus
←
→
Transkription von Seiteninhalten
Wenn Ihr Browser die Seite nicht korrekt rendert, bitte, lesen Sie den Inhalt der Seite unten

Biodiversität und Klimawandel
8
Folgen des Klimawandels für die
Biodiversität in Wald und Forst
Th. Hickler, A. Bolte, B. Hartard, C. Beierkuhnlein, M. Blaschke, Th. Blick, W. Brüggemann, W. H. O.
Dorow, M.-A. Fritze, Th. Gregor, P. Ibisch, Ch. Kölling, I. Kühn, M. Musche, S. Pompe, R. Petercord,
O. Schweiger, W. Seidling, S. Trautmann, Th. Waldenspuhl, H. Walentowski, N. Wellbrock
8.1 Einführung zialisten gelten (Walentowski et al. 2010), mit er-
heblich mehr Arten (rund 5000 Arten auf Flächen
Deutschland ist ein Waldland. Außerhalb der Al- von unter 100 ha) gerechnet werden, als man bisher
pen stellen Wälder fast überall die potenzielle na- annahm (1500 – 1800 Arten, Weidemann & Schau-
türliche Vegetation (pnV) dar (Ellenberg & Leu- ermann 1986). Bei entsprechendem Strukturreich-
schner 2010, Bohn et al. 2003). Nicht-Waldökosys- tum können solche Wälder eine erhebliche Vielfalt
teme sind natürlicherweise entweder auf azonale beherbergen (deutlich über 6000 Arten, Dorow &
(nicht vom Großklima bestimmte) Sonderstandor- Blick 2010, Meyer et al. 2011).
te beschränkt (z. B. Hochmoore, Flussauen, Küsten- Unsere mitteleuropäischen Wälder befinden
marschen, Felsen und Blockhalden) oder sie sind sich im ständigen Wandel. Ihre heutigen Eigen-
die Folge menschlicher Landnutzung. Nutzungs- schaften können nur vor ihrer historischen Ent-
bedingt bedecken Wälder heute ca. 30 % der Ge- wicklung verstanden werden. Bezüglich der na-
samtfläche der Bundesrepublik, dies sind mehr als türlichen Artenausstattung sind Zeitskalen von
11 Mio. Hektar Fläche (Schmitz et al. 2005). Es gibt mehreren hunderttausend Jahren zu beachten. Im
allerdings auch Regionen, in denen der Mensch das Pleistozän war der überwiegende Teil des heutigen
Wachstum von Wäldern erst möglich gemacht hat, Deutschlands, während der vorherrschenden Eis-
vor allem durch das Trockenlegen von Mooren in zeiten (Glaziale), von periglazialer Steppen-Tund-
Norddeutschland und im Alpenvorland (Ellenberg ren-Vegetation bedeckt (Lang 1994). Die lange an-
& Leuschner 2010). dauernden Glazialperioden wurden immer wieder
Wälder stellen komplexe terrestrische Ökosys- von relativ kurzen Warmzeiten (Interglaziale) un-
teme dar. In keinem anderen terrestrischen Öko- terbrochen. Wir befinden uns derzeit in einer sol-
systemtyp gibt es eine vergleichbare mikroklima- chen Warmzeit. Die Spuren der letzten Eiszeit sind
tische Selbstregulation. Die intensiven biotischen aber allgegenwärtig. Mit jeder warmen Interglazial-
Interaktionen erklären auch die Bereitstellung viel- periode kamen weniger Baumarten aus ihren süd-
fältiger biotisch vermittelter ökologischer Nischen. lichen Rückzugsräumen nach Mitteleuropa zurück,
Global gesehen entfällt deshalb ein großer Teil der so dass dessen Baumartenflora mit jeder Eiszeit an
biologischen Vielfalt auf Wälder. Selbst in den ver- Vielfalt verlor (Beierkuhnlein 2007). Es ist darüber
gleichsweise strukturarmen Waldökosystemen un- hinaus anzunehmen, dass viele Baumarten auch
serer Breiten, in denen wenige Baumarten domi- nach 11 000 Jahren mit mildem und warmem Kli-
nieren, werden hohe Artenzahlen erreicht (z. B. ma noch nicht ihr klimatisch mögliches Ausbrei-
938 Käferarten in einem Buchenwald mit Ahorn- tungsgebiet ausgeschöpft haben (Svenning & Skov
Linden-Schluchtwald-Anteil im Vogelsberg (Flecht- 2004).
ner 2000), bzw. 930 Käferarten in Eichenwäldern Seit der Ausbreitung des Ackerbaus in Deutsch-
des nordostdeutschen Tieflandes (MIL und MLUV land vor ca. 7000 Jahren beeinflusst der Mensch zu-
2010)). Wie die umfangreichen Untersuchungen nehmend die Wälder (Roberts 1998, Küster 2010).
in hessischen Naturwaldreservaten zeigen (s. z. B. Schon in der Steinzeit wurden lokal erhebliche Flä-
Dorow & Blick 2010), kann sogar in durchschnitt- chen entwaldet, weil die aus heutiger Sicht extrem
lichen Buchenwäldern, die generell als arm an Spe- ineffiziente damalige Landwirtschaft einen hohen
164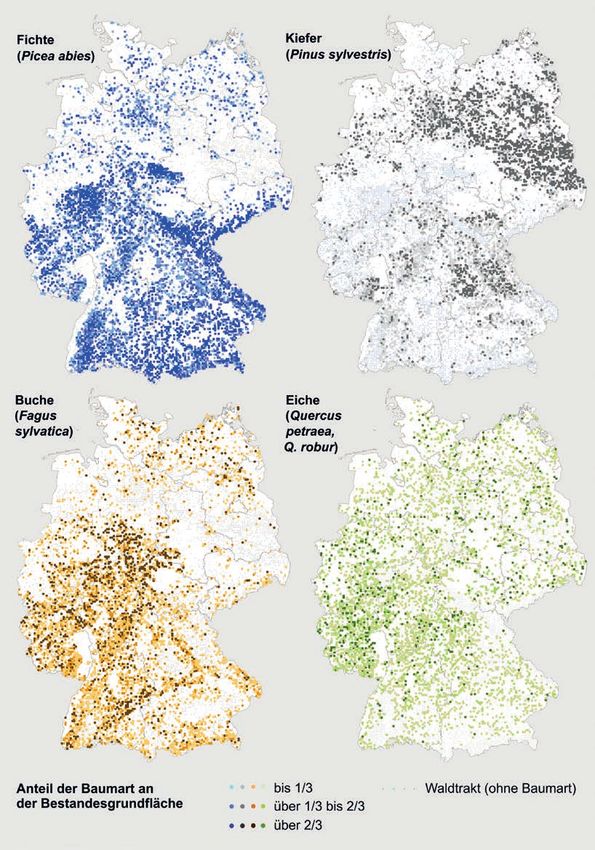
Folgen des Klimawandels für die Biodiversität in Wald und Forst
Abb. 8.1: Der Frankfurter Stadtwald bei Schwanheim gegen Ende des 19. Jahrhunderts. In vielen Gegenden
Deutschlands wurden Wälder vor der industriellen Revolution viel stärker genutzt als heute. Die Wälder waren
offen, nährstoffarm und artenreich. Fritz Wucherer: Bei den Schwanheimer Eichen 1988, Museumsgesellschaft
Kronberg 1986.
Flächenbedarf verursachte (Küster 2010). In der ginn des 19. Jahrhunderts (Backhaus et al. 2000, s.
Bronzezeit stieg der Bedarf nach Holz zusätzlich auch Abb. 8.1). Wahrscheinlich war der überwie-
durch die Erzgewinnung und Verhüttung und seit gende Teil Deutschlands seit dem 14. Jahrhundert
Beginn der Eisenzeit 800 v. Chr. durch weitere In- entwaldet (Bork et al. 1998, Kaplan et al. 2009) und
dustrien, z. B. die Salzproduktion (Backhaus et al. besaß auf weiten Flächen einen parkähnlichen Cha-
2000, Küster 2010). Die Herstellung von 1 kg Glas rakter (Ellenberg & Leuschner 2010). In dieser vom
in Glashütten im Taunus zum Beispiel erforderte Menschen geprägten Kulturlandschaft mit ihrer dif-
im 17. Jahrhundert ungefähr 1 m3 Holz (Weisbe- ferenzierten Flächennutzung (Streu und Harznut-
cker 1988). Waldweide und die Nutzung von Laub- zung, Plaggenwirtschaft, Brandwirtschaft, vielfältige
und Nadelstreu als Strohersatz im Stall und für die Formen der Nieder- und Mittelwaldnutzungen, Hu-
Bodenverbesserung auf Äckern entzogen den Wäl- tewälder u. a.) hat eine Ausweitung des vorhande-
dern zusätzlich Nährstoffe. Die starke Nutzung der nen Spektrums der Standorte und Ökosystemtypen
Wälder vor der industriellen Revolution führte stattgefunden. Einige dieser historischen Waldnut-
schließlich dazu, dass unsere Wälder zu Beginn des zungsformen sind gelegentlich noch anzutreffen,
19. Jahrhunderts einen historischen Tiefstand be- zum Beispiel im Alpenvorland oder im fränkischen
züglich ihrer flächigen Erstreckung aber auch be- Grabfeld, und haben erhebliche Bedeutung für den
züglich ihrer qualitativen Eigenschaften erreichten. Artenschutz. Waldökosysteme mit für Hochwald
Geschlossene Wälder waren kaum mehr vorhan- typischen Ökosystemeigenschaften waren bis zum
den. Die Hessische Landesforstverwaltung schätzt, Beginn des 19. Jahrhunderts in dicht besiedelten
dass die Wälder in vielen hessischen Mittelgebirgen Gegenden selten. Auch die natürlichen Wälder vor
heute zehnmal mehr Biomasse enthalten als zu Be- dem Eingriff des Menschen waren aufgrund vielfäl-
165
Biodiversität und Klimawandel
Abb. 8.2: Dürreschäden an Eichen und Buchen am Rand des Fichtelgebirges bei Bad Berneck im August 2003.
