Molekularmedizinische Grundlagen von fetalen und neonatalen Erkrankungen
←
→
Transkription von Seiteninhalten
Wenn Ihr Browser die Seite nicht korrekt rendert, bitte, lesen Sie den Inhalt der Seite unten
Molekulare Medizin
Molekularmedizinische Grundlagen von fetalen und neonatalen
Erkrankungen
Herausgegeben von Detlev Ganten und Klaus Ruckpaul
Bearbeitet von
Detlev Ganten, Klaus Ruckpaul, Roland R Wauer
1. Auflage 2004. Buch. XLIV, 684 S. Hardcover
ISBN 978 3 540 20138 0
Format (B x L): 19,3 x 27 cm
Weitere Fachgebiete > Medizin > Vorklinische Medizin: Grundlagenfächer
Zu Inhaltsverzeichnis
schnell und portofrei erhältlich bei
Die Online-Fachbuchhandlung beck-shop.de ist spezialisiert auf Fachbücher, insbesondere Recht, Steuern und Wirtschaft.
Im Sortiment finden Sie alle Medien (Bücher, Zeitschriften, CDs, eBooks, etc.) aller Verlage. Ergänzt wird das Programm
durch Services wie Neuerscheinungsdienst oder Zusammenstellungen von Büchern zu Sonderpreisen. Der Shop führt mehr
als 8 Millionen Produkte.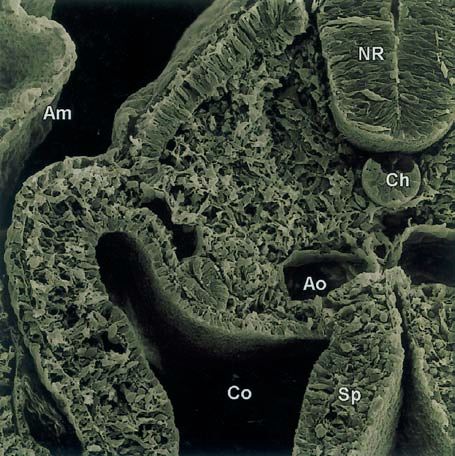
1.1 Mechanismen der Steuerung
der Embryonalentwicklung
Bodo Christ und Beate Brand-Saberi
Wer weiter nichts als die Kreaturen erkennte,
der braucht an keine Predigt zu denken,
denn jegliche Kreatur ist Gottes voll und ist ein Buch.
Meister Eckhart
Inhaltsverzeichnis
1.1.1 Geschichte des Entwicklungsbegriffs . . . 3 1.1.2.9.2 Gastrulation . . . . . . . . . . . . . . . . . . . 23
1.1.2.9.3 Regeneration . . . . . . . . . . . . . . . . . . 24
1.1.2 Grundvorgänge der Entwicklung . . . . . 7 1.1.2.9.4 Grenzziehungen . . . . . . . . . . . . . . . . 25
1.1.2.1 Wachstum . . . . . . . . . . . . . . . . . . . 7 1.1.2.9.5 Fusionen . . . . . . . . . . . . . . . . . . . . . 26
1.1.2.1.1 Zellteilung und Zellvermehrung . . . . . . 7 1.1.2.9.6 Rechts-links-Asymmetrie . . . . . . . . . . . 27
1.1.2.1.2 Zellvergrößerung . . . . . . . . . . . . . . . 9 1.1.2.10 Gefäßentwicklung . . . . . . . . . . . . . . . 29
1.1.2.1.3 Bildung von extrazellulärer Matrix (ECM) 9 1.1.2.10.1 Angiogenese . . . . . . . . . . . . . . . . . . . 29
1.1.2.2 Gene . . . . . . . . . . . . . . . . . . . . . . . 9 1.1.2.10.2 Lymphangiogenese . . . . . . . . . . . . . . . 31
1.1.2.3 Transkriptionsfaktoren . . . . . . . . . . . . 10 1.1.2.11 Entwicklung des Nervensystems . . . . . . 31
1.1.2.4 Differenzierung . . . . . . . . . . . . . . . . 11 1.1.2.11.1 Induktion des Nervensystems . . . . . . . . 31
1.1.2.5 Apoptose . . . . . . . . . . . . . . . . . . . . 14 1.1.2.11.2 Bildung des Neuralrohrs . . . . . . . . . . . 32
1.1.2.6 Extrazelluläre Matrix, Zelladhäsions- 1.1.2.11.3 Segmentierung des Gehirns . . . . . . . . . 32
moleküle und Zell-Matrix-Interaktionen . 15 1.1.2.11.4 Dorsoventrale Polarisierung
1.1.2.7 Gemeinschaftseffekt (Community Effect) 16 der Rückenmarksanlage . . . . . . . . . . . 33
1.1.2.8 Signalaustausch zwischen Zellen . . . . . . 17 1.1.2.11.5 Strukturentwicklung des ZNS . . . . . . . . 33
1.1.2.8.1 Transformierender Wachstumsfaktor . . . 17 1.1.2.11.6 Wachstum der Axone . . . . . . . . . . . . . 35
1.1.2.8.2 Fibroblastenwachstumsfaktoren . . . . . . 18 1.1.2.12 Entwicklung der Extremitäten . . . . . . . . 35
1.1.2.8.3 Epidermale Wachstumsfaktoren . . . . . . 18 1.1.2.12.1 Reziproke Interaktionen zwischen
1.1.2.8.4 Insulinähnliche Wachstumsfaktoren . . . . 18 Ektoderm und Mesoderm . . . . . . . . . . 36
1.1.2.8.5 Hedgehog-Familie . . . . . . . . . . . . . . . 19 1.1.2.12.2 Anterior-posteriore und dorsoventrale
1.1.2.8.6 WNT-Familie . . . . . . . . . . . . . . . . . . 19 Polarität . . . . . . . . . . . . . . . . . . . . . 37
1.1.2.8.7 Das Delta-Notch-System . . . . . . . . . . . 20 1.1.2.13 Entwicklung der Nieren . . . . . . . . . . . . 38
1.1.2.8.8 Die LIF-Familie . . . . . . . . . . . . . . . . 20 1.1.2.14 Die Entwicklung einer Drüse
1.1.2.8.9 Das Ephrinsystem . . . . . . . . . . . . . . . 20 am Beispiel des Pankreas . . . . . . . . . . . 39
1.1.2.8.10 Neurotrophine . . . . . . . . . . . . . . . . . 20
1.1.2.9 Morphogenetische Prozesse . . . . . . . . . 21 1.1.3 Literatur . . . . . . . . . . . . . . . . . . . . . 41
1.1.2.9.1 Morphologie und Vorkommen
von Epithelien in der Entwicklung . . . . 21
wicklung der Tiere“. Aristoteles beschreibt die Ent-
1.1.1 Geschichte des Entwicklungsbegriffs wicklung des Hühnchens im Ei. Inmitten der sich
ausbildenden Formen beobachtete er das pulsie-
Das Fragen nach der Herkunft, dem Sein, dem rende Herz und beschrieb es als den „springenden
Werden und Vergehen des Menschen hat bereits Punkt“. Die Formentwicklung (Morphogenese)
antike Philosophen beschäftigt und zu anato- wird nach Aristoteles durch ein gestaltendes Prin-
mischen und embryologischen Studien angeregt. zip „entelecheia“ vorangetrieben. Das gesamte
Die ersten ausführlichen Abhandlungen über Ent- Universum befindet sich danach in einer ständigen
wicklungsphänomene und deren Ursachen stam- Bewegung von niederen zu höheren Entwicklungs-
men von Aristoteles (384–322 v. Chr.) insbesonde- stufen. Die Vervollkommnung der Form, welche
re in seinem Werk „Von der Zeugung und Ent- die Materie prägt, geschieht nach einer Vorstel-
Ganten/Ruckpaul (Hrsg.)
gemeinsam mit R. R. Wauer
Molekularmedizinische Grundlagen
von fetalen und neonatalen Erkrankungen
© Springer-Verlag Berlin Heidelberg 2005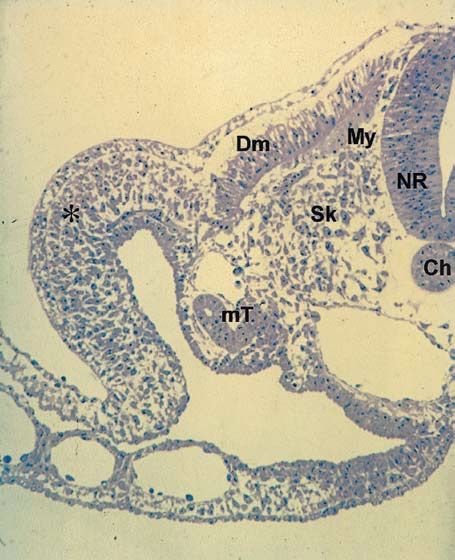
4 B. Christ und B. Brand-Saberi
lung, „eidos“, die dem wirksamen Prinzip inne-
wohnt.
Für die mittelalterlichen Menschen war die
göttliche Schöpfung der Natur und des Menschen
eher Gegenstand kontemplativer Betrachtungen.
