Das Makrozoobenthos als Indikator von anthropogenen Beeinträchtigungen an tropischen Bächen (La Gamba, Costa Rica)
←
→
Transkription von Seiteninhalten
Wenn Ihr Browser die Seite nicht korrekt rendert, bitte, lesen Sie den Inhalt der Seite unten

Das Makrozoobenthos als Indikator von
anthropogenen Beeinträchtigungen an
tropischen Bächen
(La Gamba, Costa Rica)
Diplomarbeit
ZUR ERLANGUNG DES AKADEMISCHEN GRADES
Magistra der Naturwissenschaften
Ausgeführt unter der Anleitung von
ao. Univ.-Prof. Mag. Dr. Leopold Füreder,
Institut für Ökologie
LEOPOLD-FRANZENS-UNIVERSITÄT INNSBRUCK
vorgelegt von
Verena Gertrud Duschek
Innsbruck, Jänner, 2020
Mein besonderer Dank gilt...
...ao. Univ.-Prof. Mag. Dr. Leopold Füreder, der mir diese Arbeit ermöglichte und ohne
dessen fachliche Unterstützung diese Arbeit nicht möglich gewesen wäre.
…Monika Springer für die fachliche Unterstützung und die engagierte Hilfe bei der
Bestimmung des Makrozoobenthos.
… “Rainforest Luxemburg” und COBIGA für die Finanzierung dieses Projektes.
… dem International Relations Office der Universität Innsbruck für das KWA-Stipendium.
…meinen Freunden, die während des Studiums trotz meiner längeren Auslandsaufenthalte
immer für mich da waren.
…allen, die meinen Aufenthalt in Costa Rica so unvergesslich gemacht haben. Allen voran
meinen Zimmerkolleginnen Jenny und Chiara, meinem Laborbuddy Randy und den Zivis
Simon und Matti. Gracias a todos los trabajadores de la estación para los bailes, las risas, las
fiestas, las excursiones, la ayuda… en fin gracias por todo el tiempo que compartimos juntos.
¡Sin ustedes no hubiera sido la misma experiencia!
...vor allem aber meiner Familie, die während meines Studiums immer hinter mir stand und
mich finanziell als auch persönlich immer unterstützte.
... meinem Papa, der meiner Arbeit den letzten grammatikalischen und sprachlichen
Schliff verliehen hat.
…meiner Oma für die kostenlose Verpflegung und den Schlafplatz, wenn ich zu faul war, nach
Landeck zu fahren.
…allen, die in irgendeiner Weise meinen Weg beeinflusst haben.
¡Gracias y Pura Vida!
2
Zusammenfassung
Costa Rica ist ein Land in Zentralamerika mit einer Vielzahl verschiedener Ökosysteme, das
trotz seiner geringen Größe ca. 5% der weltweiten Artenvielfalt beherbergt und somit als
wichtige Landbrücke zwischen Nord- und Südamerika gilt. Vor allem im 20. Jahrhundert litt
die Natur Costa Ricas unter der sich rasch entwickelnden Bevölkerung, die Primärwälder für
Siedlungsgebiet und landwirtschaftliche Flächen (Weideland und Plantagen) abholzte und
natürliche Lebensräume zerstörte. Um den potenziellen Einfluss anthropogener Landnutzung
auf tropische Bäche zu untersuchen, wurden 36 Makrozoobenthos-Proben aus bewaldeten und
beeinträchtigten Gebieten dreier Bäche nahe der Tropenstation La Gamba genommen und
ausgewertet. Neben der Erfassung von Abundanz und Diversität des Makrozoobenthos und der
Zusammensetzung der Makrozoobenthos-Gemeinschaft als auch der „Functional Feeding
Groups“, wurde die Wasserqualität anhand des BMWP´-CR-Index evaluiert. Trotz der hohen
Indexwerte, die für alle 6 untersuchten Standorte exzellente Wasserqualität ergaben, zeigten
sich signifikante Unterschiede in der Abundanz und der Makrozoobenthos-
Gemeinschaftsstruktur. Beeinträchtigte Gebiete wiesen deutlich höhere Individuenzahlen
toleranter Taxa (z.B. Chironomidae und Oligochaeta) auf, kennzeichneten sich aber dennoch
durch eine geringere Anzahl an Familien und einer geringeren Anzahl sensibler Taxa. Die
Abundanz des Makrozoobenthos in beeinträchtigten Gebieten spiegelt sich auch in der
Zusammensetzung der „Functional Feeding Groups“ wider. Collector-gatherers, Filterers und
Piercers waren tendenziell häufiger an beeinflussten Standorten; für Shredders, Predators und
Scarpers konnten keine klaren Muster festgestellt werden. Die Unterschiede waren an zwei
Bächen (Quebrada Negra und Quebrada Bolsa) stärker ausgeprägt als am dritten (Quebrada
Gamba), was zum Schluss führt, dass Ufervegetation anthropogenen Einfluss deutlich
reduzieren kann. Weiters konnte gezeigt werden, dass die Art der Landnutzung Einfluss auf die
Makrozoobenthos-Gemeinschaftsstruktur hat. Die Aufklärung der Bevölkerung und die
Etablierung von Waldpufferzonen sind essentiell, um Fließgewässer nachhaltig zu schützen und
um eine sukzessive Verschlechterung zu verhindern.
3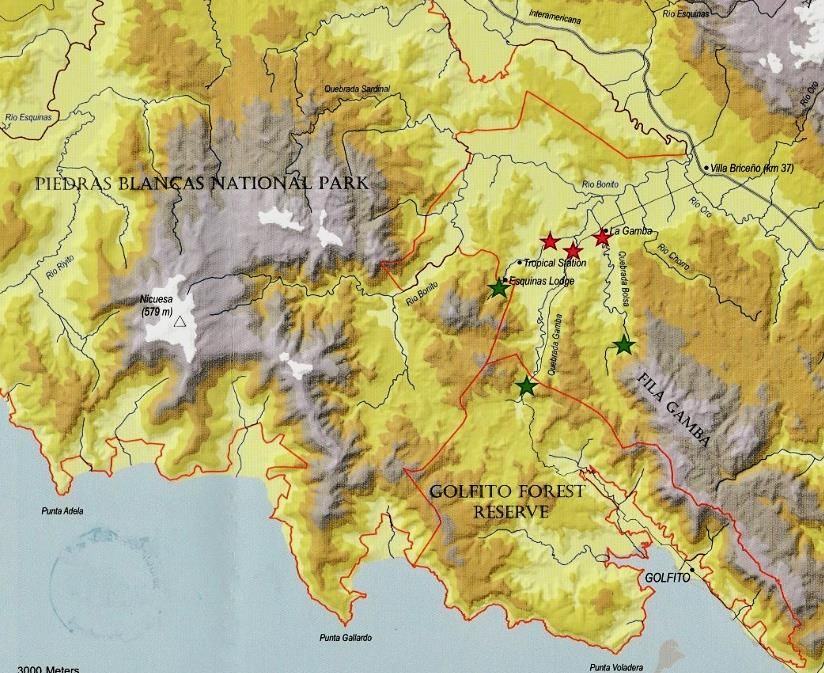
Abstract
Costa Rica is a country in Central America with a multitude of different ecosystems. Despite
its small size, it is home to about 5% of the world's biodiversity and is therefore considered an
important land bridge between North and South America. Especially in the twentieth century,
Costa Rica's nature suffered from the rapidly developing population, which deforested primary
forests and destroyed natural habitats for settlement and agricultural land (pasture and
plantations). To investigate the potential influence of anthropogenic land use on tropical
streams, 36 macroinvertebrates samples were taken and evaluated from forested and impacted
areas of three streams near the tropical research station La Gamba. In addition to recording the
abundance and diversity of macroinvertebrates and the composition of the macroinvertebrate
community, as well as the functional feeding groups, the water quality was evaluated using the
BMWP´-CR-Index. Despite the high index values, which showed excellent water quality for
all 6 investigated sites, there were significant differences in abundance and macroinvertebrates
assemblage structure. Impacted areas showed significantly higher numbers of individuals of
tolerant taxa (e.g. Chironomidae and Oligochaeta), but were nevertheless characterized by a
smaller number of families and a smaller number of sensitive taxa. The higher abundance of
aquatic macroinvertebrates in affected areas is also reflected in the composition of the
functional feeding groups. Collector-gatherers, Filterers and Piercers tended to be more
abundant at affected sites; for Shredders, Predators and Scarpers no clear patterns could be
found. The differences were more pronounced in two streams (Quebrada Negra and Quebrada
Bolsa) than in the third (Quebrada Gamba), leading to the conclusion that riparian vegetation
can significantly reduce anthropogenic influence. Furthermore, it could be shown that the type
of land use has an influence on the macroinvertebrate community structure. Environmental
education of the population and the establishment of forest buffer zones are essential for the
protection of watercourses and to prevent a successive deterioration.