Foto Jurasinski/Beierkuhnlein.
tiger Störungen, wie z. B. Brände und Überschwem- Waldes in den vergangenen Jahrhunderten massiv
mungen, unter Umständen offener als viele unserer verändert hat.
Nutzwälder heute (Bradshaw et al. 2003). In Zukunft könnte auch der Klimawandel unse-
Eine ungefähre Vorstellung der jüngeren Ver- re Wälder und deren Biodiversität stark verändern.
änderung der Biotopvielfalt im Wald lässt sich auch Auswirkungen klimatischer Extreme auf Bestandes-
aus einem Zahlenvergleich der Betriebsarten ablei- ebene waren im vergangenen Jahrzehnt mehrfach
ten. So bestand der Wald 1883 (einem Zeitpunkt großflächig in Mitteleuropa zu beobachten. Im Jahr
also, als bereits massiv mit der Überführung von 2003 war es eine in dieser Weise ungekannte Dürre-
Nieder- und Mittelwäldern in Hochwälder begon- und Hitzeperiode, welche nicht nur einzelne Arten
nen worden war) in Deutschland (alte Bundes- betraf, sondern Waldbestände flächig zu verfrühtem
länder) aus 82 % Hochwald, 7,8 % Mittelwald und Blattwurf, verbunden mit Wachstumseinbußen führ-
10,2 % Niederwald (Mantel 1990). Nach den Ergeb- te (Spiecker et al. 2005, Fischer et al. 2009, Abb. 2).
nissen der ersten Bundeswaldinventur 1987 bestand Klimatische Wetterextreme und ihre Folgen
der Wald in diesen Landesteilen aus 98,4 % Hoch- werden wahrscheinlich zu weiteren wirtschaftli-
wald, 0,5 % Mittelwald und 1,1 % Niederwald (Bun- chen Einbußen führen bzw. die Bestandsstruktur
desministerium für Ernährung, Landwirtschaft und maßgeblich ändern (Kap. 2.1 und z. B. Christensen
Forsten 1990). Schmidt (1989) belegt in seiner Ar- & Christensen 2004, Schäret al. 2004, Peñuelas et al.
beit im Schwarzwald für das Jahr 1780 ein Verhält- 2007). Wälder und Gebüschgesellschaften gehören
nis von Wald zu Reut- und Weidfeldern (durch Ro- in Deutschland zu den Pflanzengemeinschaften, die
dung für Ackerbau und Weide nutzbar gemacht) einen großen Anteil von durch den Klimawandel
von 52 % zu 48 %; bereits 1925 waren diese Reut- betroffenen Pflanzenarten aufweisen (Pompe et al.
und Weidfelder bis auf einen vernachlässigbaren 2010). Es stellt sich aber die Frage, welche Wald-
Rest verschwunden. Diese historische Betrachtung ökosysteme angesichts der kontinuierlichen Klima-
macht deutlich, dass sich auch die Biodiversität des veränderung und der voraussichtlich steigenden
166
Folgen des Klimawandels für die Biodiversität in Wald und Forst
Anzahl Extremereignisse stärker und welche weni- 8.3 Arealmodellierung
ger stark gefährdet sind und wie sich die Lebensge-
meinschaften in unseren Wäldern verändern wer- Seit Mitte der Neunziger Jahre werden Arealmodel-
den (Bittner & Beierkuhnlein im Druck). le vermehrt genutzt, um mögliche klimabedingte
zukünftige Veränderungen der Verbreitungsgebie-
te von Pflanzen- und Tierarten abzuschätzen (z. B.
8.2 Aufbau des Kapitels Sykes et al. 1996, Thuiller et al. 2005, Kölling et al.
2007, Huntley et al. 2007, Settele et al. 2008). Die
Zunächst beschreiben wir anhand eines Beispiels Modelle basieren in der Regel auf Korrelationen
aus der Forstwirtschaft ein wichtiges Werkzeug zur zwischen Klimavariablen und Verbreitungskarten.
Abschätzung möglicher Folgen des Klimawandels In einigen wenigen Fällen wurden auch nicht-kli-
für die Biodiversität, sogenannte „Arealmodelle“ matische Umweltfaktoren berücksichtigt, insbe-
(Kap. 2) und diskutieren deren Anwendbarkeit. Die- sondere Landnutzung und Böden (z. B. Pompe et
se Modelle werden benutzt, um zu untersuchen, wie al. 2008). Weil die Modelle für Forscher, die nicht
vor allem klimatische Umweltfaktoren die Verbrei- mit Modellen arbeiten oder interessierte Laien oft
tung einer Art bestimmen. In Kombination mit Kli- schwer verständlich sind, illustrieren wir sie hier an
maszenarien können solche Modelle genutzt wer- einem Beispiel, einer Studie über die Weiß-Tanne
den, um mögliche Veränderungen des Areals in die (Abies alba).
Zukunft zu projizieren.
Dann wird diskutiert, wie der Klimawandel Wald- 8.3.1 Modellierung des heutigen und
ökosysteme auf der Ebene von Bäumen, welche die potenziell zukünftigen Areals der
Habitate für eine Vielzahl von Organismen maß- Weiß-Tanne
geblich formen, voraussichtlich beeinflussen wird.
Hierbei unterscheiden wir die Auswirkungen des Wie viele andere Pflanzenarten, kommen Baumar-
Klimawandels für die Verbreitungs- und Wuchsbe- ten stets innerhalb begrenzter geografischer Räume
dingungen der wichtigsten Baumarten (Abs. 8.3), In- vor. Klima, geografische Hindernisse (wie Höhen-
teraktionen mit Luftschadstoffen (z. B. Stickstoffde- züge), aber auch Landnutzung und biologisch-öko-
position, Abs. 8.4.1) und wie der Klimawandel Stö- logische Faktoren (z. B. Konkurrenz) limitieren die
rungen, z. B. durch Schädlinge (Insektenkalamitäten, Verbreitungsgebiete (Woodward 1987). Am Bei-
Pilzbefall und Baumkrankheiten) und Sturmschä- spiel einer mitteleuropäischen Baumart, der Weiß-
den, beeinflussen könnte (Abs. 3.2). Mögliche Ent- Tanne, die vor allem im Gebirge vorkommt (Abb.
wicklungen bei der Nutzung von nicht heimischen 8.3), lässt sich dies demonstrieren. Einzelne Rand-
Baumarten werden gesondert behandelt (Abs. 8.3.3), vorkommen gibt es an der französischen Küste und
ebenso eine Fallstudie über die mögliche Eignung in Dänemark, die auf forstlichen Anbau zurückge-
von mediterranen Eichen (Kasten I). Schließlich folgt hen. Um solche geografischen Muster zu erklären,
eine Zusammenfassung möglicher waldbaulicher können u. a. klimatische Parameter herangezogen
Anpassungsmaßnahmen, welche für die Biodiver- werden. Einfache Zusammenhänge lassen sich be-
sität in unseren Wäldern relevant sind (Abs. 8.3.4). reits zwischen der Jahresmitteltemperatur (Hijmans
Im zweiten Teil werden die Folgen des Klima- et al. 2005) und den Vorkommen bzw. Nichtvor-
wandels für verschiedene Organismengruppen kommen der Art finden (Abb. 8.3, 8.4).