Mit Beginn der Renaissance im 16. Jahrhundert
begann wiederum eine mehr gegenstandsbezogene
Forschung und es wurden Befunde erhoben, die
im Verlauf der folgenden Jahrhunderte in Abhän-
gigkeit von den sich ständig verbessernden Unter-
suchungsmethoden an Exaktheit zunahmen und
die unser heutiges naturwissenschaftliches Welt-
bild, d. h. unsere „Weltanschauung“, geprägt haben.
In der Embryologie stand zunächst die Lehre
von der Präformation ganz im Vordergrund. Diese
besagte, dass der Embryo von Anfang an mit allen
Teilen ausgestattet ist. Diese Teile sollten zu Be-
ginn der Entwicklung so winzig sein, dass sie
nicht identifiziert werden könnten. Die vollständig
ausgestatteten Miniaturtiere oder -menschen (Ho-
munculi) sollten entweder in den 1677 von dem
Studenten Hamm entdeckten Spermienköpfen oder
in den von de Graaf 1672 beschriebenen Eiern
(Follikeln) enthalten sein. Die beweglichen Sper-
mien wurden zunächst als Tierchen (Zoa oder
Animalcula) beschrieben und später von Karl
Ernst von Baer als Samentiere (Spermatozoa) be-
nannt. Diejenigen, die sie als Sitz der Homunculi
ansahen, wurden als Animalkulisten bezeichnet
(Abb. 1.1.1). Demgegenüber hießen diejenigen,
welche die voll ausgestatteten menschlichen Winz-
linge in den Eiern vermuteten, Ovisten. Die Präfor-
mationslehre führte die Ovisten konsequenterweise Abb. 1.1.1. Winziger Mensch in einem Spermium. Darstel-
lung nach Nicholas Hartsoeker (1656–1725)
zur Formulierung der Einschachtelungslehre (Em-
boîtement), die auf den Philosophen Malebranche
(1688) zurückgeht und besagt, dass bereits im
Ovar der Urmutter Eva ineinander verschachtelt aus den Hüllen, diese „Entkapselung“, wurde als
200 Millionen Miniaturmenschen enthalten gewe- Entwicklung oder Evolution bezeichnet. So genaue
sen seien, die alle von Gott vor 6000 Jahren an ei- Beobachter der Embryonalentwicklung des Hühn-
nem Tag geschaffen die Erde bis an das Ende aller chens und der Amphibien wie William Harvey, Al-
Tage bevölkern würden (nach Hertwig 1906). brecht von Haller, Antoni van Leeuwenhoek, Mar-
Der Streit zwischen Ovisten und Animalkulisten cello Malpighi und Jan Swammerdam, waren von
schien zugunsten der Ovisten auszugehen, als der der Richtigkeit der Präformationstheorie überzeugt
Genfer Gelehrte Charles Bonnet die Jungfernzeu- (Abb. 1.1.2).
gung (Parthenogenese) der Blattläuse entdeckte. Er Ein Bedeutungswandel des Entwicklungsbegriffs
hatte eine Blattlaus sorgfältig isoliert und beobach- wurde durch die Arbeiten von Caspar Friedrich
tete, dass sie, ohne je mit einem Männchen Kon- Wolff eingeleitet, der 1759 die Theorie der Epi-
takt gehabt zu haben, öfter hintereinander lebendi- genese entwickelte. Er ging dabei davon aus, dass
ge Junge zur Welt brachte. Nach den Vorstellungen die Entwicklung ein Fortschreiten vom Einfachen
der Ovisten und Animalkulisten waren die unend- zum Komplizierten darstellt: „Die verschiedenen
lich kleinen Miniaturbilder der später ausgewach- Teile eines tierischen Körpers entstehen alle einer
senen Geschöpfe in Hüllen eingeschlossen, die im nach dem anderen, ein jeder Teil ist also allemal
Verlauf ihres Wachstums durchbrochen und abge- erstlich ein Effekt eines anderen vorhergehenden
worfen würden. Dieser Prozess der Auswickelung Teils und als dann wiederum Ursache anderer fol-
a 1.1 Mechanismen der Steuerung der Embryonalentwicklung 5
hen werden. Von Baer war durch seinen Jugend-
freund Pander, der 1817 in den „Beiträgen zur
Entwicklung des Hühnchens im Ei“ bereits den
Übergang von der zweiblättrigen zur dreiblättrigen
Keimscheibe beschrieben hatte, für die embryolo-
gische Forschung begeistert worden. In seinem
Hauptwerk über die Embryologie der Tiere hat
von Baer 1828 am eingehendsten die Entwicklung
des Hühnchens vom Anfang der Bebrütung bis
zum Schlüpfen aus dem Ei beschrieben (Abb.
1.1.3). Dabei entdeckte er beispielsweise den Pri-
mitivstreifen. Er beobachtete, dass die Wirbeltiere
vorübergehend ein sehr ähnliches Embryonalstadi-
um durchlaufen, wobei die Embryonen höherer
Tiere jedoch nie den bleibenden Formen niederer
Tiere entsprächen: „Im Grunde ist also nie der
Embryo einer höheren Tierform einer anderen
Tierform gleich, sondern nur ihrem Embryo.“ Er
widersprach damit der insbesondere von Meckel
und später von Haeckel (1834–1919) formulierten
These, die noch heute kontrovers diskutiert wird,
dass nämlich die Individualentwicklung (Ontoge-
nese) eine abgekürzte Form der Stammesgeschich-
te (Phylogenese) darstelle. Hier zeigte sich beson-
Abb. 1.1.2. Darstellung der Entwicklung des Hühnerembryos
nach Marcello Malpighi (1628–1694)
gender Teile.“ Die späteren Organe sind demnach
nicht als solche in kleinem Maßstab zu Beginn der
Entwicklung vorhanden, sondern sie bilden sich
allmählich aus. So beschreibt Wolff in seiner
Schrift „De formatione intestinorum“, die 1768 er-
schien, wie sich der Darm des Hühnerembryos aus
einem Darmblatt über eine Darmrinne entwickelt,
deren Ränder sich einander nähern, um schließ-
lich zu einem Rohr zu verschmelzen (nach Hert-
wig 1906).
Ein weiterer wesentlicher Anstoß zum besseren
Verständnis der Embryonalentwicklung kam vom
Abt Lazzaro Spallanzani (1729–1799), der 1780 er-
folgreich künstliche Befruchtungen sowohl von
Amphibien mit Samen aus den Samenblasen der
Männchen durchführte und dem sogar die künstli-
che Befruchtung einer Hündin durch Injektion von
Samen eines Hundes in die Gebärmutter gelang.
Dadurch wurde erstmals demonstriert, dass beide
Geschlechter einen Beitrag zur Zeugung der Nach-
kommen liefern müssen (nach Hertwig 1906).
Die Eizelle selbst wurde von Karl Ernst von
Baer (1792–1876) entdeckt. Er gehörte zu den ganz
großen Forschern des 19. Jahrhunderts und kann Abb. 1.1.3. Deckblatt des Buches über die Entwicklung des
als Schöpfer der modernen Embryologie angese- Hühnchens im Ei von Karl Ernst von Baer (1828)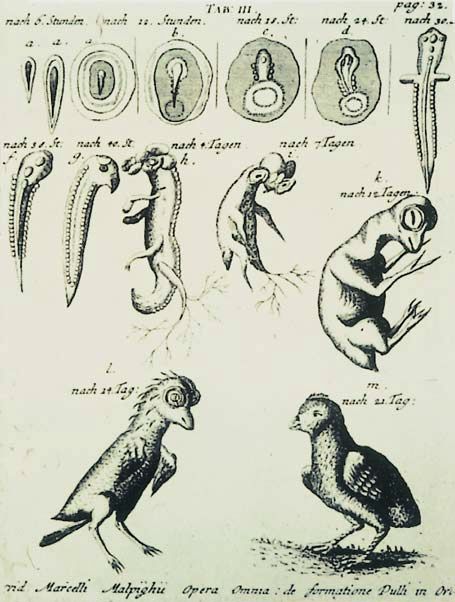
6 B. Christ und B. Brand-Saberi
ders augenfällig, wie die wachsende Erkenntnis
von naturwissenschaftlichen Zusammenhängen ei-
nen Wandel der „Weltanschauung“ bewirken kann.
Eine weitere Dimension in der Betrachtung von
Entwicklungsvorgängen wurde durch die Beobach-
tungen von Schleiden (1838) und Schwann (1839)
eröffnet, dass alle Lebewesen aus Zellen zusam-
mengesetzt seien, welche die kleinsten noch selbst-
ständig lebensfähigen Bauelemente des Organis-
mus darstellen. Hinzu kam die Erkenntnis, dass
Zellen nur durch Teilung von Zellen entstehen
können (Virchow 1855: „omnis cellulae e cellula“).
Dadurch wurde klar, dass es im Verlauf der Ent-
wicklung zu einer Spezialisierung von ur-
sprünglich gleich aussehenden Zellen kommen
muss (Differenzierung).