4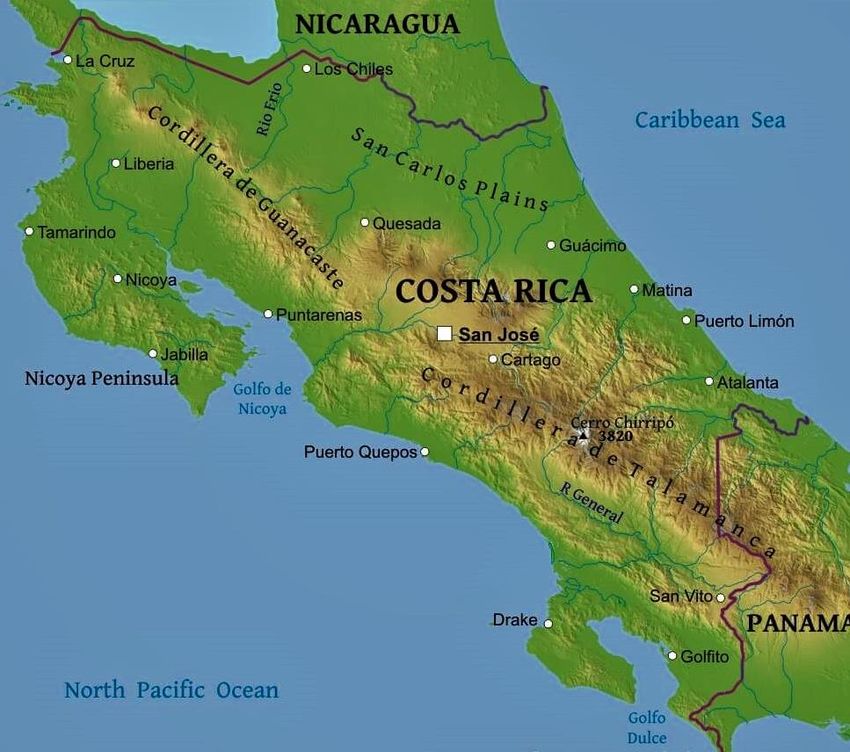
Inhaltsverzeichnis
Zusammenfassung .......................................................... 3
Abstract ........................................................................... 4
1. Einleitung.................................................................... 7
1.1. Klima ........................................................................................................................... 7
1.2. Abholzung der Primärwälder....................................................................................... 8
1.3. Erforschung tropischer Bäche ..................................................................................... 9
1.4. Erforschung anthropogener Einflüsse auf Fließgewässer ........................................... 9
1.5. Das Makrozoobenthos als Bioindikator .................................................................... 10
1.6. Bisherige Arbeiten im Untersuchungsgebiet ............................................................. 10
2. Fragestellungen und Zielsetzung ............................ 11
3. Material und Methoden ........................................... 12
3.1. Untersuchungsgebiet ................................................................................................. 12
3.1.1. Quebrada Negra.................................................................................................. 14
3.1.2. Quebrada Gamba ................................................................................................ 16
3.1.3. Quebrada Bolsa .................................................................................................. 18
3.2. Abiotische Messungen ............................................................................................... 20
3.3. Makrozoobenthos-Sampling ...................................................................................... 20
3.4. Bestimmung des Makrozoobenthos........................................................................... 21
3.5. Der BMWP´- CR- Index............................................................................................ 22
3.6. EPT-Taxa ................................................................................................................... 23
3.7. „Functional Feeding Groups“ .................................................................................... 23
3.8. Datenanalyse .............................................................................................................. 25
5
4. Ergebnisse ................................................................. 26
4.1. Physikalisch-chemische Parameter ............................................................................ 26
4.2. Abundanz und Diversität ........................................................................................... 27
4.3. Zusammensetzung der benthischen Lebensgemeinschaften ..................................... 28
4.4. EPT Taxa ................................................................................................................... 33
4.5. BMWP´-CR ............................................................................................................... 34
4.6. „Functional feeding groups“ ...................................................................................... 36
5. Diskussion ................................................................. 39
6. Schlussfolgerungen und Empfehlungen ................. 45
7. Unterrichtseinheit .................................................... 46
Anhang........................................................................... 57
Literaturverzeichnis ..................................................... 63
Abbildungsverzeichnis.................................................. 70
Tabellenverzeichnis ...................................................... 71
Abkürzungsverzeichnis ................................................ 72
Eidesstattliche Erklärung ............................................. 73
6
1. Einleitung
Costa Rica ist ein kleines Land am Isthmus von Zentralamerika, liegt zwischen Nicaragua
(Norden) und Panamá (Südosten) und wird im Osten vom Karibischen Meer (Atlantik) und im
Westen vom Pazifischen Ozean begrenzt. Costa Rica bietet eine Vielzahl von Ökosystemen
wie z.B. tropische Wälder, Feucht- bzw. Sumpfgebiete, Flüsse, Meere und aktive Vulkane
(KOHLMANN et al. 2015). Mehr als die Hälfte des Landes ist mit Wald bedeckt und mehr als
25% des Landes sind geschützte Gebiete (z.B. Nationalparks, Wildschutzgebiete, etc.)
(INSTITUTO NACIONAL DE ESTADÍSTICA Y CENSOS 2015). Es ist eines der artenreichsten Länder
der Welt und beherbergt zwischen 4-5% der weltweiten Artenvielfalt (manche Forscher gehen
sogar von bis zu 10% aus) (OBANDO 2002), obwohl es mit nur 51 000 km² terrestrischer Fläche
nur 0,03% der Gesamtfestlandfläche ausmacht (INSTITUTO NACIONAL DE ESTADÍSTICA Y
CENSOS 2015). Diese hohe Vielfalt kann nicht nur in terrestrischen, sondern auch in marinen
und limnischen Lebensräumen gefunden werden. Die hohe Biodiversität kann unter anderem
durch die geografische Lage als Landbrücke zwischen Nord- und Südamerika, das tropische
Klima, seine zwei Küsten und die Gebirge, die für unterschiedlichste Mikroklimate sorgen,
erklärt werden. Die zwei zentralen Gebirgsketten sind die Cordillera Volcánica Central und die
Cordillera de Talamanca im Süden, die das Land längs durchziehen und somit eine natürliche
Barriere zwischen karibischer und pazifischer Seite bilden (ebd.). Die Flüsse im Osten
(karibische Seite) entwässern in den Atlantik, die im Westen (pazifische Seite) in den Pazifik
(WEISSENHOFER et al. 2008a). Der Esquinas Wald reicht von Meereshöhe bis zu 579m (Cerro
Nicuesa) (ebd.).