(Vögel, Blüten- und Farnpflanzen, Pilze und aus- In bioklimatischen Arealmodellen werden die
gewählte Arthropodengruppen) von Experten auf- Beziehungen zwischen Klimavariablen und den
grund von relevanten Merkmalen und, soweit vor- Vorkommen bzw. Nicht-Vorkommen von Arten
handen, Modellierungsergebnissen, sowie der Bin- oder Populationen statistisch erfasst. Damit wird
dung an bestimmte Waldtypen abgeschätzt ( Abs. es für jeden geografischen Punkt (sofern es die Da-
8.4). In Kasten II werden die Ergebnisse von Unter- tenlage zulässt) möglich, zu ermitteln, wie wahr-
suchungen entlang eines Höhengradienten im Bay- scheinlich es ist, dass eine Art dort potenziell vor-
erischen Wald präsentiert, Schlussfolgerungen und kommt. Diese errechnete sog. Vorkommenswahr-
Handlungsempfehlungen folgen in Abs. 8.5. scheinlichkeit reicht von 0 für den Fall, dass ein
167
Biodiversität und Klimawandel
Abb. 8.3: Geographische Darstellung der Vorkommen („Präsenzen“, blaue Punkte) und Nicht-Vorkommen („Ab-
senzen“, graue Punkte) der Weiß-Tanne. Datei aus: Europäisches Waldmonitoring-Netz Level I.
bination von Jahresdurchschnittstemperatur und
Jahresniederschlagssumme andererseits in einem
Generalisierten Additiven Modell (GAM, Hastie
& Tibshirani 1990) nachbildet. Isolinien gleicher
Auftretens-Wahrscheinlichkeit verdeutlichen nach
der Art von Höhenlinien den Aufbau des resultie-
renden „Wahrscheinlichkeitsgebirges“. Seinen Gip-
fel hat dieses bei 5 bis 7 °C und 800 bis 1300 mm
Jahresniederschlag. Bei einer Wahrscheinlichkeit
von 0,1 (10 %) an der äußersten Isolinie sind die
Bedingungen für das Gedeihen der Weiß-Tanne
so ungünstig geworden, dass man die Art, wenn
überhaupt, nur noch äußerst selten antreffen wird.
Dieser geringe Wahrscheinlichkeitswert markiert
gleichzeitig den absoluten Anbauschwellenwert.
Abb. 8.4: Modellierte Vorkommenswahrscheinlichkeit Jenseits dieses Werts steigt das Anbaurisiko so
der Weiß-Tanne. stark an, dass man unter diesen Bedingungen auf
einen Anbau verzichten sollte (Kölling et al. 2011).
Vorkommen der Art auszuschließen ist, bis ma- Über diesen Weg gelangt man somit zu einer Aus-
ximal 1, wenn die Art dort mit sehr hoher Wahr- sage über potenzielle Vorkommen einer Art, zum
scheinlichkeit durch das Modell vorhergesagt wird. Beispiel auf der Grundlage von Klimavariablen. Es
In Abb. 8.4 ist ein sehr einfaches Arealmodell darge- gibt aber eine Vielzahl von Verfahren, mit denen
stellt, das den Zusammenhang zwischen dem Vor- man Techniken verfeinern und auch Modelle prü-
kommen der Weiß-Tanne einerseits und der Kom- fen kann.
168
Folgen des Klimawandels für die Biodiversität in Wald und Forst
8.3.1.1 Variablenauswahl Resultat der Anwendung von Arealmodellen
Die Variablenauswahl spielt bei der Arealmodellie- sind die vorläufigen Klima-Risikokarten, die in der
rung eine große Rolle. Je nach Ansatz und Zielstel- Bayerischen Forstverwaltung seit dem Jahr 2010
lung einer Studie werden dabei mehrere oder weni- im Einsatz sind (Kölling et al. 2009a und b, Köl-
ge Klimavariablen bevorzugt. In unserem Beispiel ling et al. 2010). Sie basieren auf einer Regionali-
mit der Weiß-Tanne ist es gelungen, bereits mit den sierung einfacher Arealmodelle für die wichtigsten
zwei Variablen Jahresniederschlagssumme und Jah- Baumarten, nämlich Gemeine Fichte (Picea abies),
resdurchschnittstemperatur eine beachtliche Erklä- Wald-Kiefer (Pinus sylvestris), Europäische Lärche
rung der Präsenzen bzw. Absenzen und eine reali- (Larix decidua), Weiß-Tanne, Douglasie (Pseu-
tätsnahe Simulation der heutigen Tannenvorkom- dotsuga menziesii), Buche (Fagus sylvatica), Stiel-
men zu erzielen. Es ist jedoch nicht klar, über welche Eiche (Quercus robur), Trauben-Eiche (Quercus pe-
physiologischen Prozesse diese beiden Faktoren die traea) und Berg-Ahorn (Acer pseudoplatanus) für
Verbreitung der Weiß-Tanne bestimmen. Darum ha- die gesamte Waldfläche Bayerns.
ben einige Autoren nur Klimavariablen ausgewählt, Die Karte für die Weiß-Tanne ist in Abb. 8.5
für die eine eindeutige Hypothese darüber besteht, dargestellt (Kölling et al. 2011). Für die drei Zeit-
durch welchen physiologischen Prozess die jeweilige punkte 2000, 2050 und 2100 enthalten die Karten
Variable für die Verbreitung einer Art maßgebend ist eine Schätzung des Anbaurisikos der Weiß-Tanne,
(Woodward & McKie 1991, Sykes et al. 1996, Sven- welches aus der im europäischen Maßstab ermit-
ning & Skov 2004). Es ist z. B. bekannt, dass Bäu- telten Abhängigkeit der Vorkommenswahrschein-
me bestimmte physiologische Anpassungen besit- lichkeit von Jahresniederschlagssumme und Jahres-
zen, um verschiedene Niveaus von Extremkälte im durchschnittstemperatur abgeleitet ist (Kölling et al.
Winter zu überleben (Woodward 1987). Deswegen 2011). Die Karten verdeutlichen die Abnahme der
wird in einigen Modellen die minimale monatliche für den Anbau optimalen Lagen mit sehr geringem
Wintertemperatur als Variable benutzt (z. B. Sykes Risiko (blaue Farbe), die sich nach und nach immer
et al. 1996). Bei der Verwendung von Mittelwerten mehr auf die höheren Gebirge konzentrieren, wäh-
als Eingangsparameter für die Arealmodellierung ist rend sich die wärmsten und trockensten Lagen im
zudem kritisch anzumerken, dass die aktuelle Bezie- Flach- und Hügelland zusehends weniger für den
hung zwischen limitierenden, extremen Witterungs- Anbau eignen (rote Farbe).
bedingungen (z. B. Hitze- und Trockenperioden) Die Klima-Risikokarten werden in Bayern als
und den verwendeten Temperatur- und Nieder- Hilfsmittel im staatlichen Forstbetrieb und bei
schlagsmitteln einfach in die Zukunft projiziert wird. der Beratung der privaten und körperschaftlichen
Davon kann aber nach heutigem Kenntnisstand eher Waldbesitzer eingesetzt. Sie können dazu beitragen,
nicht ausgegangen werden (Bolte et al. 2008). einen klimagerechten Waldumbau, bei dem anfälli-
ge durch resistentere Baumarten ersetzt werden, vo-
8.3.1.2 Regionalisierung und Projektionen ranzubringen. Letztlich geht es darum, bei der Pla-
im Klimawandel nung des Waldes von morgen die Überschreitung
Das in Abb. 8.4 vorgestellte Arealmodell kann auch von kritischen Anbauschwellenwerten möglichst zu
in der Fläche dargestellt und auf zukünftige Klima- vermeiden und optimal an die heutigen und künf-
verhältnisse angewandt werden (Kühn et al. 2009). tigen Klimabedingungen gleichermaßen angepasste
Dazu können gerasterte Klimakarten verwendet Wälder zu begründen.
werden, die entsprechende Einflussgrößen – in un- In der Forstwirtschaft haben Arealmodelle ge-
serem Beispiel die Jahresdurchschnittstemperatur rade erst Einzug gehalten. In anderen Disziplinen,
und die Jahresniederschlagssumme – nicht für die vor allem der Ökologie, wurden mit ihnen mittler-
Gegenwart, sondern auch für eine durch Klima- weile die Folgen des Klimawandels für die Verbrei-
wandel geprägte Zukunft enthalten. Je nachdem, tung von Hunderten von Pflanzen- und Tierarten
welches Szenario für Treibhausgasemissionen, wel- abgeschätzt. Die Prinzipien, nach denen die Model-
ches globale Klimamodell und welches regiona- le konstruiert werden, sind in der Regel ähnlich wie
le Klimamodell benutzt werden, resultieren unter in unserem Beispiel mit der Weiß-Tanne. Es steht
Umständen abweichende Ergebnisse. jedoch eine Vielzahl von statistischen Verfahren zur
169
Biodiversität und Klimawandel
Abb. 8.5: Klima-Risikokarten für die Weiß-Tanne 2000, 2050 und 2100 in Bayern. Kölling et al. 2011.