In der zweiten Hälfte des 19. Jahrhunderts be-
gann auf der Basis dieser Erkenntnisse und der Ver-
feinerung der mikroskopischen Untersuchungs-
methoden, deren Entwicklung insbesondere von Re-
mak, Kölliker und Hensen vorangetrieben worden
waren, die Epoche der Zellbiologie, Genetik und
der experimentellen Embryologie. Aufgrund von Er-
gebnissen mikrochirurgisch durchgeführter Defekt-
und Isolationsexperimente an Embryonen verschie-
dener Spezies begann sich die Erkenntnis durch-
zusetzen, dass embryonale Zellen bzw. Zellgruppen
für ihre Differenzierung Informationen von außen
benötigen. Driesch erkannte, dass das Schicksal ei-
Abb. 1.1.4. Originalpräparat von Spemann und Mangold mit
ner Zelle abhängig von ihrer Lage im Ganzen ist. experimentell induzierter Embryonalachse (Pfeil) nach
Demnach sind die Zellen im jungen Embryo einer Transplantation der dorsalen Urmundlippe
„Positionsinformation“ ausgesetzt, die ihr weiteres
Schicksal festlegt (determiniert). Die induktiven
Wechselwirkungen zwischen verschiedenen Keim- (1885) zur Formulierung der Keimplasmatheorie.
abschnitten wurden am Beispiel des Amphibien- Er postulierte eine besondere Substanz, das Keim-
keims von Spemann u. Mangold untersucht und plasma, das im Zellkern lokalisiert ist und den
führten zur Entdeckung eines den Embryo „organi- Träger der arteigenen Erbinformation darstellt.
sierenden“ Keimbezirks, der beim Amphibien- Dieses Keimplasma ist nach Weismann aus sehr
embryo in der oberen Urmundlippe und bei Vögeln vielen Stoffteilchen, den Determinanten, zusam-
und Säugern im Hensen-Knoten lokalisiert ist (Spe- mengesetzt, die ihrerseits aus noch kleineren Ein-
mann u. Mangold 1924, Abb. 1.1.4). Diese Fähigkeit heiten, den Biophoren, bestehen. Mit zunehmen-
multipotenter Zellen, sich entsprechend äußerer Be- den Zellteilungen sollen die Determinanten in der
fehle (Signale) zu differenzieren, wird gegenwärtig Weise auf die Tochterzellen verteilt werden, dass
in der Stammzellenforschung mit dem Ziel der Ge- am Ende in jeder Zelle nur noch eine Art von De-
webszüchtung genutzt. terminanten vorhanden ist, die den betreffenden
Aus der Beobachtung, dass sich aus einem Zelltyp spezifiziert.
Hühnerei immer nur ein Huhn und niemals ein Flemming u. Strasburger entdeckten die Chro-
Adler oder eine Taube entwickelt, kann geschlos- mosomen, die nach Boveri für die Steuerung der
sen werden, dass der Hühnchenbauplan bereits im Entwicklung verantwortlich sind. Boveri stellte
Hühnerei vorhanden sein muss. Das bedeutet, dass darüber hinaus fest, dass zwischen dem Zytoplas-
auch alle epigenetischen Prozesse während der ma und den Chromosomen Wechselwirkungen be-
Embryonalentwicklung letztendlich durch die Erb- stehen. Die Voraussetzungen für die biochemische
anlagen gesteuert werden. Diese artgemäße Steue- und molekulare Entwicklungsbiologie wurden
rung der Entwicklungsprozesse führte Weismann schließlich durch Watson u. Crick geschaffen, die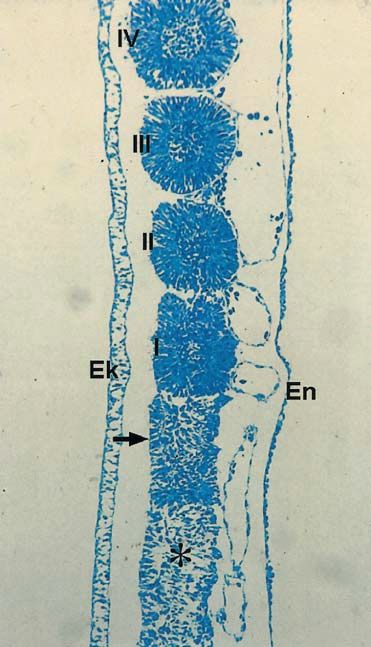
a 1.1 Mechanismen der Steuerung der Embryonalentwicklung 7
1953 die Struktur und Bedeutung der DNA als
Träger der genetischen Information aufklärten. 1.1.2 Grundvorgänge der Entwicklung
Das Wechselspiel zwischen der DNA und zytoplas-
matischen Faktoren, die Funktion einzelner Gene 1.1.2.1 Wachstum
sowie die Analyse des Austauschs und Transports
von Signalmolekülen zwischen den embryonalen Unter Wachstum verstehen wir eine Volumen- und
Zellen sind gegenwärtig Schwerpunkte der ent- Massenzunahme des Körpers. Sie kommt zustande
wicklungsbiologischen Forschung, deren Ergebnis- durch
se in den folgenden Abschnitten beispielhaft dis- • Zellvermehrung,
kutiert werden sollen. • Zellvergrößerung und
Dabei werden Befunde vorgestellt, die größten- • Bildung von Interzellularsubstanz.
teils an Modellorganismen gewonnen wurden und
zu einem besseren Verständnis der Entwicklungs- 1.1.2.1.1 Zellteilung und Zellvermehrung
prozesse bei höheren Vertebraten beitragen. Die
wichtigsten Modellorganismen für das Studium Die Zellvermehrung durch wiederholte Zellteilung
der Wirbeltierentwicklung sind der Afrikanische wird als Proliferation bezeichnet. Wenn sich die
Krallenfrosch (Xenopus laevis), der Zebrafisch Zelle teilt, durchläuft sie eine Folge von bestimm-
(Brachydanio rerio), das Hühnchen und die Maus. ten Ereignissen, die man als Zellzyklus bezeichnet.
Die Grundvorgänge der Entwicklung laufen bei In der Mitose-Phase (M-Phase) werden die dupli-
diesen Organismen grundsätzlich ähnlich ab und zierten Chromosomen auf zwei Tochterkerne ver-
sie erlauben darüber hinaus Rückschlüsse auf die teilt und es entstehen schließlich zwei Tochterzel-
Entwicklung menschlicher Embryonen. Auch die len. Der M-Phase wird die Interphase gegenüber-
molekularen Steuerungsmechanismen der Organ- gestellt, die ihrerseits aus der G1-(Gap-1-)Phase,
und Embryonalentwicklung dieser Spezies stim- der S-(Synthese-)Phase und der G2-Phase besteht.
men weitgehend überein. Nach der Zellteilung am Ende der M-Phase
Der jeweilige Modellorganismus wird in Abhän- können die Zellen entweder in die G1-Phase ein-
gigkeit von der Problemstellung und den anzuwen- treten und einen weiteren Zellzyklus durchlaufen
denden Untersuchungsmethoden ausgewählt. oder sie ziehen sich vorübergehend oder per-
Wenn im nachfolgenden Beitrag ein großer Teil manent aus dem Zellzyklus zurück, z. B. um sich
des Bildmaterials der Hühnchenentwicklung ent- zu differenzieren (Abb. 1.1.5 u. 1.1.6). Dieser post-
stammt, so werden damit keine hühnchenspezi- mitotische Ruhezustand wird als G0-Phase be-
fische, sondern allgemeingültige Entwicklungsvor- zeichnet. In der S-Phase wird die DNA verdoppelt
gänge illustriert. Der Grund für die starke Reprä- (repliziert). In der G1- und insbesondere in der
sentanz des Hühnchens liegt in dem Umstand be- G2-Phase werden Informationen von den Genen
gründet, dass die Autoren dieses Beitrags vorwie- auf mRNA umgeschrieben (transkribiert). In jun-
gend mit Hühnerembryonen gearbeitet haben und gen Embryonen können die G1- und G2-Phasen
daher auf entsprechendes Bildmaterial zurückgrei- extrem verkürzt sein, da eine Transkription noch
fen können. nicht erforderlich ist. Dadurch wird die Dauer der
Zyklen deutlich verkürzt.
Die Abfolge der Ereignisse des Zellzyklus wird
von internen Oszillatoren, den Zyklinen angetrie-
ben, die mit einem anderen kontinuierlich her-
gestellten Protein (CDC2) interagieren und den
„mitosis promoting factor“ (MPF) bilden (Draetta
1990, Kumagai u. Dunphry 1991, Murray u. Hunt
1993). MPF wird durch Kinasen und Phosphatasen
modifiziert und aktiviert. Die Steuerung der Pro-
liferation erfolgt durch Wachstumsfaktoren (z. B.