1.1. Klima
Das Klima in Costa Rica kann grob in drei Regionen unterteilt werden: die tropisch- feuchte
Atlantikregion, die zentrale intermontane Region und die tropische Pazifikregion, letztere weist
Regen- und Trockenzeit auf. Aufgrund der physisch-geografischen Gegebenheiten können
diese wiederum in weitere sieben sogenannte Basisregionen mit ihren jeweiligen Subregionen
unterteilt werden (INSTITUTO NACIONAL DE ESTADÍSTICA Y CENSOS 2015). Das Gebiet um die
Tropenstation La Gamba liegt demnach in der Region Pacífico Sur (Abb. 1). In der Region
Pacífico Sur ist die Trockenzeit meist sehr kurz (Dezember-März) oder in manchen Gebieten
nicht eindeutig vorhanden, wohingegen sich die Regenzeit über mehrere Monate (April-
November) mit intensiven, kräftigen Regen erstreckt. Im Gebiet der Tropenstation liegt die
Jahresdurchschnittstemperatur zwischen 26-28 Grad und der durchschnittliche
7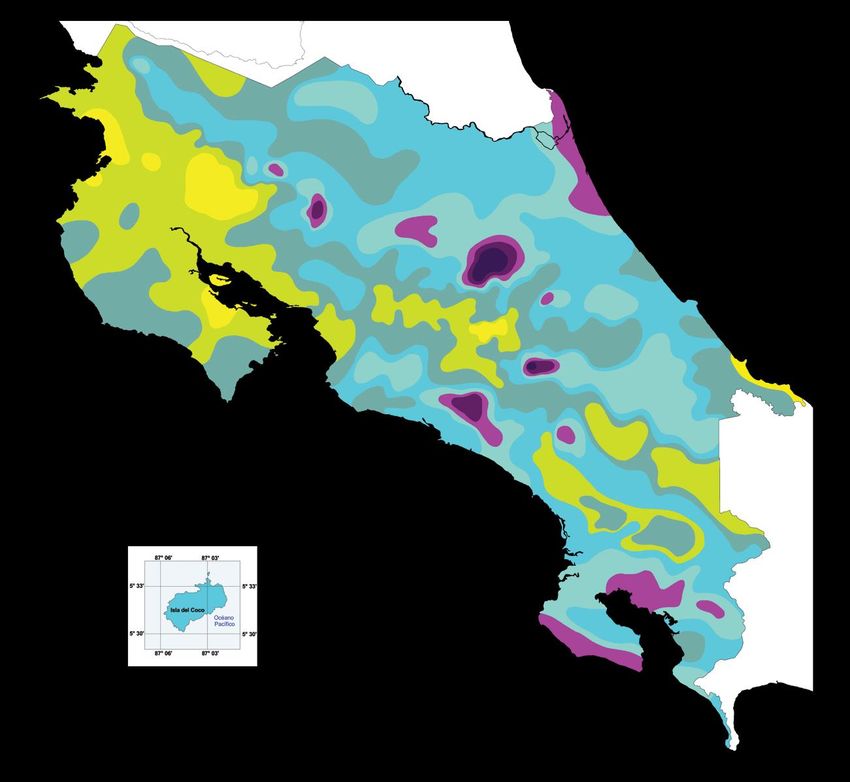
Jahresniederschlag bei über 5000mm (MINISTERIO DE AMBIENTE Y ENERGÍA, INSTITUTO
METEOROLÓGICO NACIONAL DE COSTA RICA 2009).
Abb. 1: Durchschnittlicher Jahresniederschlag in Costa Rica. Die Tropenstation (roter Stern) befindet sich in der Region
Pacifico Sur (schwarz umrandet). Entnommen und leicht verändert aus MINISTERIO DE AMBIENTE Y ENERGÍA, INSTITUTO
METEOROLÓGICO NACIONAL DE COSTA RICA (2009).
1.2. Abholzung der Primärwälder
Wie in vielen tropischen Regionen führten die rasche Entwicklung der Bevölkerung, das
schnelle Bevölkerungswachstum und die wirtschaftliche Lage vor allem im 20 Jahrhundert zur
Abholzung großer Waldflächen, um diese zu besiedeln und landwirtschaftlich zu nutzen. Die
günstigen Klimabedingungen in Costa Rica ermöglichen u.a. die Produktion von Kaffee,
Bananen, Palmherzen und Ananas (KOHLMANN et al. 2015), aber auch die Nutzung als
Weidefläche für Rinder ist vor allem in ländlichen Gebieten weit verbreitet (DUDGEON 2008,
CHARÁ-SERNA et al. 2015). Ende des 19 Jahrhunderts war vor allem die Pazifikküste von einer
Kolonialisierungswelle geprägt. In den frühen 1920er Jahren stieg die Abholzung exponentiell
8
an. Zwischen 1940 und 1986 verlor Costa Rica ca. 50% seiner Waldfläche, was zu erheblichen
Problemen wie Bodenerosion, Flussverlandung und Verschlechterung der Wasserqualität
führte. Aktuelle Entwaldungsraten sind relativ gering, da der Großteil des verbleibenden
Primärwaldes in Schutzgebieten liegt, dennoch steigt der Druck zur Nutzung dieser Gebiete/
von Holz und Wasserressourcen mit steigendem Bevölkerungswachstum (PRINGLE et al. 2016).
1.3. Erforschung tropischer Bäche
Erste Erforschungen tropischer Bäche und Flüsse begannen bereits Ende des 18. Jahrhunderts
mit Entdeckern und Naturforschern, die auf ihren Reisen aquatische Arten dieser Regionen
sammelten, wie z.B. von Humboldt und Bonpland (1799-1802) oder später Spix und von
Martius (1817-1820) oder Bates (1848-1859). Später, im 20 Jahrhundert, wurden
umfangreichere Kollektionen u.a. von professionellen Sammlern wie H. S. Parish und F.
Woytkowski erstellt (JACKSON & SWEENEY 1995). Die Anzahl der Publikationen pro Jahr zu
Forschungen in tropischen Bächen steigt seit den 1980er Jahren deutlich (ebd.); auch die Anzahl
publizierter Studien und das Interesse an Makroinvertebraten in Bächen und Flüssen in
lateinamerikanischen Ländern (RAMÍREZ & GUTIÉRREZ-FONSECA 2014a) und Costa Rica
(SPRINGER 2008, SPRINGER et al. 2014) wächst stetig an. Aktuell werden Makroinvertebraten
vor allem in Umweltstudien, Wasserqualitätsbewertungen und regelmäßigen
Gewässerüberwachungen, insbesondere von Flüssen und Bächen, verwendet (SPRINGER 2019).
1.4. Erforschung anthropogener Einflüsse auf Fließgewässer
Hunderte von Studien dokumentieren statistische Zusammenhänge zwischen anthropogener
Landnutzung und Gewässergegebenheit und liefern wichtige Beweise für die Bedeutung der
umgebenden Landschaft und deren Nutzung zur ökologischen Integrität eines Baches. Forscher
aus gemäßigten Gebieten haben die Auswirkungen der Abholzung und Landwirtschaft auf
Fließgewässer erforscht und u. a. erhöhten Sediment-, Nährstoff- und Pestizideintrag,
Änderungen des Durchflussregimes, und die Verschlechterung der Lebensräume an Ufern und
in Bächen festgestellt (ALLAN 2004). Die Auswirkungen der Entwaldung auf tropische
Flussökosysteme erfuhr vergleichsweise wenig Aufmerksamkeit, fand aber vor allem in den
letzten Jahren immer mehr Beachtung. Zahlreiche Forscher stellen u. a. Veränderungen der
physisch-chemischen Gegebenheiten und Nahrungsverfügbarkeit fest, was in der Folge zu
Verschiebungen in der Abundanz, Vielfalt und Gemeinschaftsstruktur des Makrozoobenthos
führt (LORION & KENNEDY 2009).
9
1.5. Das Makrozoobenthos als Bioindikator
Aquatische Makroinvertebraten, insbesondere die Insekten, sind von besonderer Bedeutung für
die Bestimmung der Gesundheit aquatischer Ökosysteme, da einige Gruppen aufgrund ihrer
Sensibilität gegenüber Wasserverschmutzung und Lebensraumveränderung als verlässliche
Bioindikatoren für die Wasserqualität gelten (ROSENBERG 1993; SPRINGER 2010b). Aquatische
Insekten sind die häufigste und artenreichste Gruppe von Organismen in tropischen
Fließgewässern, sind in einer großen Vielfalt aquatischer Ökosysteme zu finden und erreichen
ihre höchste Diversität in sauberen, schnell fließenden Gebirgsflüssen und Bächen. Die
artenreichsten Gattungen in diesen Habitaten sind Köcherfliegen (Trichoptera), Zweiflügler
(Diptera) und die Käfer (Coleoptera) (SPRINGER 2008). Die wichtigste wissenschaftliche
Sammlung aquatischer Makroinvertebraten in Costa Rica befindet sich im Zoologischen
Museum der Universität von Costa Rica (MZUCR). SPRINGER (2019) bietet eine aktuelle Liste
der in der Kollektion vorkommenden Wasserinsekten, welche 98 Familien und 341
identifizierte Taxa aus elf Insektenordnungen auflistet.
1.6. Bisherige Arbeiten im Untersuchungsgebiet
Verglichen mit anderen Fachgebieten der Biologie liegen nur wenige
Fließgewässeruntersuchungen im und um den Nationalpark Piedras Blancas vor, obwohl sich
das Flussnetzwerk des Rio Esquinas geradezu anbietet, tropische Bäche zu erforschen.