Verfügung (Guisan & Zimmermann 2000, Pearson (Abb. 8.6) errechnet wird. Unterschiedliche Fähig-
& Dawson 2003, Kühn et al. 2009, Pompe et al. keiten der Arten, Entfernungen zu klimatisch neu
2008 für Deutschland). Die Spannbreite reicht von passenden Gebieten zu überwinden, können zwar
relativ einfachen multiplen Regressionsmodellen auch berücksichtigt werden, doch liegen bei der
bis zu neuronalen Netzwerken und bayesianischen Mehrzahl der Arten nur unzureichende Informati-
Ansätzen (Guisan & Zimmermann 2000, Franklin onen über das Ausbreitungsvermögen vor, weshalb
2010, Pagel & Schurr 2012).
Verschiedene Analysevariablen können für eine
Risikoanalyse benutzt werden, wobei der räumli-
che Verlust (Arealverlust), Gewinn (Arealgewinn)
oder Arealwandel für Arten unter Berücksichtigung
von Gewinn und Verlust an potenziellen Gebieten
Abb. 8.6: Schema eines potenziellen Verbreitungsge-
bietes und dessen Projektion für eine Art durch ein
Arealmodell für einen definierten geographischen
Raum (Rasterzellen), Unterscheidung Arealverlust:
2 Rasterzellen, Arealgewinn 2 Zellen, Arealverlust =
Fläche der Rasterzellen des jetzigen Areals, welche
in Zukunft nicht mehr den Umweltansprüchen der Art
entsprechen, Arealgewinn = Fläche der Rasterzellen,
die zurzeit, nicht aber in der Zukunft, den Umwelt-
ansprüchen der Art entsprechen (welche jedoch oft
nicht schnell genug erreicht und kolonisiert werden
können), Arealwandel = Arealgewinn – Arealverlust.
Modifiziert nach Midgley et al. 2006. Vgl. Pompe et al. 2011.
170
Folgen des Klimawandels für die Biodiversität in Wald und Forst
i. d. R. für Berechnungen alle potenziell geeigneten Regel mobiler sind, scheinen weitestgehend ihr kli-
Räume einbezogen werden. matisches Potenzial erreicht zu haben (Hawkins
und Porter 2003).
8.3.2 Kritik an Arealmodellen Von großer Bedeutung könnten biotische Inter-
aktionen mit Pathogenen sein, deren Ausbreitung
Je nach verwendetem Verfahren weichen artspezi- durch den Klimawandel befördert wird ( s. Abs.
fische Aussagen voneinander mehr oder minder ab 8.3.2). In den vergangenen Jahrzehnten kam es in
(Pompe et al. 2010), was bei Interpretationen von Deutschland mehrfach zu einem sich expansiv aus-
Modellen beachtet werden muss. Dennoch kann breitenden Auftreten von Krankheitserregern von
für eine Vielzahl von Arten gleichzeitig eine erste Bäumen, z. B. des Erregers des Eschentriebsterbens
grobe Risikoabschätzung unter den Bedingungen (Schumacher et al. 2007). Das Ulmensterben hat
des Klimawandels durchgeführt werden. In diesem gezeigt, dass solche Ereignisse zum lokalen Erlö-
Zuge werden jedoch einige biologisch und evoluti- schen von Populationen einzelner Baumarten füh-
onär relevante Punkte, wie biologische Interaktio- ren können (Butin 2011). Das (vermehrte) Auftre-
nen, Adaptation, genetische und ökologische Un- ten eines pathogenen Organismus ist oftmals nicht
terschiede zwischen regionalen Populationen und oder nicht zweifelsfrei auf Klimawandelwirkungen
Konkurrenz vernachlässigt (u. a. Bolte et al. 2008). zurückzuführen, verdeutlicht aber die Grenzen der
Dies führt zu Unsicherheiten in den Modellaussa- Modellierbarkeit. Dies kommt bei sogenannten
gen (Kühn et al. 2009, Garzón et al. 2011). Die mo- Komplexkrankheiten mit Synergiewirkungen ver-
dellierten Arealveränderungen dürfen deshalb kei- schiedener Stressoren (siehe unten) noch stärker
nesfalls als konkrete Prognosen gedeutet werden. zum Tragen.
Sie stellen lediglich dar, wie sich ein hypothetisches
Fenster von Umweltvariablen, welches mit der heu-
tigen Artverbreitung korreliert, in der Zukunft in 8.4 Folgen des Klimawandels
einem bestimmten Klimaszenario verschiebt. Weil für die Verbreitungsgebiete
die Korrelationen auf der heutigen Verbreitung ba- und Wuchsbedingungen der
sieren, spiegeln sie nur die unter Interaktionen mit
anderen Arten, vor allem Konkurrenz, realisier-
wichtigsten Baumarten
te Nische wider, nicht die physiologisch mögliche Zu den heimischen Baumarten mit der größten
fundamentale Nische (sensu Hutchinson 1957). aktuellen Verbreitung in Deutschland gehören die
Gerade im Zuge des raschen Klimawandels können Gemeine Fichte und die Wald-Kiefer als Nadel-
sich jedoch biotische Interaktionen verändern, z. B. baumarten sowie die Buche und die Trauben- und
dadurch, dass ein wichtiger Konkurrent oder eine Stiel-Eiche als Laubbaumarten. Die häufigste nicht
Nahrungs- bzw. Beuteart bzw. Bestäuber wegfällt, heimische Baumart ist die Douglasie (s. Tab. 8.1).
weil er sich nicht so schnell ausbreiten kann wie die Die deutschen Verbreitungsschwerpunkte dieser
Zielart (Schweiger et al. 2008, 2010, 2012). Außer- fünf Baumarten unterscheiden sich z. T. erheblich
dem ist unklar, inwiefern die heutigen Verbreitun- (Abb. 8.7). Während Fichte und Buche vorwiegend
gen von Arten aktuelle Umweltverhältnisse oder in den Mittelgebirgslagen Deutschlands verbreitet
historische menschliche oder klimatische Einflüsse sind, liegt der Schwerpunkt des Wald-Kiefern-Vor-
widerspiegeln (z. B. Giesecke et al. 2007). Wie be- kommens im norddeutschen Tiefland neben regio-
reits erwähnt, haben Baumarten eventuell ihr kli- nalen Verbreitungen in Franken, der Oberpfalz und
matisches Potenzial noch gar nicht erreicht (Sven- dem Pfälzer Wald. Die heimischen Eichenarten sind
ning & Skov 2004). In Bezug auf unser Beispiel mit gleichmäßiger verbreitet mit regionalen Schwer-
der Weiß-Tanne ist jedoch zu erwähnen, dass die punkten in süd- und westdeutschen Mittelgebirgen.
Verteilung der Hauptbaumarten in Deutschland Auch die Douglasie kommt häufiger in den Mittel-
heute wahrscheinlich nicht mehr durch die Migra- gebirgen Südwestdeutschlands vor (Baden-Würt-
tion seit der letzten Eiszeit limitiert ist. Außerdem temberg, Hessen, Rheinland-Pfalz), wurde und wird
unterscheiden sich verschiedene Organismengrup- aber auch auf einigen Standorten im Norddeutschen
pen in ihrer Mobilität. Schmetterlinge, die in der Tiefland angebaut (Schmitz et al. 2005).