Fibroblastenwachstumsfaktor, FGF), die zum Teil
zellartspezifisch wirken, und durch andere Signal-
proteine (z. B. Sonic Hedgehog, SHH) oder WNT-
Proteine, die in der G1-Phase an die entsprechen-
den Rezeptoren der Zelloberfläche binden. Durch
Transduktion der Signale in den Zellkern erfolgt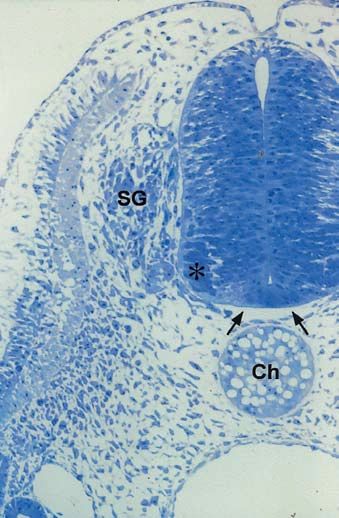
8 B. Christ und B. Brand-Saberi
Abb. 1.1.6. Transversalbruch eines 2-tägigen Hühnerembryos
im Bereich des Dermomyotoms. Beachte die hochprismati-
schen Epithelzellen und die abgerundeten Zellen, die sich in
der Mitose befinden (*). Aufnahme: Dr. H. J. Jacob, Bochum
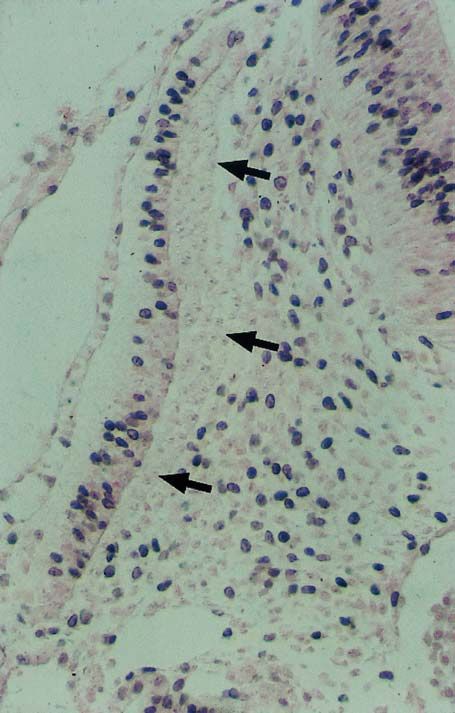
Abb. 1.1.5. Querschnitt eines 2 Tage alten Hühnerembryos
nach Applikation von 5-Brom-2'-deoxyuridin und 20-minüti-
ger Wiederbebrütung. Die dunkel gefärbten Kerne haben die
S-Phase durchlaufen. Die Zellen des Myotoms (Pfeile) sind
postmitotisch und haben sich aus dem Zellzyklus zurückge-
zogen (G0-Phase)
eine Aktivierung des Zellzyklus. Wachstumshor-
mone sind weitere Faktoren, welche die Zellprolife-
ration fördern. Die insulinähnlichen Wachstums-
faktoren 1 und 2 (IGF-1 und IGF-2) sind dafür
Beispiele (Baker et al. 1993, Heyner u. Garside Abb. 1.1.7. Transversalschnitt eines 2-tägigen Hühnerembry-
1994). Werden die Gene für IGF-1 oder IGF-2 bei os. Mittels In situ-Hybridisierung ist die Expression des
der Maus inaktiviert, so ist das Körpergewicht der Myostatin-Gens im Dermomyotom dargestellt
Neugeborenen stark reduziert (Fournier u. Lewis
2000). Andererseits kann das Körpergewicht durch
vermehrte Bildung dieser Wachstumsfaktoren be- der auch Myostatin genannt wird (Abb. 1.1.7). Die-
trächtlich erhöht werden, wobei sowohl die Zell- ser zur TGF-b-Superfamilie (transformierender
zahl erhöht ist als auch die einzelnen Zellen ver- Wachstumsfaktor b) gehörende Faktor begrenzt
größert sind (Coleman et al. 1995). Neben Signal- Zahl und Größe der Skelettmuskelfasern (McPher-
molekülen, welche die Proliferationsrate der Zellen ron et al. 1997). Mäuse mit inaktiviertem Myosta-
erhöhen, sind auch solche bekannt, welche die tin-Gen sind 30% schwerer als Wildtypmäuse, da
Proliferation hemmen. Als Beispiel sei der „growth die Einzelmuskeln der Mutanten zwei- bis dreimal
and differentiation factor“ 8 (GDF-8) angeführt, soviel Masse aufweisen wie die entsprechenden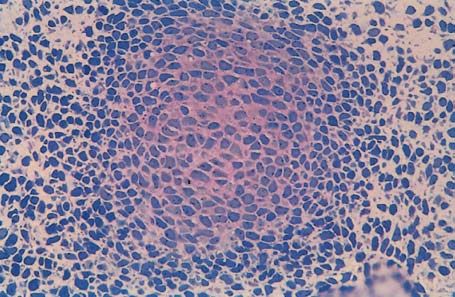
a 1.1 Mechanismen der Steuerung der Embryonalentwicklung 9
Muskeln der Normaltiere. Die Blockade der
Myostatin-Wirkung durch die Applikation von An-
tikörpern führt zu einer Vermehrung der Muskel-
masse und einer Verbesserung der Muskelfunktion
bei der mdx-Maus, die eine Muskeldystrophie ent-
wickelt (Bogdanovich et al. 2002).
1.1.2.1.2 Zellvergrößerung
Einige Gewebe bzw. Organe wachsen insbesondere
in der Fetalzeit (Entwicklungsperiode vom Beginn
des 3. Monats bis zur Geburt) sowie nach der Ge-
burt durch Größenzunahme der Zellen. Als Bei-
Abb. 1.1.8. Knorpeldifferenzierung in der Extremitätenanla-
spiel sei das zentrale Nervensystem angeführt, das ge eines Hühnchens. Beachte die rötliche knorpelspezifische
durch Vermehrung und Wachstum der Zellfortsät- extrazelluläre Matrix zwischen den noch eng beieinander
ze, deren Ummantelung mit Myelin sowie durch liegenden Knorpelbildungszellen
Volumenzunahme der Perikarya der Nervenzellen
an Masse zunimmt. Ein weiteres Beispiel für ex-
tensives Zellwachstum stellt die Skelettmuskulatur
dar. Durch die Einlagerung von Strukturproteinen
in die Muskelfasern, die für die Kontraktilität der
Fasern von Bedeutung sind, erfolgt deren Volu-
menzunahme. IGF-1 und IGF-2 wirken auch auf
das Wachstum von Muskelfasern stimulierend
(Coleman et al. 1995). Interessanterweise werden
beide insulinähnlichen Wachstumsfaktoren auch
von den Muskelzellen selbst gebildet, sodass sie
nicht nur systemisch, sondern auch auto- oder pa-
rakrin wirken dürften. Auf die das Wachstum der
Muskelzellen hemmende Wirkung von Myostatin
wurde bereits hingewiesen.
1.1.2.1.3 Bildung von extrazellulärer Matrix (ECM)
Die Bildung von Zwischenzellensubstanz, die auch
extrazelluläre Matrix genannt und im angloame-
rikanischen Schrifttum ECM abgekürzt wird, be-
ginnt mit der Transformation von Epithelien in
Abb. 1.1.9. Kollagenes Fibrillennetz der extrazellulären Ma-
embryonales Bindegewebe (Mesenchym). Die ECM trix (ECM) im embryonalen Bindegewebe. * Fibroblast
ist gewebsspezifisch zusammengesetzt und wird
von lokalen Mesenchymzellen sezerniert. Das Men-
genverhältnis von Zellen zur ECM wechselt in Ab- 1.1.2.2 Gene
hängigkeit vom Gewebstyp und verändert sich
auch während der Entwicklung (Abb. 1.1.8 u. Ein Gen ist ein DNA-Abschnitt, der Funktionen im
1.1.9). Besonders deutlich ist das durch starke Leben eines Organismus hat. Die meisten Gene
ECM-Produktion verursachte Wachstum des hyali- von eukaryonten Zellen sind in den Chromosomen
nen Knorpels, bei dem der Volumenanteil der der Kerne lokalisiert. Einige Gene lassen sich in
ECM den der Zellsubstanz weit übertrifft. Die Ver- der DNA der Mitochondrien nachweisen. Entwick-
mehrung der ECM im hyalinen Knorpel ist die Ur- lungskontrollgene sind solche Gene, die bei der
sache seines von innen heraus erfolgenden (inter- Festlegung und Steuerung des Körperbauplans so-
stitiellen) Wachstums (Benninghoff u. Drenckhahn wie bei der Differenzierung der Gewebe wichtige
2003). Funktionen haben. Bei der Expression eines Gens
wird der dem Gen entsprechende Abschnitt der
DNA transkribiert, d. h. in RNA übersetzt. Bei den
10 B. Christ und B. Brand-Saberi
meisten Genen wird diese RNA auch in Proteine über einen langen Zeitraum der Evolution erhalten
translatiert und nur bei wenigen Klassen von Ge- geblieben sind. Die 183 Basenpaare der Homöo-
nen (z. B. rRNA-Genen) erfüllt die transkribierte box-Gene kodieren ein aus 61 Aminosäuren beste-
RNA eine biologische Funktion und wird nicht in hendes Proteinsegment, dass die Eigenschaft be-
ein Protein translatiert. In der Entwicklung spielt sitzt, spezifisch an die DNA zu binden und damit
die unterschiedliche (differenzielle) Genexpression die Expression anderer Gene zu steuern. Dieses
eine große Rolle, da in allen Zellen die für die dif- Proteinsegment wird als Homöodomäne oder He-
ferenzierten adulten Zellen typischen Expressions- lix-Turn-Helix-Motiv bezeichnet. Die Expressions-
muster irgendwann einmal angeschaltet werden domänen von Homöobox-Genen sind häufig
müssen. Die Kontrolle der Genexpression ist ab- räumlich auf bestimmte Strukturen begrenzt, für
hängig von regulatorischen Sequenzen der RNA, deren Entwicklung sie zuständig sind. Oftmals
den Enhancer- und Promotorregionen und von wirken mehrere Homöobox-Gene einer Steue-
Proteinen, die Transkriptionsfaktoren genannt wer- rungsebene in kombinatorischer Weise zusammen.