SCHIEMER et al. (2010) bietet eine umfassende gewässerökologische Studie des Gebiets,
während TSCHELAUT et al. (2008) eine umfassende Beschreibung des Einzugsgebiets des Rio
Esquinas erarbeiteten. Über die Biodiversität und Ökologie des Makrozoobenthos in den
Flüssen und Bächen La Gambas ist bisher wenig bekannt: In FÜREDER et al. (2014) werden
hydromorphologische Gegebenheiten, abiotische Parameter und biologische Auswertungen für
die Beschreibung der aquatischen Biodiversität mehrerer Flüsse und Bäche der Region
herangezogen. Das Ergebnis ist ein Artikel, der die Vielfalt des Makrozoobenthos, der Fische,
Krebse und Garnelen der Region darstellt. TSCHELAUT (2005) beschäftigte sich einerseits mit
der Besiedlung von Makroinvertebraten verschiedener Blattarten und deren Abbau und
andererseits mit der Verteilung der Invertebraten und der funktionellen Zusammensetzung des
Makrozoobenthos in vier verschiedenen Habitattypen. In der Masterarbeit von HOLZER (2018)
geht es zwar primär um die Diversität und Verbreitung von Süßwassergarnelen in der Golfo
Dulce Region, dennoch wurden auch Proben des Makrozoobenthos genommen, ausgewertet
und interpretiert. Über die potenziellen Auswirkungen anthropogener Landnutzung auf die
Bäche und Flüsse gab es bisher noch keine ökologischen Studien im Untersuchungsgebiet.
102. Fragestellungen und Zielsetzung
Die vorliegende Arbeit befasst sich mit der Biodiversität aquatischer Invertebraten tropischer
Bäche unter dem Einfluss anthropogener Landnutzung. Das generelle Ziel dieser Studie war es,
Makrozoobenthos Proben und physikalisch-chemische Werte von Wald- und anthropogen
beeinflussten Standorten desselben Baches zu vergleichen, um den potenziellen anthropogenen
Einfluss darzustellen.
Daraus ergaben sich diese Fragestellungen:
1) Welche physikalisch-chemische Gegebenheiten herrschen in tropischen Fließgewässern?
2) Welche Makrozoobenthosgemeinschaften kommen in tropischen Fließgewässern vor?
3) Wie wirkt sich die anthropogene Landnutzung auf die physikalisch-chemischen
Gegebenheiten und die Zusammensetzung des Makrozoobenthos aus?
4) Welche Korrelationen gibt es zwischen physikalisch-chemischen Gegebenheiten und
Makrozoobenthosgemeinschaften?
Diese Fragen führten zu folgender Zielsetzung:
Ziel dieser Studie war es also herauszufinden, wie sich unterschiedliche anthropogene
Belastung auf physikalisch-chemische Gegebenheiten des Baches und auf (i) die Abundanz,
(ii) die Zusammensetzung und (iii) die Funktion des Makrozoobenthos auswirkt und welche
Faktoren dabei eine entscheidende Rolle spielen.
113. Material und Methoden
3.1. Untersuchungsgebiet
Die Untersuchung wurde in der Golfo Dulce Region, im Süden von Costa Rica, unweit der
Tropenstation La Gamba, im gleichnamigen Dorf durchgeführt. Die Region hat einen jährlichen
Niederschlag von 5.836 mm und gehört zu den feuchten Tieflandregenwäldern. Die jährliche
Durchschnittstemperatur liegt bei 28,2°C und die Luftfeuchtigkeit bei 88,3% (WEISSENHOFER
et al. 2008a).
Abb. 2: Costa Rica und die Golfo Dulce
Region mit dem Piedras Blancas
Nationalpark und dem Untersuchungsgebiet.
Die Beprobungsstandorte sind mit grünen
(Waldstandort) und roten (beeinträchtigter
Standort) Sternchen markiert. Abk.: QN
Quebrada Negra, QG Quebrada Gamba, QB
Quebrada Bolsa. Bilder bzw. Karten
entnommen und leicht verändert aus:
(1)https://www.wpclipart.com/geography/Co
untry_Maps/global_location/North_America/
Costa_Rica_on_globe.png.html
(2)https://www.freeworldmaps.net/centralam
erica/costarica/costarica-geography.jpg
(3) WEISSENHOFER et al. (2008c) .
QN
QB
QG
QN
QB
QG
12Der Forschungszeitraum erstreckte sich von Dezember 2018 bis Februar 2019, während der
Trockenzeit. Es wurden drei Bäche des Einzugsgebiets des Rio Esquinas (Abb. 2), welcher
schließlich in den Golfo Dulce (pazifischer Ozean) fließt, untersucht. Alle Bäche entspringen
in der Fila Gamba und fließen über in den Rio Bonito. Es wurden insgesamt sechs Proben aus
drei Flüssen entnommen. Die oberen Standorte liegen im Primärregenwald und sind keinen
anthropogenen Beeinträchtigungen ausgesetzt (Referenzstandorte), wohingegen die unteren
Standorte neben Weideland, Straße und Wohngebiet liegen (Tab. 1; QN: Abb. 3; QG: Abb. 6;
QB: Abb. 9). Letztere werden im Weiteren als beeinträchtigte/beeinflusste Standorte
bezeichnet.
Tab. 1: Koordinaten und Standortbeschreibungen der einzelnen Untersuchungspunkte. Die Koordinaten sind im Dezimalgrad
angegeben und wurden mit einem GPS- Gerät der Tropenstation La Gamba gemessen.
Fluss Koordinaten Standortbeschreibung
N 8,69981° Waldstandort: ca. 100 Meter flussaufwärts hinter der
Quebrada
W 83,20378° Tropenstation
Negra
N 8,70136° Beeinflusster Standort: große Weide nahe der Tropenstation;
(QN)
W 83,19829° am Weg Richtung Dorfzentrum auf der linken Seite
N 8,67207° Waldstandort: Zustieg bei der letzten Brücke auf dem Weg
Quebrada
W 83,19926° nach Golfito; Beprobung ca. 200m flussaufwärts
Gamba
N 8,70180 Beeinflusster Standort: Zustieg und Standort bei der letzten
(QG)
W 83,19158 Brücke der Nebenstraße von der Tropenstation ins Dorfzentrum
N 8,68882° Waldstandort: Standort ca. 1 km flussaufwärts ab der letzten
Quebrada
W 83,18017° Zustiegsmöglichkeit am Ende der Seitenstraße
Bolsa
N 8.70712° Beeinflusster Standort: Zustieg und Standort bei der Brücke
(QB)
W 83.18556° auf der Straße durch La Gamba
Die Referenzstandorte zeichnen sich durch ausgedehnte Bewaldung und folglich viel Schatten
aus, wohingegen bei den beeinträchtigten Standorten am Ufer nur vereinzelte Bäume relativ
wenig Schatten liefern. Die Bachlänge für die Makrozoobenthos-Probenentnahme betrug
zwischen 15 und 30 Meter, wobei jeweils drei Proben aus Riffle (Schnellen) und drei Proben
aus Pools (Gumpen) entnommen wurden.
13Die Sedimentgröße wurde visuell geschätzt. Dazu wurden die gängigen Korngrößen
Schluff/Sand (< 2 mm), Kies (2-63 mm), Geröll (63- 200 mm) und Blöcke (>200mm) gewählt
und deren prozentueller Anteil im Flussbett angegeben. Auch die Abschattung des Flusses wurde
visuell in 5% Schritten geschätzt. Alle Werte sind in folgender Tabelle zu finden:
Tab. 2: Geschätzter prozentueller Anteil der Sedimentgröße und der Abschattung aller sechs Standorte. Waldstandorte (W);
anthropogen beeinflusste Standorte (A).
Quebrada Negra Quebrada Gamba Quebrada Bolsa
W A W A W A
Sand 10% 30% 20% 30% 20% 40%
Kies 40% 60% 40% 50% 30% 50%
Sedimentgröße
Geröll 40% 10% 25% 20% 30% 10%
Blöcke 10% - 15% - 20% -
Abschattung 90% 10% 95% 50% 85% 5%
3.1.1. Quebrada Negra
Quebrada Negra ist ein Bach erster Ordnung, der im Schluchtwald auf etwa 180m Seehöhe
oberhalb der Esquinas Rainforest Lodge entspringt, an der Tropenstation vorbeifließt und nach
ca. 2,7km in den Quebrada Gamba mündet (FÜREDER et al. 2014). Er entwässert neben
Primärregenwald, auch Sekundärwald und Viehweiden, und ist, aufgrund der Nähe zur
Tropenstation, der am besten untersuchte Bach in La Gamba. In manchen Bereichen nahe der
Tropenstation wurde das Flussufer, aufgrund früherer landwirtschaftlicher Nutzung, befestigt
(vgl. HOLZER 2018)
Abb. 3: Waldstandort (links) und beeinflusster Standort (rechts) am Quebrada Negra.