171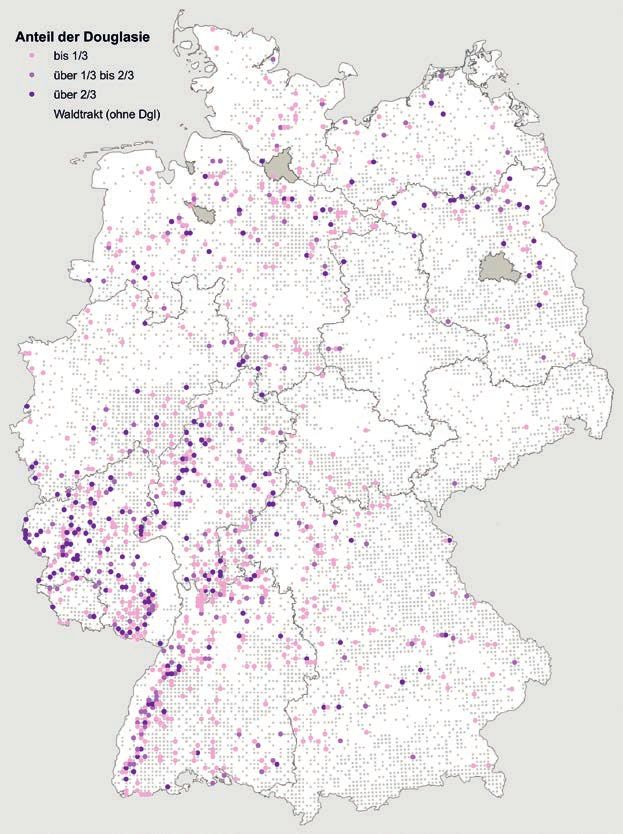
Biodiversität und Klimawandel
Tab. 8.1: Waldfläche (nur Holzboden) und Waldflächenanteil der wichtigsten Baumarten in Deutschland (Stich-
jahr 2008) sowie Veränderung der Waldfläche in der Periode von 2002 bis 2008. Datengrundlage: Inventurstudie
Bundeswaldinventur*, Oehmichen et al. 2011
Baumart Waldfläche* [Mio. ha] Waldflächenanteil [%] Veränderung 2002 – 2008 [ha]
Fichte 2,68 26,0 – 210 555
Wald-Kiefer 2,41 23,4 – 52 082
Buche 1,68 16,3 + 83 146
Eichen 0,95 9,2 + 42 494
Douglasie 0,22 2,1 + 27 357
Andere Baumarten 2,13 20,6 + 100 392
Blößen, Lücken 0,24 2,4 + 9 248
Gesamt 10,31 100,0
* nach Inventurstudie 2008; Abgänge der Waldfläche zwischen 2002 (Stichjahr 2. Bundeswaldinventur: 10,57 Mio. ha Holzbodenflä-
che) und 2008 wurden berücksichtigt, Erstaufforstungen allerdings nicht (Oehmichen et al. 2011).
Im Fall der Eichen und der Buche liegt können deutlich höher sein (vgl. Bolte et al. 2007).
Deutschland mehr oder weniger im Zentrum des Im Allgemeinen gilt aber, dass die Buche und ins-
europäischen Verbreitungsgebietes, während die besondere die Fichte höhere Niederschläge wäh-
Fichte in den Tieflagen an die Wärmegrenze ihres rend des Jahres und der Vegetationsperiode (Mai
Verbreitungsgebietes stößt (Abb. 8.8). Weiter süd- bis September) sowie kühlere Sommer benötigen
lich kommt die Fichte natürlich vor allem mon- als die Wald-Kiefer und die beiden Eichenarten.
tan und subalpin vor. In gewissen Grenzen können Günstige Klimabedingungen für Buche und Fichte
Bäume jedoch auch außerhalb ihres natürlichen liegen in den deutschen Mittelgebirgslagen und für
Verbreitungsgebietes angebaut werden (Abb. 8.8), Buche im küstennahen Tiefland vor, so dass diese
u. a. weil man bei Pflanzungen Limitierungen wäh- Baumarten in diesen Räumen produktiv und kon-
rend der Keimlingszeit (Dahl 1998) umgeht. Auch kurrenzstark aufwachsen können. Bei der Buche
die Wald-Kiefer ist eine nordöstlich verbreitete Art, treten allerdings deutliche Unterschiede zwischen
sie ist jedoch toleranter als die Fichte gegenüber ho- verschiedenen Herkünften bezüglich der Sensibili-
hen Temperaturen und Trockenheitsstress, so dass tät gegenüber Dürre und Spätfrost auf, so dass man
sie weiter ins Tiefland und nach Süden vordringt. nicht einfach südliche Herkünfte einsetzen könnte
Szenarien für das Klima am Ende des Jahrhunderts (Kreyling et al. 2011, 2012).
entsprechen einer Verschiebung der Klimazonen Für die in Mitteleuropa im Vergleich mit der
um mehrere hundert Kilometer nach Norden bzw. Buche generell konkurrenzschwächere Wald-Kiefer
Nordosten (Ohlemüller et al 2006). Viele Gebiete, und die beiden Eichenarten (Ellenberg & Leuschner
in denen heute Fichtenwald steht, würden dann 2010) sind günstige Wuchsbedingungen dagegen im
Klimabedingungen aufweisen, unter welchen die kontinental getönten Klima des zentralen östlichen
Fichte zurzeit gar nicht oder nur schlecht gedeiht Norddeutschen Tieflands zu finden, wo die Wuchs-
(z. B. Sykes et al. 1996). bedingungen für Buche (und auch Fichte) aufgrund
Die unterschiedlichen Verbreitungsschwer-
punkte der Baumarten spiegeln ihre klimatischen Abb. 8.7: Beteiligung der Hauptbaumarten Fichte,
Minimumansprüche wider (Tab. 8.2). Allerdings Wald-Kiefer, Buche und Eichen (Trauben- und Stiel-
treffen die genannten Ansprüche meist nur auf Eiche) in Deutschland an der Bestandesgrundfläche
die angepassten Lokalherkünfte an den Verbrei- (Stichjahr 2002, Bearbeitung Franz Kroiher, vTI) nach
tungsrändern zu, die unter den entsprechenden den Aufnahmen der zweiten Bundeswaldinventur.
Extrembedingungen wachsen. Die Ansprüche von (BWI, Schmitz et al. 2005); Angaben zum Vorkommen der
Herkünften aus dem zentralen Verbreitungsgebiet Douglasie finden sich in Abb. 8.10.
172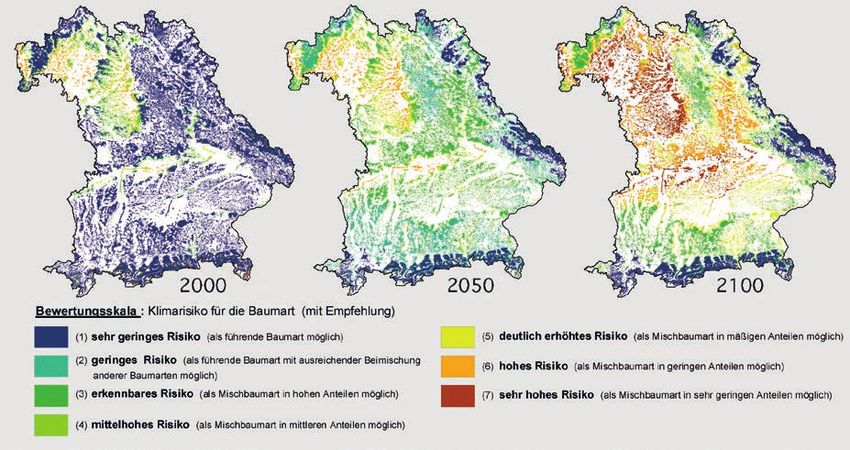
Folgen des Klimawandels für die Biodiversität in Wald und Forst
173Biodiversität und Klimawandel
Abb. 8.8: Natürliche europäische Verbreitungsgebiete der Stiel-Eiche (a.) und der Fichte (b.). Tutin et al. 1964 – 1980.
der Niederschlagsarmut, der Sommerwärme sowie Douglasie weichen von diesem Muster jedoch ab,
der geringen Wasserspeicherkapazität der dominie- da sie einerseits an der nordamerikanischen Pazi-
renden Sandböden eher ungünstig sind. fikküste und dem Kaskadengebirge bei hohen Win-
Minimumansprüche wie die Wald-Kiefer und terniederschlägen gedeihen. Andererseits sind die
die beiden Eichenarten haben auch die amerika- Vegetationsperioden in Regionen der Kaskaden mit
nischen Inlandsherkünfte der Douglasie, die bei- größerer Entfernung zum Meer von geringen Nie-
spielsweise aus den kontinentalen Klimaten der Ro- derschlägen und Hitze geprägt (Hermann & Laven-
cky Mountains stammen. Die Küstenherkünfte der der 1990). Aus der kontinental-borealen Verbrei-
174Folgen des Klimawandels für die Biodiversität in Wald und Forst
Tab. 8.2: Klimatische Minimumansprüche wichtiger Baumarten sowie zusätzliche Faktoren, die deren Vor-
kommen und Wachstum beeinflussen: Jones (1959), Carlisle & Brown (1968), Sakai & Okada (1971), Sakai
(1974), Larsen & Ruetz (1980), Mayer (1984), Institut für Waldbau (1987), Hermann & Lavender (1990), Lyr et
al. (1992), Walter & Breckle (1991), Deans & Harvey (1996), Višnjić & Dohrenbusch (2004), Czajkowski & Bol-
te (2006), Bolte et al.(2007), Morin et al. (2007), Ellenberg & Leuschner (2010), Kathke & Bruelheide (2011),
USDA (2011); k. A.: keine Angaben.