den, und die mit diesen DNA-Sequenzen interagie- So wird die regionale Gliederung der Wirbelsäule
ren und die Transkription der Gene entweder und der Extremitäten durch spezifische Expressi-
hemmen oder aktivieren. onsmuster von Homöobox-Genen gesteuert. Ande-
Zum Nachweis, dass ein bestimmtes Gen zu ei- re Gruppen von Entwicklungskontrollgenen sind
nem bestimmten Zeitpunkt an einem bestimmten die pax-Gene oder die Gene, die für die myogenen
Ort exprimiert wird, kann man das translatierte Determinationsfaktoren (MYF-5, MyoD, MRF4 und
Protein z. B. durch spezifische Antikörper nachwei- Myogenin) kodieren (Abb. 1.1.10). Letztere gehö-
sen. Häufiger jedoch wird die transkribierte RNA ren zu den basischen Helix-Loop-Helix-(bHLH-)
mittels In situ-Hybridisierung nachgewiesen. Um Proteinen. Weitere Transkriptionsfaktoren sind die
den Effekt eines Gens auf die Entwicklung zu stu- Zinkfinger-Proteine und die T-Box-Faktoren.
dieren, hat sich die Methode der Ausschaltung
spezifischer Gene in sog. Knock out-Mäusen be-
währt. Dafür wird ein DNA-Molekül (Vektor) her-
gestellt, das mit den Sequenzen in dem aus-
zuschaltenden Gen homolog ist. Durch homologe
Rekombination wird dieser Vektor in das Gen ein-
gebaut, das dadurch in seiner Nukleinsäuresequenz
so verändert wird, dass es nicht mehr ordnungs-
gemäß transkribiert werden kann. Derartige ge-
zielte Mutagenesen werden an embryonalen
Stammzellen der Maus durchgeführt, die in Blasto-
zysten implantiert werden.
1.1.2.3 Transkriptionsfaktoren
Transkriptionsfaktoren sind die Proteine, welche
die Transkription regulieren. Den Transkriptions-
faktoren ist gemeinsam, dass sie sich an spezi-
fische Stellen der DNA anlagern und dadurch die
Transkription bestimmter Gene beeinflussen
können. Gene, die in der Entwicklung für Tran-
skriptionsfaktoren kodieren, werden als Entwick-
lungskontrollgene bezeichnet. Eine besonders
wichtige Gruppe von Transkriptionsfaktoren sind
die Produkte der Homöobox-(hox-)Gene. Der Na-
me Homöobox bezeichnet einen hochgradig kon-
servierten DNA-Abschnitt von 183 Basispaaren.
Abb. 1.1.10. Expression des myod-Gens bei einem Hühner-
„Konserviert“ heißt in diesem Zusammenhang, embryo während des 5. Bebrütungstages. Beachte die seg-
dass identische DNA-Sequenzen bei vielen ver- mentalen Muskelanlagen in der Rumpfwand und die Vor-
schiedenen Lebewesen zu finden und sie somit muskelmassen in den Extremitätenknospen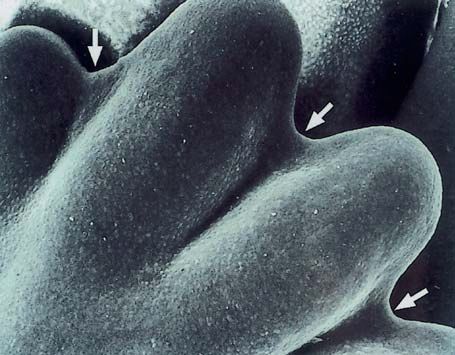
a 1.1 Mechanismen der Steuerung der Embryonalentwicklung 11
1.1.2.4 Differenzierung ert. Bei jedem Schritt des Differenzierungsprozes-
ses verfügt die Zelle jeweils über nur wenige Op-
tionen. Die von außen einwirkende Information ist
In vielen Geweben geht die Proliferation der Zellen daher weniger instruktiv als vielmehr permissiv.
ihrer Differenzierung voraus. Die Balance zwischen So kann sich ein Myoblast unter normalen Bedin-
diesen beiden Prozessen bestimmt die artspezi- gungen nicht zu einer Nervenzelle differenzieren.
fische Menge von Zellen in den sich entwickelnden Welches sind nun die Mechanismen, die zur zell-
Geweben und Organen (Raff 1996, Christ et al. typspezifischen Expression von Genen führen?
2001, Patel et al. 2002). In den meisten Geweben Hierbei ist die Regulation der Transkription eines
bleiben in ihrem Schicksal zwar festgelegte (deter- Gens von entscheidender Bedeutung. Das soll am
minierte), jedoch noch teilungsfähige Zellen erhal- Beispiel der Differenzierung der Skelettmuskulatur
ten, die als Stammzellenersatz für physiologischer- (Myogenese) dargestellt werden. Zellen, die Ske-
weise absterbende Zellen oder für Reparaturpro- lettmuskulatur bilden, entstammen einem einzigen
zesse zur Verfügung stehen, wie z. B. die Satelliten- Mesodermkompartiment, das neben den Axial-
zellen der Skelettmuskulatur. Die Zelldifferenzie- organen, Neuralrohr und Chorda dorsalis, gelegen
rung führt zu unterschiedlichen Zelltypen, wie ist, dem paraxialen Mesoderm (Christ et al. 1977,
Nerven-, Muskel-, Fett-, und Knorpelzellen, die Christ u. Ordahl 1995) (Abb. 1.1.11, 1.1.12, 1.1.13,
sich in ihrer Morphologie, Proteinausstattung und 1.1.14). In ihm entstehen segmental angeordnete
Funktion unterscheiden. In höheren Vertebraten Somiten, die in dorsoventraler Richtung in zwei
können mehr als 200 differenzierte Zelltypen ein- Kompartimente untergliedert werden, das dorsal
deutig voneinander unterschieden werden. Der gelegene epitheliale Dermomyotom und das ventral
Zelldifferenzierung liegt eine kontinuierlich erfol- gelegene mesenchymale Sklerotom (Abb. 1.1.15).
gende Veränderung der Genexpression zugrunde, Die teilungsfähigen Zellen des Dermomyotoms, die
die schließlich zur Bildung der zelltypischen Pro- z. B. das pax3-Gen exprimieren, das für einen
teine führt. Die gewebsspezifische Programmie- Transkriptionsfaktor kodiert, haben die Option,
rung (Spezifizierung) der Zellen läuft in der Regel sich zu Muskel-, Dermis- und Endothelzellen zu
schrittweise ab und erstreckt sich über mehrere differenzieren (Huang et al. 2003) (Abb. 1.1.16).
Zellgenerationen. In den frühen Phasen erfolgen Unter dem Einfluss von Signalmolekülen, die von
offenbar Aktivitätsänderungen nur einiger weniger Zellen der Axialorgane abgegeben werden, erfolgt
Gene, die Zellen werden jedoch hinsichtlich ihrer die Determination der medial im Dermomyotom
Differenzierungspotenzen immer stärker einge- lokalisierten Zellen in Richtung Myogenese. Zu
schränkt (Restriktion der prospektiven Potenz). den determinierenden Signalmolekülen gehören
Wenn sie sich nur noch zu einem einzigen Zelltyp das von der Chorda dorsalis sezernierte Sonic
entwickeln können, werden sie als „determiniert“ Hedgehog (SHH) (Abb. 1.1.17 u. 1.1.18) und die
bezeichnet. Differenziert ist die Zelle, wenn sie vom dorsalen Neuralrohr gebildeten WNT-Prote-
aufgrund der gewebs- oder organtypischen Merk- ine, WNT-1 und WNT-3a (Münsterberg u. Lassar
male identifiziert werden kann. Die noch teilungs- 1995, Fan et al. 1997, Fan u. Tessier-Lavigne 1994).