14Der Referenzstandort befindet sich nur wenige 100 Meter von der Station entfernt. Er ist ein
Beispiel für einen natürlichen, dynamischen und unregulierten Flusslauf und weist das typische
Profil eines Quellflusses auf: steinig, flach, schmal und dichte Ufervegetation (TSCHELAUT et
al. 2008). Die Vegetation entspricht der eines schmalen Flusses mit Anpassungen an flache
Terrassen. Merkmale dieses Typs sind u.a.: hohe Bäume (bis zu 50m) mit massiven Stämmen
und Brettwurzeln und dichter Unterwuchs (FÜREDER et al. 2014). Der Standort der Beprobung
liegt im Bereich des alten Sekundärregenwalds (WEISSENHOFER et al. 2008b), wobei davon
ausgegangen wird, dass mittlerweile natürliche Bedingungen herrschen. Durch hineingefallene
Baumstämme (Abb. 5a) und in den Bach hängende Pflanzen und Wurzeln (Abb. 5c) bietet
dieser Bach eine Vielzahl an Habitaten für Makroinvertebraten. Häufige Hochwasserereignisse
prägen zusätzlich das Flussbett und sorgen für reich strukturierte Uferbereiche (FÜREDER et al.
2014).
Abb. 4: Referenzstandort Quebrada Negra: a: Riffle an im Bach liegenden Stamm; b: Probenahme; c: Wurzeln im Wasser
bieten zusätzliche Mikrohabitate.
Der beeinflusste Standort ist Teil einer Rinderweide und ist auch für die Tiere frei zugänglich.
Nur einzelne Bäume, die meisten der Gattung Senna, stehen am Ufer. Aufgrund der geringen
Ufervegetation gibt es kaum Schatten, weshalb die Sonneneinstrahlung sehr hoch ist. Obwohl
der Standort sehr nahe dem Referenzstandort liegt, ist das Bachbett deutlich breiter und die
Fließgeschwindigkeit deutlich langsamer; tiefe Pools reihen sich aneinander. In den Pools
wachsen teils dicht aneinander Wasserpflanzen (Abb. 4a). Starker Algenbewuchs trübt das
Wasser (Abb. 4b/c) und ist typisch für diesen Abschnitt.
Abb. 5: Weidestandort Quebrada Negra: a: Wasserpflanzen in einem kniehohen Pool; b und c: Starker Algenwuchs am
Probestandort.
153.1.2. Quebrada Gamba
Der Quebrada Gamba entwässert im Oberlauf primäres Regenwaldgebiet und wird im Mittel-
bzw. Unterlauf zunehmend anthropogen beeinträchtigt (u.a. Straße, Ölpalmplantage,
Viehweide). Er entspringt auf etwa 260m Seehöhe der Fila Gamba und windet sich über 7,7 km
ins Tal, wo er vorerst mit dem Quebrada Bolsa zusammenfließt (TSCHELAUT 2005) und
schließlich in den Rio Bonito mündet.
Abb. 6: Waldstandort (links) und beeinflusster Standort (rechts) am Quebrada Gamba.
Der Standort im Primärregenwald Abb. 6war vergleichsweise seicht und führte zum Zeitpunkt
der Beprobung wenig Wasser. Kleinere Kiesbänke an denen sich Laub ansammelte, waren im
Flussbett vorhanden (Abb. 7c) und wurden an mehreren Stellen durchflossen. Die
Ufervegetation entspricht der eines Schluchtwaldes und weist 30-40 m hohe Bäume und dichten
Unterwuchs auf (Abb. 7b). Weiters ist die Ufervegetation mit zahlreichen Lianen und
Epiphyten bewachsen (Abb. 7a).
Abb. 7: Waldstandort Quebrada Gamba: a: Epiphyten an Uferpflanzen; b: dichter Unterwuchs am Standort; c: Schotterinsel
im Bach.
16Der anthropogen beeinflusste Standort hat an beiden Uferseiten einen schmalen Baumgürtel,
der teils unterbrochen ist. Orographisch links befindet sich landwirtschaftlich genutzte Fläche
und rechts die Hauptstraße von La Gamba. Im Vergleich zum Referenzstandort führt der
Quebrada Gamba hier mehr Wasser und auch das Flussbett ist deutlich breiter, was
wahrscheinlich einerseits an der großen Distanz zwischen den beiden Standorten liegt und
andererseits am Zufluss mehrerer kleiner Bäche. Tiefe Pools wechseln sich mit seichten,
schnellerfließenden Riffeln ab (Abb. 8); Schotterbänke werden von Wasser um- bzw.
durchflossen. Im Vergleich zu den anderen beeinflussten Standorten wirkt dieser deutlich
naturbelassener.
Abb. 8: anthropogener Standort Quebrada Gamba: a & b: tiefe Pools am Probestandort; c: Riffle am Probestandort.
173.1.3. Quebrada Bolsa
Quebrada Bolsa ist ein tropischer Tieflandregenwaldbach zweiter Ordnung (lt. TSCHELAUT et
al. 2008; FÜREDER et al. 2014), welcher bogig verzweigt zuerst Primärregenwald,
Sekundärregenwald, landwirtschaftliche Flächen (z. B. Ölpalmplantagen) und schließlich
Siedlungsgebiet durchquert (vgl. WEISSENHOFER et al. 2008b). Im oberen Flusslauf sind oft
große Gesteinsbrocken und Baumstämme inmitten des Bachbetts, welche zahlreiche
Lebensräume für Lebewesen bieten. Der Primärwald in diesem Bereich reicht bis zum Ufer und
steht teils auf steilen Flanken (FÜREDER et al. 2014). Oft sind breite Schotterbänke vorhanden,
die von krautigen Pionierpflanzen besiedelt werden. In der Trockenzeit kann es in manchen
Teilen zur oberflächlichen Austrocknung des Flusses kommen.
Abb. 9: Waldstandort (links) und beeinflusster Standort (rechts) am Quebrada Bolsa.
18Der erste Standort befindet sich im Primärregenwald und wurde nach oben hin von
umgestürzten Bäumen begrenzt (Abb. 10a). Riffle und Pools wechselten sich gleichmäßig ab;
Uferpflanzen und andere Strukturen hingen teils ins Wasser und boten zusätzliche Lebensräume
(Abb. 10b/c). An einer Seite ragte eine steile Flanke mit dichtem Wald empor. Trotz der
umliegenden Bäume, die den Großteil des Baches beschatten, gibt es immer wieder Bereiche,
die aufgrund des breiten Flussbettes gut belichtet werden.
Abb. 10: Waldstandort Quebrada Bolsa: a: umgefallene Bäume am Probebach; Uferstrukturen (b) und Pflanzen (c) die ins
Wasser reichen.
Der anthropogen beeinflusste Standort befindet sich im Siedlungsgebiet von La Gamba. Häuser
entlang des Flusses werden nur durch einen schmalen Baumgürtel, der kaum Schatten bietet,
vom Fluss getrennt. Direkt über den Fluss führt eine Brücke (Abb. 11a), bei der man zum Fluss
steigen kann. Hunde als auch Kinder wurden des Öfteren beim Spielen am Fluss angetroffen
und bieten somit eine zusätzliche Beeinflussung. Die orographisch linke Seite des Flusses ist
durch Erosion geprägt (Abb. 11b). Weiter flussaufwärts verläuft der Fluss entlang einer
Ölpalmplantage, wobei auch hier das Ufer stark erodiert.
Abb. 11: beeinträchtigter Standort Quebrada Bolsa: a: Brücke am Probeort, b: erodiertes Ufer c: beschatteter Pool durch
Pflanzen der Gattung Senna.
193.2. Abiotische Messungen
Die Messungen wurden mit einem Gerät der Firma HACH aus der HQ- Serie (HQ40d)
durchgeführt. Es wurden Temperatur, PH- Wert, Leitfähigkeit und Sauerstoffkonzentration und
-sättigung gemessen. Da die LDO (Luminescent-Dissolved-Oxygen) - Elektrode nach den
ersten beiden Benutzungen nicht mehr richtig funktionierte, sind für Quebrada Gamba und
Quebrada Bolsa keine Daten zu Sauerstoffgehalt und -sättigung zum Untersuchungszeitpunkt
vorhanden. Die Angaben zu QB konnten durch Messungen der zoologischen Exkursion der
Universität Innsbruck im Februar ergänzt werden und sind dementsprechend gekennzeichnet.