Baumart Niederschlagsmittel Mitteltemp. Max. Winterfrostresistenz Zusätzliche wichtige
Jahr / Mai – Sept. max. Juli/ Altbestand/Verjüngung Schadfaktoren
[mm] min. Jan. [°C] [°C]
Fichte 600 / 300 18 / –18 –60 / –50 (–107)1 Windwurf, Borkenkäferbefall
Spätfrost,
Buche 500 / 250 19 / –3 –35 / –19 (–21)1 Buchenrindenkrankheit
(Schleimfluss)
Douglasie2 Spätfrost, Frosttrocknis,
600 / 200 30 / –9 –33 / –20 (–25)1
Küstenform Douglasienschütte,
360 / 220 21 / –9 –32 / –30
Inlandsform Hallimasch
Insektenbefall, Komplex-
Wald-Kiefer 400 / 200 27 / –20 –90 / –70
schädigung (Beispiel Wallis)
Eichenarten: Spätfrost,
3003 / 200 25 / –16 k. A. / –561
Stiel-Eiche Eichen-Komplexkrankheit,
490 / k.A. 25 / –4 –30 / –201
Trauben-Eiche Insektenbefall
1
Verjüngung LT 50 (Letale Temperatur, bei der 50 % der Verjüngungspflanzen absterben). 2Amerikanisches Verbreitungsgebiet 3Mit
Grundwasseranschluss.
tung von Fichte (bis Nordskandinavien, Westsibiri- und -bruch gilt die Fichte, deren häufig flach aus-
en) und Wald-Kiefer (gesamtes Sibirien) leitet sich geprägtes Senkerwurzelsystem besonders auf ver-
deren extreme Winter- und analog auch Spätfrost- nässten Mittelgebirgsstandorten höheren Bäumen
härte ab (Walter & Breckle 1991). Die Buche, die wenig Halt bei Stürmen liefert (Köstler et al. 1968,
Trauben-Eiche und auch die Küsten-Douglasie mit Puhe 2003).
ausschließlich temperater Verbreitung leiden dage- Anhand der o. g. klimatischen und standört-
gen stärker unter extremer Winterkälte und Spät- lichen Toleranzgrenzen sowie den begleitenden
frostereignissen. Die Stiel-Eiche, mit kontinentaler Schädigungen, lassen sich grundsätzliche Annah-
Verbreitung bis nach Südrussland und die Inlands- men zu den zukünftigen Wuchs- und Existenzbe-
Douglasie sind hier als intermediär anzusehen. dingungen der Baumarten unter den Bedingungen
Als zusätzliche Gefährdung, die mit Klima- und eines Klimawandels ableiten. Bei einer erwarteten
Witterungseinwirkung in Zusammenhang steht, Erwärmung, dem verstärktem Auftreten von Tro-
kann Frosttrocknis angesehen werden, unter der ckenphasen und einer möglichen Häufung von
besonders die Douglasie leiden kann. Frosttrocknis ex-tremen Sturmereignissen in Mitteleuropa (s.
wird ausgelöst durch Wassermangel aufgrund von Abs. 8.3.2) würden sich auf vielen Standorten die
winterlicher Transpiration an Strahlungstagen bei Wuchs- und Existenzbedingungen besonders der
gefrorenem Boden und dadurch eingeschränkter Fichte verschlechtern. Eine wichtige destabilisieren-
Wassernachlieferung. Im Herkunftsgebiet der Dou- de Rolle kann dabei die Wechselwirkung zwischen
glasie tritt Frosttrocknis wegen der hohen Schnee- Trockenheit, Totholzangebot nach Windwurf und
bedeckung und geringer winterlicher Verdunstung Borkenkäferbefall spielen (Schlyter et al. 2006, Bol-
nicht auf (Larsen & Ruetz 1980, Institut für Wald- te et al. 2010, Abs. 8.3.2).
bau 1987). Nach Trockenjahren tritt bei der Buche Generell sind es Interaktionen zwischen ver-
vermehrt eine Komplexerkrankung mit Kambium- schiedenen Stressfaktoren, welche den Wald am
schäden und Schleimfluss (Buchenrindenkrank- stärksten schädigen. Bei Eichen und Wald-Kiefer
heit) auf (Röhrig et al. 2006, Arbeiter & Blaschke z. B. werden Komplexschädigungen (sog. Eichen-
2009). Als besonders gefährdet durch Windwurf sterben, Thomas et al. 2003, Kiefernsterben im Wal-
175Biodiversität und Klimawandel
lis, Bigler et al. 2006, Rigling et al. 2010) beobach- 2005, Michiels et al. 2009). Insbesondere auf flach-
tet, an denen wahrscheinlich sowohl abiotische als gründigen oder sandigen Böden und im Norddeut-
auch biotische Ursachen beteiligt sind. Die Doug- schen Tiefland könnten sich die Wachstumsbedin-
lasie kann insbesondere von pilzlichen Schaderre- gungen für sie durch erhöhte Temperaturen und
gern wie der Rostigen Douglasienschütte, der Ru- geringere Sommerniederschläge (vgl. Kapitel 2.1)
ßigen Douglasienschütte und von Hallimasch befal- entscheidend verschlechtern. Wald-Kiefer und Ei-
len werden. Insbesondere die Inlandsherkünfte der chen können hier auf zukünftig trockenen Stand-
Douglasie sind durch die Rostige Douglasienschütte orten möglicherweise Anbauflächen zuungunsten
so gefährdet, dass diese in Europa derzeit nicht an- der Buche hinzugewinnen. Eine wichtige Option
gebaut werden (Butin 2011). Die Buche ist von bioti- der Erhöhung der Trockenheitstoleranz der Buche
schen Schäden bisher vergleichsweise wenig betrof- kann die Einmischung von Herkünften aus Rand-
fen (Abs. 8.3.2); die mögliche Einwanderung bzw. regionen ihrer Verbreitung mit heute schon wär-
das Einführen neuer Schaderreger können hier die merem und trockenerem Klima sein. Die bessere
Situation aber schnell ändern (vgl. Abs. 8.3.2, Coul- Anpassung solcher Herkünfte an sich verändernde
ston et al. 2008). Nur auf den höher gelegenen Mit- Umweltbedingungen muss dabei aber vorher durch
tel- und Hochgebirgsstandorten mit einer bisherigen Tests bestätigt werden (Walentowski et al. 2009,
Kältelimitierung des Wachstums und auf Standorten Bolte & Degen 2010).
mit einer dauerhaft gesicherten, guten Wasserversor- Die Rolle der Eichenarten als mögliche „Ge-
gung können sich die Wuchsbedingungen durch die winner“ des Klimawandels in Deutschland ist an
Erwärmung verbessern. die erfolgreiche Kontrolle der wärmeliebenden Ei-
Die unteren Anbaugrenzen der Fichte in den chenschädlinge gekoppelt, welche die zukünftige
Gebirgslagen erhöhen sich vermutlich deutlich, was Existenz und Produktivität der Eiche maßgeblich
zu einer erheblichen Reduktion der Fichtenanbau- bestimmen können (s. Abs. 8.3.2); Ähnliches gilt
fläche insgesamt führen dürfte. Die Anbauoptionen auch für die Wald-Kiefer.
der Fichte im Tiefland könnten weitgehend verlo- Für die Wald-Kiefer und die Fichte stellt sich
ren gehen. Nutznießer dieser Entwicklung kann bei die Frage, wie sich eine mögliche Wintererwär-
einer erfolgreichen Begrenzung der Erwärmung auf mung und verstärkte Sommertrockenheit auf den
maximal 2 °C die Buche sein, die bisherige kälte- internen Kohlenstoff-Haushalt mit seinen Kom-
bedingte Standorte der Fichte in Form von Rein- ponenten Assimilation, Photosynthese und Ver-
beständen oder Beimischung in Mischbeständen atmung auswirken wird. Störungen der Kohlen-
besetzt und so ihre Anbaufläche und Wuchskraft stoffgewinne einerseits und erhöhte Dunkel- und
erhöht. Allerdings zeigen die meisten Klimaszena- Wurzelatmung andererseits bergen die Gefahr ei-
rien eine stärkere Erwärmung (Kap. 2) und auch bei nes langfristigen Kohlenhydrat-Defizits und des
geringfügiger Erwärmung steigen die Risiken für „Verhungerns“ („carbon starvation“, McDowell et
Schäden durch episodische Hitze- und Trocken- al. 2008, Allen et al. 2010). Submediterrane Klima-
perioden (Bolte et al. 2009a). Auch das Risiko für te mit milden Wintern und guten Bedingungen für
Schäden durch Spätfröste kann steigen, weil die Ve- eine Schädlingsausbreitung können daher mögli-
getationsperiode früher beginnt und lokale Popula- cherweise Grenzen für die erfolgreiche Anpassung
tionen an einen bestimmten Jahresgang der Tempe- der Wald-Kiefer an den Klimawandel darstellen. In
ratur angepasst sind (Bolte & Ibisch 2007, Kreyling Bezug auf die zukünftigen Eignung der Wald-Kiefer
et al.2011, 2012). unterscheiden sich die Einschätzungen der nord-
Die hohe Plastizität der Buche spricht dafür, deutschen forstlichen Versuchsanstalten („wenig
dass sie in weiten Gebieten auch künftig eine wich- anfällig“) von denen der süddeutschen („anfällig“)
tige Haupt- und Mischbaumart bleiben wird, falls (Bolte et al. 2009b).