fähige determinierte Zelle vererbt ihr Differenzie- Diese Signalmoleküle binden spezifisch an Rezep-
rungsprogramm auf ihre Tochterzellen. Es wird so- toren, die von den Muskelvorläuferzellen im Der-
mit ein Zellgedächtnis etabliert, das gewebs- bzw. momyotom exprimiert werden. Über ein komple-
organtypische Regenerationen oder Reparaturen xes Signaltransduktionssystem werden myogene
ermöglicht. So gehen beispielsweise bei einer Mus- Steuerungsgene, wie myod („myoblast determining
kelschädigung aus den Stammzellen, die mit den genes“), angeschaltet, die Meistergene der Muskel-
Muskelfasern assoziiert sind, den Satellitenzellen, differenzierung darstellen und untergeordnete Ef-
immer Muskelzellen und nie Nervenzellen hervor. fektorgene aktivieren (Abb. 1.1.10). Man kennt vier
Einmal installierte Differenzierungsprogramme Schlüsselgene der Myogenese, die für Proteine ko-
werden normalerweise zeitlebens beibehalten. Än- dieren, die auch als MDF („muscle determination
derungen dieses Programms, die als Transdifferen- factors“) bezeichnet werden: myod, myf5, mrf4
zierung bezeichnet werden, stellen normalerweise und Myogenin (Übersicht bei Arnold u. Braun
selten vorkommende Ereignisse dar, bei der 2000). Es handelt sich um Proteine mit einer basi-
Züchtung und Spezifizierung von Stammzellen schen Helix-Loop-Helix-(bHLH-)Domäne, die sich
sind sie dagegen von großer Bedeutung. an eine Steuerregion der nachgeschalteten muskel-
Die einzelnen Schritte der Zelldifferenzierung spezifischen Gene (E-Box des Promoters) heftet
werden von zahlreichen äußeren Signalen gesteu- und diese aktiviert. Die myogenen Determinati-12 B. Christ und B. Brand-Saberi
Abb. 1.1.12. Zwei Tage alter Hühnerembryo mit Darstellung
der Expression des Paraxis-Gens. Paraxis markiert das Kom-
partiment des paraxialen Mesoderms
und Myogenin aktivieren und muss demnach „up-
stream“ dieser MDF-Gene wirken. Die MDF intera-
gieren mit einem zweiten Typ von myogenen Re-
Abb. 1.1.11. Rasterelektronenmikroskopische Dorsalansicht
eines 2 Tage alten Hühnerembryos. Auf der rechten Seite ist gulationsgenen den mef („myocyte enhancing
das Oberflächenektoderm zur Darstellung der darunter gele- factors“). mef2 verstärkt und stabilisiert die Ex-
genen Somiten entfernt (Aufnahme: Dr. H. J. Jacob, Bo- pression der MDF-Gene und trägt so zur myoge-
chum) nen Determination bei (Molkentin u. Olson 1996).
Die Aufrechterhaltung der zelltypspezifischen
Genexpression und damit des Differenzierungs-
onsfaktoren regulieren demnach die Transkription zustandes der Zellen ist an Wechselwirkungen des
als Transkriptionsfaktoren. Werden Zellen, die nor- Zellkerns mit dem Zytoplasma gebunden. Kerne
malerweise keine Muskulatur bilden, wie z. B. Fi- aus adulten Darm-, Haut- oder Nierenzellen
broblasten, in der Kultur mit dem myod-Gen können nach ihrer Injektion in entkernte Eizellen
transfiziert, so ändern sie ihr Programm in Rich- die Embryonalentwicklung in Gang bringen. Das
tung Muskeldifferenzierung (Weintraub 1993). Die wurde sowohl mit Eiern des Krallenfrosches Xeno-
MDF steuern nicht den gleichen Schritt des Diffe- pus wie auch mit Eizellen von Säugern experimen-
renzierungsprozesses, vielmehr agieren die MDF tell gezeigt (Gurdon 1986, Willmut et al. 1997,
Myogenin und mrf4 „downstream“ von myf5 und Wolf et al. 1998). Es hat sich dabei herausgestellt,
myod. Mäuse, bei denen die beiden Gene myf5 dass die Erfolgsrate höher war, wenn die Kern-
und myod inaktiviert wurden, bilden überhaupt spendenden Zellen in ihrer Entwicklung noch
keine Skelettmuskulatur aus (Rudnicki et al. 1993). nicht weit fortgeschritten waren. Diese Experimen-
Mausmutanten mit fehlender Transkription von te zeigen, dass durch Interaktionen mit dem Zyto-
pax3 und myf5 fehlt die Körpermuskulatur, wäh- plasma der Eizelle das genetische Programm von
rend die Kopfmuskulatur normal gebildet wird differenzierten Zellkernen experimentell verändert
(Tajbakhsh et al. 1998). pax3 kann myf5, myod werden kann. Wenn die Gene den Einflüssen vona 1.1 Mechanismen der Steuerung der Embryonalentwicklung 13
Abb. 1.1.15. Rasterelektronenmikroskopische Aufsicht auf ei-
nen Querbruch durch einen 3 Tage alten Hühnerembryo in
Höhe eines bereits kompartimentierten Somiten. Ek Ekto-
derm, En Entoderm, NR Neuralrohr, Ch Chorda dorsalis,
Ao Aorta, Co Coelom, So Somatopleura, Sp Splanchnopleura,
Am Amnion
Abb. 1.1.13. Sagittalschnitt (semidünn) durch einen 2 Tage
alten Hühnerembryo im Bereich der Somitenbildung. Aus
dem mesenchymalen präsomitischen Mesoderm wird gerade
ein epithelialer Somit abgegliedert (Pfeilmarkierung). * Noch
locker strukturiertes unsegmentiertes paraxiales Mesoderm.
Die Somiten sind mit römischen Zahlen durchnummeriert.
Ek Ektoderm, En Entoderm
Abb. 1.1.16. Querschnitt eines 3 Tage alten Hühnerembryos
und Darstellung der Expressionsdomänen von pax3: dorsales
Neuralrohr, Dermomyotom und in die Extremitätenanlage
auswandernde myogene Zellen
Abb. 1.1.14. Transversalschnitt durch einen 2 Tage alten
Hühnerembryo in Höhe des Somiten III. Ek Ektoderm, En
Entoderm, NR Neuralrohr, Ch Chorda dorsalis, Ao Aorta.
Aufnahme: Dr. Corina Schmidt, Freiburg14 B. Christ und B. Brand-Saberi
Abb. 1.1.19. Zehenanlagen eines 15,5 Tage alten Mausembry-
os. Im Zusammenhang mit der Separation der Fingeranla-
gen werden die „Schwimmhäute“ (Pfeile) durch Apoptose
abgebaut
Zytoplasma der Eizelle ausgesetzt werden, verhal-
ten sie sich wie die Gene im Kern einer befruchte-
ten Eizelle. Wird eine Leberzelle mit einer Muskel-
faser fusioniert, so werden im Kern der Leberzelle
die lebertypischen Gene herunterreguliert und
muskelspezifische Gene angeschaltet (Blau 1989,
Blau u. Baltimore 1991). Das ist ein weiterer Beleg
Abb. 1.1.17. Sonic-Hedgehog-Expression in der Chorda dor- dafür, dass die Inaktivierung von nicht zellspezi-
salis (Pfeil) eines Hühnerembryos zu Beginn des 2. fischen Genen in differenzierten Zellen rückgängig
Bebrütungstages gemacht werden kann.
1.1.2.5 Apoptose
Das Absterben von Zellen ist ein wesentliches Er-
eignis nicht nur im adulten Organismus, sondern
bereits im jungen Embryo (Glücksmann 1951,
Saunders 1966, Hurle et al. 1996). Dieses auf phy-
siologische Weise erfolgende und für die reguläre
Entwicklung außerordentlich wichtige Absterben
von Zellen wird als „programmierter“ Zelltod oder
Apoptose bezeichnet. Die Separation der Finger
und Zehen durch Rückbildung der Schwimmhäute
(Abb. 1.1.19) sowie die Eliminierung überschüssig
gebildeter Nervenzellen oder autoreaktiver Immun-
zellen erfolgen durch Apoptose. Untersuchungen
an Fadenwürmern (Nematoden) haben gezeigt,
Abb. 1.1.18. Querschnitt von einem 2 Tage alten Hühner- dass der programmierte Zelltod durch die Aktivie-
embryo mit Sonic-Hedgehog-Expression in der Chorda dor-
salis und der darüber liegenden Bodenplatte des Neural-
rung von zwei Genen ced3 und ced4 eingeleitet
rohrs wird (Metzstein et al. 1998). Eine Inaktivierung
dieser Gene hat das Überleben von Zellen zur Fol-
ge, die normalerweise absterben würden. Anderer-
seits hat die Inaktivierung eines weiteren Gens,
ced9, zur Folge, dass zusätzlich zu den normaler-a 1.1 Mechanismen der Steuerung der Embryonalentwicklung 15
weise absterbenden Zellen zahlreiche Zellen, die lagentypen bilden Fasern, die ihrerseits wiederum
normalerweise überleben würden, durch Apoptose fibrillär strukturiert sind. Ein Prokollagenmolekül
eliminiert werden. Wird dagegen ced9 überexpri- wird von drei untereinander gewundenen Polypep-
miert, so findet keine Apoptose mehr statt. Das tidketten, den a-Ketten, gebildet, die endständige
ced9 homologe Gen wird bei Säugern als bcl2 be- Polypeptide, Registerpeptide, aufweisen. Nach der
zeichnet. bcl2 ist demnach ein Apoptosehemmer Sekretion des Prokollagens werden diese endständi-
(Newton u. Strasser 1998). Die die Apoptose kon- gen Polypeptide im Zwischenzellraum durch Pro-
trollierenden Gene regulieren ein kaskadenartig kollagenpeptidasen abgespalten. Es entsteht auf die-
aktivierbares System von speziellen Proteasen, Ca- se Weise Tropokollagen, das nun zu größeren Ein-
spasen, welche die Apoptose über eine Fragmentie- heiten, den Fibrillen, polymerisieren kann. Typ-
rung der DNA auslösen. Morphologisch sind die I-Kollagen bildet den Hauptbestandteil der ECM.