Es wurden jeweils drei Messungen jedes Faktors pro Standort durchgeführt und der daraus
resultierende Mittelwert errechnet. Die einzelnen Messungen wurden mindestens fünf Meter
voneinander durchgeführt, um etwaige Unterschiede zu erfassen. Da das Gerät zwei Anschlüsse
hat, wurden zwei Messungen parallel durchgeführt; der dritte Messwert wurde, nachdem eine
Elektrode ausgetauscht wurde, im Anschluss erfasst. Der Temperaturwert wurde von der
Leitfähigkeitselektrode übernommen, da dieser der genaueste ist (Dechent Elias, persönliche
Mitteilung).
3.3. Makrozoobenthos-Sampling
An allen Standorten wurden quantitative Proben genommen; für QN und QB liegen zusätzlich
qualitative Daten der Exkursion der Universität Innsbruck vor, welche auch miteinbezogen
werden. In der ersten Dezemberwoche wurden die Probenstandorte ausgesucht, dabei war
wichtig, einen möglichst repräsentativen Abschnitt des Baches auszuwählen, wo sowohl Riffle-
, als auch Pool -Sektionen vorhanden sind. Im Laufe der weiteren Wochen wurden dann die
Proben an den verschiedenen Standorten genommen und ausgewertet.
Für das quantitative Sampling wurde ein 30x30cm großer Kicksampler mit einer Maschenweite
von 150μm verwendet. Insgesamt wurden 36 Proben genommen; pro Standort wurden jeweils
drei Riffle und drei Pools beprobt. Diese wurden mit der Absicht, möglichst viele Mikrohabitate
abzudecken, ausgesucht. Für das Sammeln des Makrozoobenthos (MZB), wurde kurz oberhalb
des Kicksamplers in einem ca. 90cm² großen Feld (Grundfläche des Samplers) der Boden des
Bachs von Hand bzw. Fuß für ca. eine Minute lang aufgewirbelt; die so „freiwerdenden“
Lebewesen wurden durch die Strömung in das Netz getrieben. In Pools musste mit der Hand
selbst eine Strömung Richtung Netz erzeugt werden. Große Steine in dem 90cm² Feld wurden
in einem weißen Kübel mit einer gewöhnlichen Küchenbürste abgeschrubbt und anschließend
zurück in den Bach gelegt. Der Inhalt des Kicksamplers wurde ebenfalls in den Kübel geleert.
20Dieser wurde dann geschwungen, um kleine Steine und Sand von organischem Material zu
trennen. Anschließend wurde das organische Material durch ein Handnetz gesiebt. Dieser
Vorgang wurde ca. 3mal wiederholt. Der Inhalt des Handnetzes wurde in kleine Plastiksäcke
abgefüllt und mit 80% Ethanol fixiert. Die Säcke wurden verknotet, in einen Plastikbehälter
gelegt und ins Labor transportiert.
Für das qualitative Sampling wurden mehrere Techniken angewandt: die Benützung eines
Handnetzes (Vorgang ähnlich wie beim Kicksampling) und das Sammeln von organischem
Material (wie z.B. Blatthaufen) in einer Schale. Alle Individuen wurden anschließend per
Pinzette aus dem Substrat gepickt. Das qualitative Samplen dauerte ca. 30 Minuten und
inkludierte sowohl die Zeit des Sammelns als auch die Zeit der Separierung des MZB und
wurde von vier Personen gleichzeitig durchgeführt. Die Individuen wurden mit 80% Ethanol in
Eppendorfer-Tubes fixiert und für die weitere Bestimmung ins Labor gebracht.
3.4. Bestimmung des Makrozoobenthos
Die quantitativen Proben wurden zum Teil in Costa Rica und zum Teil in Österreich am Institut
für Ökologie der Universität Innsbruck bestimmt und ausgezählt. In Costa Rica wurden die
Proben unter einer Stereolupe der Marke Leica von organischem und anorganischem Material
getrennt, in der Folge bestimmt und in Eppendorfer-Röhrchen in 80% Alkohol im Labor
gelagert. In Österreich erfolgte ein Zwischenschritt: Die Proben wurden vor dem Auswerten
durch ein 1000μm, 500 μm und 100μm Sieb gegeben: anschließend wurde das
Makrozoobenthos aus den 1000μm und 500μm Proben unter einem Auflichtmikroskop
aussortiert, bestimmt und in Glasröhrchen mit 80% Alkohol bei Raumtemperatur gelagert.
Alle gesammelten Makroinvertebraten wurden zur tiefstmöglichen taxonomischen Ebene,
meist Gattung oder Familie, bestimmt. Dazu wurden taxonomische Bestimmungsschlüssel aus
Macroinvertebrados de Agua Dulce de Costa Rica I (SPRINGER et al. 2010) und Keys to
neotropical hexapoda (ROGERS et al. 2018) verwendet. Zusätzlich wurden ROLDÁN PÉREZ
(1996) und WIGGINS (2015) herangezogen.
Die qualitativen Proben wurden in der Tropenstation mithilfe einer Stereolupe auf
Familienniveau bestimmt. Dabei wurden neben oben genannter Literatur unter anderem
sogenannte „Guías fotográficas“ von PABLO E. GUTIÉRREZ-FONSECA et al. (2016) und
VÁSQUEZ et al. (2010) genutzt.
213.5. Der BMWP´- CR- Index
Die “The Biological Monitoring Working Party” (BMWP) entstand 1978 und wurde zur
Bewertung von Flüssen in Großbritannien entwickelt. Später wurde der Index von ALBA-
TERCEDOR & SÁNCHEZ-ORTEGA (1988) für Spanien angepasst und schließlich 2007 für Costa
Rica (CR) modifiziert (SPRINGER 2010a). Der Index basiert auf der Anzahl der vorkommenden
Familien des Makrozoobenthos und deren zugewiesenen Toleranz-Punkte (von 1 bis 9). Weder
die Anzahl der Individuen, noch die der Gattungen spielen eine Rolle (ebd.). Die Punkteanzahl
jeder Familie ist im DECRETO NO. 33903- MINAE-S. (17.09. 2007) gelistet. Tab. 3 zeigt den
Grad der Wasserqualität, die dazugehörigen Indexzahlen und die Farbe jeder Kategorie. Zu
beachten ist, dass Familien, die im Pelagial leben (z.B. Gerridae oder Veliidae) nicht zum
Makrozoobenthos zählen und deshalb nicht zur Berechnung der Wassergüte herangezogen
werden.
Tab. 3: Klassifizierung der Wasserqualität anhand der Gesamtpunktezahl des BMWP´-CR- Indexes entsprechend der
Makrozoobenthos- Familien und die zugehörige Referenzfarbe.
Grad der Wasserqualität Index Wert: Farbe
BMWP´- CR
Exzellente Wasserqualität >120
Sehr gute Wasserqualität 101-120
Gute Wasserqualität, leicht kontaminiert 61-100
Schlechte Wasserqualität, kontaminiert 36-60
Sehr schlechte Wasserqualität, sehr kontaminiert 16-35
Extrem schlechte Wasserqualität, extrem kontaminiert3.6. EPT-Taxa
Ephemeroptera, Plecoptera und Trichoptera (kz. EPT Taxa) gelten als relativ sensible Gruppen
betreffend der Wasserqualität (vgl. u.a. SPRINGER 2010a; MAFLA 2005; AB HAMID & MD RAWI
2017) und werden daher häufig als Bioindikatoren für aquatische Systeme genutzt. In der
vorliegenden Arbeit werden die taxonomische Vielfalt, Abundanz und der relative Anteil der
EPT-Taxa an den Probestandorten betrachtet.
3.7. „Functional Feeding Groups“
Die “functional feeding groups” (kz. FFG) Klassifizierung wurde von Cummins (1973) und
Kollaboratoren (CUMMINS & KLUG 1979) eingeführt, um die Einbeziehung von
Makroinvertebraten in gewässerökologische Studien zu erleichtern (RAMÍREZ & GUTIÉRREZ-
FONSECA 2014b). Zur Klassifizierung der verschiedenen Gruppen wurden morphologische
Charakteristiken ( z.B. Mundwerkzeuge) und Verhaltensweisen (z.B. Art der Futteraufnahme)
kombiniert betrachtet (CUMMINS & KLUG 1979), um deren Rolle in Ökosystemprozessen zu
verstehen.