die Klimaerwärmung und Trockenheitsverschär- Große Hoffnungen bei der Anpassung an den
fung in einem begrenzten Rahmen bleibt. Über die Klimawandel werden von der Forstpraxis in die
Grenzen der Trockenheitstoleranz der Buche wird Douglasie gesetzt (vgl. Kölling 2008). Im Fokus des
allerdings kontrovers diskutiert (Rennenberg et al. Interesses stehen bei dieser in Nordamerika sehr
2004, Ammer et al. 2005, Kölling et al. 2005, Bolte weit verbreiteten Art küstenfernere Herkünfte der
176Folgen des Klimawandels für die Biodiversität in Wald und Forst
Küstendouglasie, bei denen eine hohe Toleranz ge- häufig auch Effekte einer geänderten Landnutzung
genüber Sommertrockenheit (s. Tab. 8.2) mit ei- wie z. B. der Aufgabe der Almweide eine Rolle, die
ner ausreichenden Resistenz gegenüber biotischen sich deutlicher auf das Ansteigen der Waldgrenze
Schaderregern (Stephan 1973) kombiniert ist. Ein auswirkt als der Klimawandel (Gehrig-Fasel et al.
Problem ist allerdings, dass viele europäische Popu- 2007).
lationen nicht mehr in ihre Quellgebiete zurückver- Die Unterstützung der natürlichen Ausbreitung
folgt werden können. bzw. „Wanderung“ (Migration) von Baumarten
Die Betrachtungen machen klar, dass der Kli- („assisted migration“) durch die Beseitigung von
mawandel zu einer Änderung der Habitate der do- Ausbreitungshindernissen wie Habitatfragmentie-
minierenden Baumarten in bisher unklarem Um- rung und die Duldung natürlicher Sukzessionen,
fang und zu einer Ablösung von Waldgesellschaf- aber auch durch das gezielte Versetzen von Popula-
ten (Sukzession) unter Beteiligung spontan einwan- tionen wird zwar kontrovers diskutiert aber nichts-
dernder Baumarten, insbesondere aus dem Süden, destotrotz als ein wichtiges Element eines adaptiven
führen kann (Walentowski et al. 2009). Die spon- Waldmanagements angesehen (Millar et al. 2007,
tane Etablierung und Ausbreitung einwandernder Bolte & Degen 2010).
Baumarten ist allerdings ein langwieriger Prozess,
der von der Änderung der Umweltbedingungen, 8.4.1 Interaktionen mit Luftschadstoffen
der Samenverbreitung der einzelnen Arten und
der Konkurrenzkraft der etablierten Arten abhängt Wälder in Mitteleuropa unterliegen bereits seit Lan-
(Meier et al. 2012). Insbesondere die teilweise oder gem erhöhten Stoffeinträgen aus Emissionen. Diese
gänzliche Beseitigung der bestehenden Vegetation sind seit dem 19. Jahrhundert relevant und als Fol-
durch Störungen wie z. B. Windwurf, Trockenschä- ge der Industrialisierung, der Verkehrsentwicklung
den, Waldbrand oder Schädlingsbefall spielen eine und zunehmender Intensivierung der Landwirt-
wichtige Rolle bei einer gegenseitigen Ablösung von schaft anzusehen. Die Einträge verändern die che-
Waldgesellschaften (Sukzession). Aktuelle Beispiele mische Umwelt und damit die Lebensbedingungen
hierfür sind die Ausbreitung der Flaumeiche (Quer- der einzelnen Pflanzen mit Folgen für die Biodiver-
cus pubescens) als Baumart der submediterranen sität.
Breiten in absterbenden Wald-Kiefernbeständen Schädigende Einträge in Wälder wurden zu-
im schweizerischen Wallis (z. B. Rigling et al. 2006 nächst lokal als sogenannte „Rauchschäden“ spür-
a, b, Weber et al. 2007, vgl. auch Kasten I) und die bar. In den Zeiten der Frühindustrialisierung spiel-
Ausbreitung der Buche als nemorale Art in der von ten direkte Schäden an den Nadeln und Blättern
Fichte dominierten hemiborealen Zone Südschwe- der Bäume durch schwefelsaure Rauchgase nahege-
dens nach Windwurf, Trockenheit und Borkenkä- legener Emittenten (z. B. Metallhütten) eine wichti-
ferbefall (vgl. Bolte et al. 2010, Grundmann et al. ge Rolle (Schröder 1883, Burschel 1989).
2011). Seit den 1970er Jahren wurden dagegen auch
Die Häufung von Absterbeereignissen und Stö- in industriefernen Regionen Kronenschäden beob-
rungen an der trockenheits- und wärmebestimm- achtet, die durch den Ferntransport von Luftschad-
ten, meist südlichen Verbreitungsgrenze der Bau- stoffen ausgelöst wurden. Diese besonders in den
marten („rear edge“, Hampe & Petit 2005) deutet Höhenlagen der Mittelgebirge zunächst an Weiß-
auf eine bereits sichtbare großräumige Verschie- Tanne und Fichte, später an allen Hauptbaumar-
bung der Habitate einzelner Baumarten durch Kli- ten beobachteten sog. „neuartigen Waldschäden“
mawandel nach Norden hin (Allen et al. 2010). führten in ihrer gebietsweise großflächigen Symp-
Auch der Anstieg der Waldgrenze im europäischen tomatik und Intensität ab dem Beginn der 1980er
Gebirgsraum und die vertikalen Ausbreitungsten- Jahre zu der Furcht vor einem „Waldsterben“ und
denzen heimischer Baumarten wie der Buche zuun- dem Beginn einer umfassenden Waldökosystem-
gunsten der Fichte im Bayerischen Wald ist in ge- forschung, eines Waldschadensmonitorings sowie
wissen Grenzen der Erwärmung und Verschiebung politischer Anstrengungen zur Verminderung der
der Höhenzonierung der Vegetation zuzuschrei- Luftschadstoffbelastung (u. a. Genfer Luftreinhalte-
ben (s. Kasten II). Allerdings spielen in den Alpen konvention; vgl. Seidling & Bolte 2009). Die „neu-
177Biodiversität und Klimawandel
artigen Waldschäden“ wurden nicht mehr nur als gern, weil die Leitfähigkeit der Spaltöffnungen
Baumschädigungen angesehen, sondern als umfas- in einer CO2-angereicherten Atmosphäre sinkt
sende Beeinträchtigung der Wälder mit ihren öko- (Ainsworth & Rogers 2007, Leuzinger & Körner
systemaren Leistungen und ihrer Artenzusammen- 2007, Sitch et al. 2007). Dieser Effekt scheint je-
setzung (z.B. Wittig et al. 1985), und damit auch der doch bei Koniferen kaum oder gar nicht aufzu-
Biodiversität. treten (Körner et al. 2007).
Als wichtige Ursachen der „neuartigen Wald- (2) Eine Erwärmung kann die Verwitterungsraten
schäden“ wurden (1) eine direkte Schädigung der der Oberböden erhöhen, mit positiven Effekten
Assimilationsorgane durch saure Schwefelimmissi- auf die Pufferung eingetragener Säuren; ande-
onen (Wentzel 1979, 1982) und Ozon (Bucher 1982, rerseits wird die Mineralisierung der organi-
1984), (2) Bodenversauerung, Basenauswaschung schen Bodenkomponenten (bei ausreichender
und Freisetzung von Aluminium und Schwerme- Feuchte) angeregt, mit negativen Folgen einer
tallen in der Bodenlösung mit nachfolgenden Wur- möglicherweise erhöhten Stickstofffreisetzung,
zelschädigungen und Ernährungsstörungen (Ulrich -auswaschung und Bodenversauerung.
1986, 1995) sowie (3) eutrophierende Stickstoff- (3) Stickstoffeinträge können im Zusammenspiel
einträge mit nachfolgenden Ungleichgewichten mit einer Erwärmung das Baumwachstum po-
im Nährstoffangebot identifiziert (Nihlgård 1985). sitiv beeinflussen, wenn andere Ressourcen wie
Eine vermittelnde Ursachenerklärung lieferte die Wasser und übrige Nährstoffe ausreichend zur
sog. „Stresshypothese“ (Schütt et al. 1984, ähnlich Verfügung stehen; Ozon dagegen hemmt eher
zu Manion 1981), die neben der Wirkung von Luft- die Assimilation und wirkt einer Wachstums-
verunreinigungen auch biotische und klimatische steigerung durch Erwärmung entgegen (King et
Einflüsse als Stressoren mit einbezog. al. 2005, Sitch et al. 2007).