Zellen, die bei der Apoptose zugrunde gehen, Typ-II-Kollagen findet sich in der ECM der Chorda
durch pyknotische Zellkerne charakterisiert. Die dorsalis und des hyalinen Knorpels. Typ-III-Kolla-
abgestorbenen Zellen werden schließlich von gen ist in der Grenzschicht, welche die Basallamina
Nachbarzellen phagozytiert. Es gibt Hinweise da- mit dem darunter gelegenen Mesenchymkompar-
rauf, dass die meisten Zellen suizidal vorprogram- timent verbindet, und im retikulären Bindegewebe
miert sind und von der Realisierung des Selbst- nachweisbar (Kuhn 1987). Typ-IV-Kollagen, das kei-
mordprogramms durch externe Faktoren abgehal- ne Fibrillen bildet, ist ein wesentlicher Bestandteil
ten werden müssen. So verhindert beispielsweise der Basallamina.
der Nervenwachstumsfaktor (NGF) das Absterben Eine weitere ECM-Komponente stellen die Gly-
von Neuronen des sympathischen Nervensystems kosaminglykane (GAG) dar, die größtenteils an
(Levi-Montalcini 1958, 1976) und Sonic Hedgehog Proteine gebunden sind und mit diesen Proteogly-
sichert das Überleben von Somitenzellen (Teillet et kane bilden (Esko 1991). GAG enthalten vor allem
al. 1998, Cann et al. 1999). Andererseits können Aminozucker, Uronsäure, Essigsäure und teilweise
auch Signalmoleküle, wie beispielsweise konzentra- auch noch Schwefelsäure. Zu den GAG gehören
tionsabhängig BMP-4 („bone morphogenetic pro- Hyaluronsäure, Chondroitin-4-Sulfat, Chondroitin-
tein 4“), die Apoptose induzieren (Schmidt et al. 6-Sulfat, Dermatansulfat, Heparansulfat und Kera-
1998). Die Apoptose lässt sich als besonderes Dif- tansulfat. Hyaluronsäure besitzt ein hohes Wasser-
ferenzierungsprogramm der Zellen ansehen. bindungsvermögen und ist dadurch in der Lage,
die interzellulären Räume zu erweitern und auf
diese Weise Zellwanderungen zu ermöglichen. Das
1.1.2.6 Extrazelluläre Matrix, Zelladhäsions- Aufrichten der Gaumenfortsätze während der Em-
moleküle und Zell-Matrix-Interaktionen bryonalentwicklung des Menschen kommt dadurch
zustande, dass in Folge vermehrter Produktion
Nicht alle von der Zelle gebildeten Proteine und von Hyaluronsäure und nachfolgender Hydratation
Glykoproteine bleiben innerhalb der Zelle. Ein von der extrazellulären Matrix der Gewebsturgor an-
Gewebstyp zu Gewebstyp variierender Anteil wird steigt und die Gaumenfortsätze anhebt.
von den Zellen sezerniert und füllt den Zwischen- Das Glykoprotein Fibronektin ist ein weiterer
zellraum als extrazelluläre Matrix (ECM) aus. An- Bestandteil der ECM, das insbesondere für Zell-
dere Proteine sind an der Zelloberfläche lokalisiert Matrix-Interaktionen und die Migration von Zellen
und dienen dem Zusammenhalt von Zellen oder wichtig ist, da es Verbindungen der Zellen zur ex-
deren Verbindung mit Molekülen der ECM. trazellulären Matrix vermittelt (Abb. 1.1.20). Es ist
Eine wichtige Funktion einiger ECM-Kom- ein Dimer aus zwei Untereinheiten, die durch Di-
ponenten besteht in der vorübergehenden Bin- sulfidbrücken verbunden sind, und besitzt ver-
dung, dem Transport, der Verteilung und der Prä- schiedene Domänen. Weiterhin verfügt Fibronektin
sentation von Signalmolekülen. Epitheliale Zellver- über eine Erkennungssequenz mit dem Motiv RGD
bände fußen auf einer Basallamina, die aus ge- (Arg-Gly-Asp), die mit Zelladhäsionsmolekülen
schichteten speziellen ECM-Bestandteilen besteht vom Integrintyp, die an der Zelloberfläche lokali-
und mit den angrenzenden Zellen über Kontakte siert sind, interagiert (Hynes 1992).
interagiert. Darüber hinaus muss die Basallamina Laminin ist ein großes Glykoprotein, das ins-
die Permeation von Signalmolekülen ermöglichen. besondere in der Basallamina vorkommt. Es ist,
Zu den Bestandteilen der ECM gehört Kollagen, ähnlich wie Fibronektin, für Zell-Matrix-Inter-
dessen verschiedene Typen in gewebsspezifischer aktionen von Bedeutung (von der Mark u. Good-
Weise verteilt sind (Abb. 1.1.9). Die meisten Kol- man 1993). Es besitzt Bindungsdomänen für die16 B. Christ und B. Brand-Saberi
Abb. 1.1.20. Querschnitt durch einen Hühnerembryo zu Be-
ginn des 3. Bebrütungstages mit immunhistochemischem
Nachweis der Fibronektinverteilung
Matrixmoleküle Typ-IV-Kollagen, Heparansulfat
und Entactin. Wird beispielsweise die Anheftung
von wandernden Neuralleistenzellen an Laminin
und Fibronektin durch die Blockierung der Inte-
grin-b1-Untereinheit inhibiert, so resultieren Fehl-
Abb. 1.1.21. Sagittalschnitt eines 3 Tage alten Hühnerembry-
bildungen der Neuralleistenderivate im Kopf- os mit immunhistochemischem Nachweis von N-Cadherin
bereich. Die Bedeutung von Zell-Matrix-Interaktio- im Dermomyotom und Myotom
nen konnte auch für andere wandernde Zellpopu-
lationen, wie z. B. Muskelvorläuferzellen, nach-
gewiesen werden. Die Integrine sind Glykoproteine der Zellober-
Der Zusammenhalt von Zellen wird durch Zell- fläche, die hauptsächlich mit Komponenten der
adhäsionsmoleküle bewirkt. Es werden drei Klas- ECM interagieren. Es sind Heterodimere, die aus
sen von Zelladhäsionsmolekülen unterschieden. a- und b-Untereinheiten bestehen. Innerhalb der
Die Cadherine (kalziumabhängige Adherine) sind Zelle sind sie mit Aktinfilamenten des Zytoskeletts
Transmembranproteine, die in gewebsspezifischen verbunden. Über die Integrine können Informatio-
Formen vorkommen, z. B. als E-Cadherin oder nen aus der ECM vermittelt werden, die das Zell-
N-Cadherin (Takeichi 1990, 1995) (Abb. 1.1.21). verhalten modulieren (Hynes 1992, Howe et al.
Cadherine binden Zellen in Anwesenheit von Kal- 1998).
zium. Dabei interagieren identische Moleküle be- Die Wegfindung migrierender Zellen sowie die
nachbarter Zellen (homophile Bindung). Cadherine Zell-Zell-Erkennung dürften ganz wesentlich durch
stellen die wichtigsten Adhäsionsmoleküle em- eine zelltypspezifische Adhäsivität vermittelt wer-
bryonaler Zellen dar. Der in das Zytoplasma hi- den (Brand-Saberi et al. 1996 a, b).
neinragende Schwanz der Cadherine ist über Cate-
nin mit intrazellulären Aktinbündeln verankert.
E-Cadherin, das auch als Uvomorulin bezeichnet 1.1.2.7 Gemeinschaftseffekt (Community Effect)
wird, bindet die Furchungszellen (Blastomeren)
junger Embryonen. Als Gemeinschafts- oder Community-Effekt be-
Ein anderer Typ von Zelladhäsionsmolekülen zeichnet man das Phänomen, dass zelltypische
gehört zur Immunglobulin-Superfamilie und ver- Differenzierungen häufig nur dann erfolgen, wenn
mittelt die Zell-Adhäsion kalziumunabhängig. Das die Zellen in einer Gruppe von mindestens 50–200
neuronale Zelladhäsionsmolekül N-CAM („neural Zellen beieinander liegen, wohingegen Einzelzellen
cell adhesion molecule“) zählt zu dieser Gruppe. oder kleinere Gruppen von Zellen am identischen
Es wird in der frühen Embryonalentwicklung nicht Ort keine Differenzierung zeigen. Diese Abhängig-
nur auf Nervenzellen gefunden. Auch bei diesem keit des Zellverhaltens von der Anzahl der Zellen
Molekül ist die Bindung homophil, d. h. die wurde zuerst von His (1868) für die Knorpeldiffe-
N-CAM einer Zelle binden an die N-CAM der renzierung beschrieben: „Die weiche parablasti-
Nachbarzelle (Kreis u. Vale 1999). sche Gewebsanlage muss, damit sie zu Knorpela 1.1 Mechanismen der Steuerung der Embryonalentwicklung 17
werde, in einer gewissen Reichlichkeit angehäuft 1.1.2.8.1 Transformierender Wachstumsfaktor
sein.“ Diese Auffassung wurde am Beispiel der
Knorpelentwicklung durch Isolationen und Trans- Der transformierende Wachstumsfaktor b (TGF-b)
plantationen unterschiedlicher Mengen des Anla- bildet eine große Familie interzellulärer Signalsub-
gematerials bei jungen Hühnerembryonen bestätigt stanzen (Assoian et al. 1987, Frolik et al. 1983, Ro-
(Christ 1969). In den letzten beiden Dekaden wur- berts u. Sporn 1990). Er wurde ursprünglich als
de dem Problem des Gemeinschaftseffektes von mitogener Faktor beschrieben, der von transfor-
Gurdon und Mitarbeitern besondere Aufmerksam- mierten Zellen abgegeben wird. Zu dieser Familie
keit geschenkt (Gurdon et al. 1993). Dabei konnte gehören Aktivin, Vg-1, BMP, Nodal und Myostatin
am Beispiel der Differenzierung von Muskel- und (Abb. 1.1.7 u. 1.1.22). TGF-b agieren als Dimere.