Alle Taxa wurden, wenn möglich, entsprechend ihrer FFG in „scrapers“, „piercers“,
„shredders“, „collectors“, „filterers“ und „predators“ nach TOMANOVA et al. (2006), CHARÁ-
SERNA et al. (2010) und RAMÍREZ & GUTIÉRREZ-FONSECA (2014b) zugeteilt. Genaue
Beschreibungen der sechs Gruppen sind Tab. 4. zu entnehmen. Um Missverständnisse zu
vermeiden werden die englischen Bezeichnungen beibehalten; die Übersetzungen in den
Klammern dienen als Orientierungshilfe und wurden sinngemäß übersetzt. Eine laufend
aktualisierte Liste der FFG in Lateinamerika kann unter https://www.ramirezlab.net/ffg
eingesehen werden.
23Tab. 4: Beschreibung der „Functional Feeding Groups“ nach RAMÍREZ & GUTIÉRREZ-FONSECA (2014b).
Functional Feeding Group Beschreibung
Benutzen ihre Mundwerkzeuge, um Rohstoffe. die auf
Substraten wachsen abzuschaben. Zu diesen Rohstoffen
SCRAPERS (~WEIDEGÄNGER)
zählen u.a. Algen, die über Steine wachsen, Biofilme aus
Bakterien, Pilzen und Algen.
zerschneiden oder stechen Pflanzengewebe mit ihren
PIERCERS (~ZELLSTECHER)
Mundwerkzeugen, um an den Pflanzensaft zu kommen
Schneiden oder kauen Stücke von lebendem oder totem
Pflanzenmaterial sowie Blätter oder Holz zu kleineren
SHREDDERS (~ZERKLEINERER) Stücken, welche flussabwärts transportiert werden. Dabei
werden auch Nährstoffe für Mikroben frei. Sie werden
auch als Konsumenten des CPOM bezeichnet.
COLLECTOR-GATHERERS Haben modifizierte Mundwerkzeuge, um kleine Partikel
(~SAMMLER) (FPOM) am Flussboden zu sieben oder sammeln.
Haben spezielle Anpassungen, um Partikel direkt aus dem
FILTERERS ODER COLLECTOR- Wasser zu fangen, wie z.B. das Konstruieren von Netzen
FILTERERS (~FILTRIERER) (einige Familien der Trichoptera) oder modifizierte
Mundwerkzeuge (Bsp. Simuliidae).
Fangen andere Organismen als Nahrung. Dabei gibt es
PREDATORS (~RÄUBER) verschiedene Strategien, um Beute zu fangen, u.a.
modifizierte Mundwerkzeuge (Bsp. Libellen)
243.8. Datenanalyse
Unterschiede in der Gesamtabundanz, der taxonomischen Vielfalt und dem Beitrag von
Schlüsselgruppen wie Eintagsfliegen (Ephemeroptera), Köcherfliegen (Trichoptera) und
Steinfliegen (Plecoptera) oder „Functional Feeding Groups“ (FFG) in der benthischen
Gemeinschaft zwischen Referenz- und beeinträchtigten Standorten wurden mit Hilfe von t-
Tests (unter Verwendung des Bootstrap-Verfahren) zusammen mit Nullhypothesentests (p-
Werte für Differenzen) untersucht. Mittelwerte, Standardabweichungen, Differenzen mit 95%
Konfidenzintervall (CI) und Signifikanzniveaus wurden aus 9999 bootstrapped Proben mit
9999 Permutationen errechnet. Um die Verteilung der FFGs an den Standorten graphisch zu
veranschaulichen, wurde eine Hauptkomponentenanalyse (engl. PCA) durchgeführt.
Um die Unterschiede in der Makrozoobenthos Gemeinschaftsstruktur zwischen Wald- und
beeinträchtigten Standorten zu quantifizieren, wurden eine multivariate, nicht- parametrische
Varianzanalyse (einfaktorielle PERMANOVA) angewendet, die Signifikanz durch
Permutation beurteilt. Bei signifikanten Ergebnissen der PERMANOVA mit 9999
Permutationen, wurden paarweise Tests zwischen den einzelnen Standorten durchgeführt. Eine
nicht-metrische multidimensionale Skalierung (NMDS) verbildlicht die Unterschiede zwischen
den Standorten; umso größer die Distanz zwischen den Probenahmen, desto größer der
Unterschied in der Makroinvertebraten- Lebensgemeinschaft. Schließlich gibt die
„Ähnlichkeitsprozentsatzanalyse“ (SIMPER) Aufschlüsse darüber, welche taxonomischen
Gruppen die Unterschiede in der Makroinvertebraten-Struktur zwischen Wald- und
anthropogenen Standorten ausmachen. Alle drei Verfahren basieren auf dem Bray-Curtis
Ähnlichkeitsindex der Gemeinschaftsstruktur (Taxa x Abundanz).
Alle statistischen Analysen wurden mit Hilfe von PASTv3.25 (HAMMER et al. 2001)
durchgeführt; PCA- und NMDS-Grafiken wurden mit der Versuchsversion von Canoco5
(SMILAUER & LEPŠ 2014) erstellt.
254. Ergebnisse
4.1. Physikalisch-chemische Parameter
Für die hydrochemische Analyse wurden Temperatur [°C], Konduktivität [µS/cm],
Sauerstoffkonzentration [mg/L] und -sättigung [%] und pH-Wert jedes Flusses gemessen. Eine
Auflistung aller Werte befindet sich in Tab. 5. Unterschiede zwischen Temperatur,
Konduktivität und Sauerstoffsättigung zwischen Wald(W)- und beeinflussten (A) Standorten
sind ersichtlich. Die Temperatur an Waldstandorten war deutlich geringer als an den
anthropogenen Standorten. Die geringste Temperatur mit 24,3°C wurde am Referenzort des
QG verzeichnet, die höchste im beeinträchtigen Gebiet des QB. Der geringste Unterschied
(1,4°C) zwischen den Standorten bestand am QG, der größte am QB (2,49°C). Die
Konduktivität war an den Referenzstandorten prinzipiell höher als an deren zugehörigen
Standorten flussabwärts. So lagen die Unterschiede bei 19,1µS/cm (QN), 51,5µS/cm (QG) und
25,14µS/cm (QB). Sauerstoffkonzentration und –sättigung konnten nur an QN problemlos
gemessen werden; Werte für QB liegen aus der zoologischen Exkursion der Universität
Innsbruck vor; für QG konnten keine Werte gemessen werden. Die Sauerstoffsättigung war an
den anthropogen beeinflussten Standorten höher als an den Waldstandorten und unterschied
sich um 12% bei QN und um 27,40% bei QB. Der pH-Wert lässt keine Rückschlüsse auf
Unterschiede zwischen den Standorten schließen und befindet sich zwischen 7,61(QN-A) und
8,05 (QB-W).
Tab. 5: Physikalisch- chemische Parameter der sechs Beprobungsstandorte. Mit * gekennzeichnete Werte entstammen der
Exkursion im Februar (siehe 3.2); nicht vorhandene Daten wurden mit n.a. (not available) gekennzeichnet.
Abk.: QN Quebrada Negra QG Quebrada Gamba, QB Quebrada Bolsa; W Wald- und A anthropogener Standort.