Im Verlauf der „Waldschadensdiskussion“ der (4) Allgemein verändern Stoffeinträge die individu-
letzten fast 30 Jahre, die sehr stark medial überprägt elle Empfindlichkeit von Organismen und Arten
und nicht immer wissenschaftlich geführt wurde, gegenüber Störungen, die im Zuge des Klima-
trat immer mehr der Klimawandel mit seinen Kom- wandels verstärkt erwartet werden. Stickstoffeu-
ponenten Erwärmung, Trockenheit und Sturmge- trophierung erhöht die Anfälligkeit gegenüber
fahr in den Vordergrund und die Bedeutung der Spätfrost, Trockenheit und dem Befall durch
Immissionen dahinter zurück. Bisher wurde den Pathogene und Parasiten. Auch die Sturmfestig-
Interaktionen zwischen Klima- und Witterungs- keit der Wälder kann durch Stoff-einträge ver-
effekten und Schadstoffeinträgen vergleichsweise mindert sein. So haben Bodenversauerung bzw.
wenig Aufmerksamkeit geschenkt und es existieren geringe Basensättigung der Böden und Stick-
noch viele Kenntnislücken (Paoletti et al. 2007). stoffeutrophierung nach Braun et al. (2003) die
Bytnerowicz et al. (2007) stellten in einem Sturmschäden durch den Orkan „Lothar“ in der
Übersichtsartikel den Forschungsstand zu inter- Schweiz erhöht. Eine geringere Tiefenerschlie-
aktiven Effekten von Schadstoffeintrag und Klima- ßung der Durchwurzelung (vgl. Bodenversau-
wandel zusammen: erungshypothese nach Ulrich, s.o.) und ein er-
(1) Die direkte phytotoxische Wirkung von Ozon höhtes Auftreten wurzelschädigender Pilze bei
sowie von hoch konzentriertem SO2, NH3 und erhöhten Stickstoffeinträgen (Buche: Phytoph-
NOx wird einerseits von der Länge der Vegeta- tora ssp., Fichte: Heterobasidion annosum) sol-
tionsperiode und damit von der Einwirkungs- len die Ursachen sein. Szenarien der Emissions-
zeit und andererseits vom Öffnungsgrad und entwicklung von Stickstoffverbindungen in Eu-
der Öffnungsdauer der Spaltöffnungen beein- ropa bis 2020 (Winiwarter & Hettelingh 2011)
flusst. Eine verlängerte Vegetationszeit im Zuge deuten darauf hin, dass die Freisetzung von NH3
des Klimawandels verstärkt die Wirkung der annährend gleich bleibt, während sich der NOx-
Schadstoffexposition, zunehmende Trocken- Ausstoß vermindert. Damit bliebe die flächende-
heit verringert dagegen ihre Wirkung. Auch die ckende N-Eutrophierungsgefahr weiterhin groß,
erhöhte Konzentration von CO2 in der Atmo- während sich die versauernde Wirkung des N-
sphäre könnte die Schadstoffexposition verrin- Eintrags insgesamt abschwächt.
178Folgen des Klimawandels für die Biodiversität in Wald und Forst
8.4.2 Abiotische und biotische Störungen modernen Forstwirtschaft hat der Mensch durch
vielfältige Maßnahmen versucht, Einfluss auf das
Störungen, also zeitlich begrenzte, oft abrupte Aus- jeweils aktuell bestehende Gleichgewicht zwischen
lenkungen von Umweltfaktoren, ob abiotisch oder Waldökosystemen und Umweltfaktoren zu Gunsten
biotisch, sind in Waldökosystemen systemimma- seiner forstwirtschaftlichen Zielsetzung zu nehmen.
nent (Otto 1994) und tragen maßgeblich zur Ent- Die vergleichsweise hohe Stabilität der Umweltfak-
wicklung und zum Erhalt der Biodiversität in Wäl- toren in den vergangenen 200 Jahren ermöglichte
dern bei (Jentsch et al. 2002). Dies erscheint zu- den Aufbau einer nachhaltigen Forstwirtschaft. Ak-
nächst überraschend, da wir Wälder als stabile, tive Waldschutzmaßnahmen minimierten Schäden
langlebige Ökosysteme wahrnehmen. Entscheidend und ermöglichten den Anbau von Baumarten auch
ist die zu beachtende Zeitskala. Oft sind es Ereignis- außerhalb ihres natürlichen Verbreitungsgebietes.
se, die im Abstand von mehr als einem Jahrhundert Der aktuelle Bericht von UNECE (United Na-
stattfinden, welche ganz entscheidend die natürli- tions Economic Commission for Europe) und FAO
chen Eigenschaften von Waldökosystemen bedin- (Food and Agriculture Organisation of United Na-
gen. Zeitliche und räumliche Diskontinuität eröff- tions) zum Zustand der europäischen Wälder gibt
net gerade bei langlebigen Ökosystemen sozusagen den Anteil der geschädigten Waldbestände inner-
„windows of opportunity“ beispielsweise für die Re- halb der EU mit gut 6 % der Gesamtwaldfläche an.
generation etablierter Arten und auch für das Hin- Die Bedeutung von Insekten und Krankheiten als
zutreten neuer. Bleiben Störungen ganz aus bzw. häufigste beobachtete Schadfaktoren wird dabei be-
werden diese kontinuierlich durch den Menschen tont (Köhl & San-Miguel-Ayanz 2011).
unterbunden, so führt dies zum Verlust von Diver- In Folge des Klimawandels werden die Um-
sität und von Funktionalität. Erhöht sich aber die weltfaktoren voraussichtlich variabler werden ( s.
Störungsfrequenz (Häufigkeit) oder auch die Inten- Kap. 2). Störungen von Waldökosystemen werden
sität von Störungen (ihre Magnitude), dann kann höchst wahrscheinlich zunehmen und die nachhal-
dies ebenfalls zu substanziellen Veränderungen und tige Bewirtschaftung der Wälder erschweren. Insbe-
zu Funktionsverlust führen. sondere Insekten und Pilze werden, aufgrund ihrer
Erst bezüglich der forstwirtschaftlichen Nut- schnellen Anpassungsfähigkeit, zu Schlüsselfakto-
zung der Wälder wird eine ökologische Störung zu ren der zukünftigen Waldentwicklung werden.
einem Schaden und der diesen auslösende Faktor
zu einem Schadfaktor. Eine solch anthropozentri- 8.4.2.1 Wind- und Sturmwurf:
sche Sichtweise ist entsprechend der multifunktio- Im vergangenen Jahrzehnt traten in Deutschlands
nalen Ansprüche der modernen Gesellschaft an den Wäldern beträchtliche Schäden durch Sturmtiefs,
Wald nachvollziehbar und wird durch die steigende lokale Gewitterstürme und Tornados auf. Am
Nachfrage nach nachwachsenden Energieträgern in schwerwiegendsten waren die Sturmtiefs Lothar
Zukunft noch an Bedeutung gewinnen. Auch der 1999 und Kyrill 2007, die allein in Deutschland zu
Aspekt der Planungssicherheit spielt eine Rolle. einem Schadholzanfall von jeweils mehr als 30 Mil-
Allerdings ist zu beachten, dass die Unterbin- lionen m3 führten. Dies entspricht mehr als 40% des
dung zeitlicher Dynamik und das Verhindern von gesamten jährlichen Nutzholzeinschlags in ganz
Störungen negative Langzeiteffekte mit sich brin- Deutschland (Jahresdurchschnitt 2002 bis 2008:
gen können. Bei ausbleibenden oder menschlich 70,5 Millionen m3, Oehmichen et al. 2011). Selbst
unterdrückten systemimmanenten Störungen wer- diese Extremereignisse waren allerdings vergleich-
den Dominanzbestände einzelner Arten gefördert. bar mit Sturmereignissen, die in der Vergangenheit
Solche monodominanten Bestände besitzen oft eine auftraten und können nicht als Beleg für eine Zu-
geringe Resilienz. In der Folge können dann even- nahme von Sturmereignissen oder deren Intensität
tuell nicht zu kontrollierende bzw. zu verhindernde durch den Klimawandel in der Vergangenheit gel-
Störungen besonders katastrophal wirken. ten (DWD 1999). Auch der erkennbare Trend einer
Grundsätzlich ist die Ökosystemstabilität an Zunahme der Schadholzmengen durch Sturmwürfe
die Stabilität der Umweltfaktoren gebunden (Mül- kann nicht als Hinweis gelten, da hierbei nicht nur
ler-Dombois & Ellenberg 1974). Mit Beginn der das Klimasignal, sondern auch die Anfälligkeit der
179Sie können auch lesen

















