Nervengewebe gezeigt werden, dass Einzelzellen Zwei Moleküle bilden einen Komplex, der einen
oder kleinere Gruppen von Zellen ihren Determi- ebenfalls dimeren Rezeptor aktiviert. Sie wirken
nationszustand in einer fremden Umgebung nicht oft inhibitorisch auf die Zellproliferation und
beibehalten, während das bei größeren Zellgrup- fördern die Sekretion von ECM-Komponenten. Ak-
pen der Fall ist. Gurdon et al. (1993) diskutieren tivinähnliche Faktoren wie Vg-1 und Nodal sind
die Mechanismen, die für den Community-Effekt im Vertebratenembryo an der Induktion und Ver-
verantwortlich sein könnten. Wenn Zellen ein Sig- teilung von Mesoderm beteiligt (Joubin u. Stern
nalmolekül abgeben, das für den Ablauf eines Dif- 1999). Nodal ist ein Bestandteil des Signalnetz-
ferenzierungsprozesses oder die Erhaltung eines werks, der die Rechts-links-Asymmetrie determi-
Differenzierungsprogramms in einer gewissen niert (Rodriguez-Esteban et al. 2001, Schlange et
Konzentration benötigt wird, so ist leicht vorstell- al. 2002). Die BMP haben vielfältige Funktionen,
bar, dass durch eine größere Anzahl beieinander zu denen die Hemmung der neuralen Differenzie-
liegender Zellen eine höhere Konzentration des rung, die Spezifizierung des Körperbauplans, die
Faktors erreicht werden kann. Die Kopplung von Induktion von Skelettgewebe, die Aufrechterhal-
Zellen durch Kommunikationskontakte („gap tung der Proliferation der Muskelvorläuferzellen
junctions“) könnte die Funktion benachbarter Zel- und, bei höherer Konzentration, die Auslösung des
len synchronisieren und ebenfalls beim Gruppen-
effekt eine Rolle spielen. Auch die Kommunikation
über Zelladhäsionsmoleküle könnte daran beteiligt
sein.
1.1.2.8 Signalaustausch zwischen Zellen
Wie bereits ausgeführt wurde, können zwei be-
nachbarte Zellen Signale über Zelladhäsionsmole-
küle austauschen. Auch nicht benachbarte Zellen
eines Embryos kommunizieren miteinander und
beeinflussen sich gegenseitig, entweder durch die
Abgabe von Signalmolekülen, die an der Zielzelle
von spezifischen Rezeptoren gebunden werden,
oder durch das Ausstrecken langer dünner Zell-
fortsätze, der Zytonemata, die durch die ECM auch
mit weiter entfernt liegenden Zellen vorüberge-
hend in Kontakt treten. Signalmoleküle erreichen
die Zielzellen entweder durch Diffusion oder mit-
hilfe von ECM-Komponenten, die diese Moleküle
binden und den Zielzellen präsentieren. Man
nennt Signalmoleküle, die das Differenzierungsver-
halten der Zielzellen beeinflussen, auch Indukti-
onsfaktoren und den Vorgang der Zellinformation
Induktion. Bei den Signalmolekülen können ver-
Abb. 1.1.22. Aufsicht auf den kaudalen Abschnitt eines 2-tä-
schiedene Familien unterschieden werden. gigen Hühnerembryos mit Darstellung der Expression des
bmp4-Gens in der Somatopleura18 B. Christ und B. Brand-Saberi
Apoptoseprogramms gehören (Amthor et al. 1998,
1999).
Die TGF-b binden an den Typ-II-Rezeptor, der
dann mit dem benachbarten Typ-I-Rezeptor einen
Komplex bildet. Die Aktivierung des Typ-I-Rezep-
tors führt zur Phosphorylierung von SMAD-Prote-
inen im Zytoplasma, die in den Kern transportiert
werden, wo sie als Transkriptionsfaktoren Zielgene
regulieren (Attisano u. Wrana 1998).
1.1.2.8.2 Fibroblastenwachstumsfaktoren
Eine weitere wichtige Familie von Signalproteinen
stellen die FGF dar. Sie wurden in der Zellkultur
als Faktoren identifiziert, welche die Proliferation
von Fibroblasten anregen (Armelin 1973, Gospoda-
rowicz 1974, 1975). Inzwischen sind für den Säu-
ger 17 fgf-Gene (fgf1–fgf17) kloniert worden. Es
handelt sich bei den Genprodukten um Proteine
von 155–268 Aminosäuren, die alle eine konser-
vierte Sequenz von 120 Aminosäuren enthalten,
die an Heparin bzw. Heparansulfatproteoglykan
binden kann. Diese ECM-Komponenten werden
auch als FGF-Rezeptoren niedriger Affinität („low-
affinity FGFR“) bezeichnet. Der FGF-Heparansul- Abb. 1.1.23. Aufsicht auf einen 3-tägigen Hühnerembryo mit
fat-Komplex bindet an Rezeptoren mit hoher Affi- doppelter In situ-Hybridisierung für fgf8 (schwarz) und
nität („high-affinity FGFR“), welche membranstän- myod (rot). Beachte die fgf8-Expression in der ektodermalen
dige Tyrosinkinasen sind, FGFR-1–FGFR-4 (Green Randleiste der Flügelanlage (Pfeil). Aufnahme: Daniel Stolte,
et al. 1996). Diese Bindung führt zur Phosphory- Freiburg
lierung des Rezeptors und zur Aktivierung des
komplexen MAP-(„mitogen-activated protein“-)Ki- 1.1.2.8.4 Insulinähnliche Wachstumsfaktoren
nase-Transduktionsweges, an dessen Ende ERK
(„extracellular signal regulated kinase“) in den IGF-1 und IGF-2 sind Polypeptide, die den Effekt
Zellkern gelangt und Transkriptionsfaktoren durch von Wachstumshormonen verstärken (Kaye 1993).
Phosphorylierung aktiviert (Fantl et al. 1996). Das Sie werden von zahlreichen Zelltypen in engen
Wachstum der Extremitäten wird beispielsweise zeitlichen Fenstern und in spezifischer Weise ex-
über FGF gesteuert, die von Zellen der ektoderma- primiert. Im Blut und im Extrazellularraum bin-
len Randleiste AER abgegeben werden und die den die IGF an IGF-bindende Proteine („IGF-bin-
Proliferation der benachbarten mesodermalen Zel- ding proteins“, IGFBP) (Zapf et al. 1975). IGF-1
len stimulieren (Abb. 1.1.23). aktiviert den Typ-1-IGF-Rezeptor (IGFR), der auf
den meisten Zellen exprimiert wird, und bindet an
1.1.2.8.3 Epidermale Wachstumsfaktoren diesen mit hoher Affinität. IGF-2 bindet mit hoher
Affinität an den IGF-Typ-1-Rezeptor (Roth et al.
Die epidermalen Wachstumsfaktoren (EGF) stellen 1987). Eine Überexpression der IGF führt bei-
eine weitere Familie wichtiger Signalmoleküle dar spielsweise zu einer Hypertrophie und Hyperplasie
(Carpenter u. Cohen 1979). EGF, TGF-a und Neu- der Skelettmuskulatur (Adams u. McCue 1998,
roreguline sind Vertreter dieser Familie, die an Re- Awede et al. 1999). Das embryonale und postnatale
zeptoren vom Tyrosinkinasetyp (z. B. ErbB2) bin- Wachstum wird ganz wesentlich durch die IGF re-
den und über den MAP-Kinase-Signaltransdukti- guliert, deren Produktion durch komplexe hormo-
onsweg Gene aktivieren (Downward et al. 1984). nelle Regelkreise gesteuert wird. Es wird nur das
EGF sind an der Musterbildung bei Drosophila, auf dem väterlichen Chromosom lokalisierte IGF-
bei der geordnete räumliche Muster verschieden 2-Gen transkribiert, während das auf dem mütter-
differenzierter Zellen entstehen, und an der Diffe- lichen Chromosom gelegenen Gen inaktiv bleibt
renzierung des Nervensystems beteiligt. (genomische Prägung, Imprinting).Sie können auch lesen

















