Fluss QN-W QN-A QG-W QG-A QB-W QB-A
Datum 11.12.18 15.12.18 23.01.19 24.01.19 25.12.18 27.12.18
Temperatur [°C] 24,80 26,02 24,30 25,70 24,58 27,07
Konduktivität [µS/cm] 210,20 191,20 236,50 185 186,37 161,23
O2 [mg/L] 7,14 7,90 n.a. n.a. 7,91* 9.06*
O2 [%] 89,30 101,30 n.a. n.a. 95,20* 122,60*
pH- Wert 7,63 7,61 7,82 7,62 8,05 7,58
264.2. Abundanz und Diversität Insgesamt wurden 23838 Lebewesen beim quantitativen Beproben gesammelt, 6694 an den Referenzstandorten und 17144 an den beeinflussten Standorten. Alleine an dem Weidestandort am Quebrada Negra (QN) wurde fast die Hälfte der Individuen (9209) gesammelt, davon gehören alleine 7594 Individuen der Familie Chironomidae (Zuckmücken) an. Die zweithöchste Individuenzahl pro Standort wurde am unteren Standort des Quebrada Bolsa (QB) mit insgesamt 5290 Individuen verzeichnet, wobei alleine 1475 Individuen den Oligochaeta und 1320 den Chironomidae angehören. Es folgt der anthropogen beeinflusste Standort des Quebrada Gamba (QG) mit 2645 Individuen. Bei den Waldstandorten ist der Unterschied in der Individuenzahl weitaus niedriger: mit 2564 liegt QB an erster Stelle, gefolgt von QG mit nur 68 Individuen weniger (2496); QN hat deutlich am wenigsten Individuen mit nur 1627. Wenn man die beiden Standorttypen vergleicht, sind benthische Lebensgemeinschaften in anthropogen beeinflussten Standorten deutlich abundanter als in Referenzgegenden. Statistisch signifikant (p
Die taxonomische Vielfalt reichte von 41 Taxa am beeinflussten Standort des QB bis zu 50
Taxa am Waldstandort desselben Flusses. QN hat 48 Taxa am Waldstandort und 45 am
Weidestandort und QG 43 am Waldstandort und 46 am Standort flussabwärts. Eine
Gesamttaxaliste, die alle Beprobungsorte umfasst, ist im Anhang (Tab. 9) zu finden. Insgesamt
wurden 92 Makroinvertebraten- Taxa in den Untersuchungsflüssen identifiziert, wobei 44
Fluginsektenfamilien (Pterygota) und zehn andere taxonomische Gruppen (Hydrachnidia,
Oligochaeta, Ostracoda, Copepoda, Platyhelminthes, Nematomorpha, Collembola, Decapoda,
Bivalvia und Hirudinea) vorhanden sind.
Bei den qualitativen Proben von QN und QB wurden insgesamt 412 Individuen gesammelt, 296
von QB (141 Waldstandort und 155 beeinflusster Standort) und 116 von QN (65 Waldstandort
und 51 beeinflusster Standort). Wie auch bei den quantitativen Proben ist die Anzahl der
Familien bei den Waldstandorten deutlich höher als bei den anthropogen Beeinflussten. So
wurden an den oberen Abschnitten des QN und QB 16 bzw. 14 und an den unteren Abschnitten
8 bzw. 11 Familien gezählt. Bemerkenswert ist, dass am Referenzort des QN trotz geringer
Individuenzahl mehr Familien als am Referenzort des QB vorkommen.
4.3. Zusammensetzung der benthischen Lebensgemeinschaften
Aquatische Insekten waren die dominierende Gruppe in den drei Flüssen. Sogenannte Nicht-
Insekten Taxa machten an Waldstandorten nur 2,76% aus; an beeinflussten Standorten 8,79%,
wobei davon ca. 90% aller Individuen (1475) der Gruppe der Oligochaeta des beeinflussten
Standorts des QB angehörten. Insgesamt waren mehr als 27% aller Individuen an diesem
Standort Oligochaeta und somit die am zweitstärksten vertretene Gruppe nach den
Chironomidae mit 33%. Diptera war die häufigste Ordnung in allen Gebieten (~65% aller
Individuen) mit der zahlenmäßig größten Familie der Chironomidae (~58% aller Individuen).
Am Weidestandort des QN waren knapp 84% aller Makroinvertebraten Zuckmücken
(Chironomidae). Neben den Diptera waren Ephemeroptera, Oligochaeta, Trichoptera und
Coleoptera die häufigsten Taxa. Abb. 14 zeigt die absolute und relative Häufigkeit der
Großgruppen an allen sechs Standorten:
2810000
Ephemeroptera Trichoptera
9000
8000
Diptera Coleoptera
7000
Anzahl der Individuen
6000
other insect taxa Non insect taxa
5000
4000
3000
2000
1000
0
Wald Weide Wald Straße Wald Siedlung
Quebrada Negra Quebrada Gamba Quebrada Bolsa
100%
90%
80%
70%
60%
50%
40%
30%
20%
10%
0%
Wald Weide Wald Straße Wald Siedlung
Quebrada Negra Quebrada Gamba Quebrada Bolsa
Abb. 14: Taxonomische Zusammensetzung des Makrozoobenthos in Wald- und beeinflussten Standorten gemessen an der
absoluten Häufigkeit (oben) und der relativen Häufigkeit (unten). Vgl. QN und QB in DUSCHEK et al. (2019) .
29Ein “One-Way PERMANOVA” zeigte, dass benthische Lebensgemeinschaften sich maßgeblich zwischen Waldstandort und anthropogenem Standort unterscheiden, und dass das Vorhandensein bzw. Fehlen von Ufervegetation mit der Zusammensetzung der benthischen Gemeinschaft zusammenhängt (p
Die Simper-Analyse gibt Aufschlüsse, welche taxonomischen Gruppen die Unterschiede
zwischen Wald- und anthropogen beeinflussten Standorten erklären. Der Unterschied zwischen
den Standorttypen konnte zu 82,96% von den elf häufigsten Taxa erklärt werden. Den größten
Beitrag leistete die Ordnung Diptera (3 Taxa, 46.69%), gefolgt von Ephemeroptera (5 Taxa,
20.12%), Oligochaeta (1 Taxon, 11.34%), Trichoptera (1 Taxon, 3,32%) und Coleoptera (1
Taxon, 1,46%). Die Unähnlichkeit (average dissimilarity) der Makroinvertebraten-
Gesellschaften war bei QN am größten mit mehr als 70%, bei QB lag sie bei ungefähr 66% und
bei QG bei knapp unter 50%. Meist haben dieselben Taxa zu Unterschieden zwischen den
Standorttypen an den drei Flüssen beigetragen. Sieben der 15 Taxa der Ordnungen Diptera
(Chironomidae, Ceratopogoniae und Simuliidae), Ephemeroptera (Baetodes, Tricorythodes
und Leptohyphidae) und Trichoptera (Leptonema) tragen in allen drei Flüssen zu Unterschieden
zwischen Wald- und anthropogenen Standorten bei. Einige Unterschiede zwischen den
Standorten waren dennoch offensichtlich. Bei QN und QG trägt die Familie der Chironomidae
(QN: 77,28%, QG: 21,62%) den größten Unterschied in der Zusammensetzung der
Lebensgemeinschaften bei, bei QB die Gruppe der Oligochaeta (29%), die bei QN und QG
keinen relevanten Beitrag leistet. Die Gattung Baetodes ist bei QG und QB an Waldstandorten
deutlich häufiger, bei QN kommt sie am Weidestandort öfter vor. 80% der Ungleichheit
(dissimilarity) wird bei QN von den ersten drei Taxa ausgelöst, währenddessen es bei QB die
ersten sieben sind und bei QG 13 der 15 gelisteten Taxa, die 80% der Ungleichheit ausmachen.
Die Reihung der ersten 15 Taxa der Simper-Analyse können Tab. 6 entnommen werden.
31Tab. 6: Liste der Taxa, die die größten Beiträge zur Unähnlichkeit zwischen Wald- und anthropogenen Standorten leisten,
basierend auf der SIMPER - Analyse, in der Reihenfolge abnehmender Bedeutung. Fettgedruckte Taxa kamen häufiger an
Waldstandorten vor. Die durchschnittliche Unähnlichkeit (Average dissimilarity) in Klammer.
Quebrada Negra Quebrada Gamba Quebrada Bolsa Gesamt
(70,87%) (47,12%) (65,82%) (60,57%)
1 Chironomidae Chironomidae Oligochaeta Chironomidae
2 Baetodes Simuliidae Chironomidae Oligochaeta
3 Simuliidae Leptohyphes Leptohyphes Leptohyphes
4 Elmidae larvae Farrodes Tricorythodes Tricorythodes
5 Tricorythodes Chimarra Baetodes Baetodes
6 Americabaetis Baetodes Ceratopogoniae Simuliidae
7 Thraulodes Thraulodes Leptonema Ceratopogoniae
8 Leptonema Tricorythodes Simuliidae Leptonema
9 Hydroptilidae 1 Leptonema Smicridea Farrodes
10 Leptohyphes Naucoridae Hydroptilidae 1 Thraulodes
11 Oxyethira Ceratopogoniae Hydroptila Elmidae larvae
12 Baetidae 2 Hydrachnidia Baetidae 1 Naucoridae
13 Baetidae 1 Anacroneuria Naucoridae Hydroptilidae 1
14 Ceratopogoniae Elmidae larvae Americabaetis Americabaetis
15 Hydrachnidia Dixella Farrodes Smicridea
32Sie können auch lesen

















































