NACHWEIS UND ANALYSE BAMA-ASSOZIIERTER FALTUNGSINTERMEDIATE DES OMPA DURCH ORTSGERICHTETE FLUORESZENZSPEKTROSKOPIE - KOBRA
←
→
Transkription von Seiteninhalten
Wenn Ihr Browser die Seite nicht korrekt rendert, bitte, lesen Sie den Inhalt der Seite unten
Nachweis und Analyse BamA- assoziierter Faltungsintermediate des OmpA durch ortsgerichtete Fluoreszenzspektroskopie Dissertation zur Erlangung des akademischen Grades des Doktors der Naturwissenschaften (Dr. rer. nat.) angefertigt im Fachbereich 10 – Mathematik und Naturwissenschaften Institut für Biologie, Abteilung Biophysik Universität Kassel vorgelegt von Nicole Schürmann Kassel 8. Mai 2020
Betreuer: Prof. Dr. J. H. Kleinschmidt Promotionskommission: 1. Gutachter: Prof. Dr. J. H. Kleinschmidt 2. Gutachter: Prof. Dr. M. Maniak 3. Prüfer: Prof. Dr. R. Schaffrath 4. Prüfer: Prof. Dr. F. W. Herberg Tag der mündlichen Prüfung: 16.07.2020
Inhaltsverzeichnis Abkürzungsverzeichnis 1 Formelsymbole und Einheiten 2 1. Einleitung 4 1.1 Faltung von β-Fass Proteinen 6 1.1.1 Modellprotein für OMP-Faltungsstudien: OmpA 6 1.1.2 Faltungsmechanismus – Schrittweise Faltung und Insertion 8 1.2 Der BAM Komplex 10 1.2.1 Die Hauptkomponente des BAM Komplexes: BamA 11 1.2.2 Lipoproteine des BAM Komplexes 13 1.3 Modelle zum BamA-abhängigen Faltungsmechanismus von OMPs 15 1.4 Motivation und Ziel der Arbeit 16 2. Materialien und Methoden 19 2.1 Materialien 19 2.1.1 Bakterienstämme 19 2.1.2 Oligonukleotide 19 2.1.3 Plasmide 20 2.2 Methoden 21 2.2.1 Homologie Sequenzvergleich 21 2.2.2 Gezielte Punkt-Mutagenese 21 2.2.3 Mutagenese zur Deletion der Signalsequenz von OmpA 22 2.2.4 Expression und Reinigung von BamA und OmpA 23 2.2.5 Präparation von Lipidfilmen und großen unilamellaren Vesikeln 23 2.2.6 Faltung in LUVs und LDAO 24 2.2.7 Proteolyse mittels Trypsin 25 2.2.8 Faltungsintermediate von OmpA in Ab- und Anwesenheit von BamA 26 2.2.9 Zirkulardichroismus Spektroskopie 27 2.2.10 Fluoreszenzspektroskopie 28 3. Ergebnisse 30 3.1 Charakterisierung der Tryptophan- und Cystein-freien BamA Mutante 30 3.1.1 Identifizierung konservierter Tryptophan- und Cysteinreste in BamA-Proteinen mittels multiplem Proteinsequenzabgleich 30 3.1.2 Analyse der Faltung und Insertion der WaF, CaA-BamA Mutante 33 3.1.3 Analyse der Temperaturstabilität von wt-BamA und WaF, CaA-BamA 36 3.1.4 Nachweis der Funktionalität von WaF, CaA-BamA 38 3.2 Entwicklung von Modellmembransystemen für OmpA-BamA Interaktionsstudien 41 3.2.1 Voraussetzungen des Modellmembransystems 41 3.2.2 Zusammensetzung der LD 42 3.2.3 Temperatur und pH-Wert 44 I
3.2.4 Fluoreszenzspektroskopische Interaktionsstudie von wt-OmpA und WaF, CaA- BamA 47 3.3 Ortsspezifische Interaktionsstudien von OmpA und BamA 51 3.3.1 WaF, CaA-OmpA Faltungskinetik in Ab- und Anwesenheit von BamA 52 3.3.2 Ortsspezifische Interaktionsstudien von OmpA und BamA 55 3.3.3 Einfluss von BamA auf β-Stränge des OmpA 58 3.3.4 Einfluss von BamA auf extrazelluläre Schleifen des OmpA 62 4. Diskussion 66 4.1 Faltungsfähiges und funktionelles WaF, CaA-BamA 66 4.2 Modellmembransystem aus kurzkettigen Lipiden für dynamische BamA- Wechselwirkungsstudien mit OMPs 69 4.2.1 OmpA Faltungsintermediate erstmalig in kurzkettigen Lipiden und in Gegenwart von BamA 69 4.2.2 WaF, CaA-OmpA und ETM von OmpA zeigen stabilen Komplex mit BamA 71 4.2.3 BamA zeigt Chaperone ähnliche Funktion in wässeriger Umgebung 71 4.3 Faltungsmechanismus von OmpA in LD 72 4.4 Faltungsmechanismus von OmpA in Gegenwart von BamA 74 5. Zusammenfassung 76 6. Literatur 78 Anhang 89 Liste der Veröffentlichungen 92 Danksagung 93 Eidesstattliche Erklärung 94 II
Abkürzungsverzeichnis ATP Adenosintriphosphat BAM β-barrel assembly machinery BSA Bovines serum albumin Borax Dinatriumtetraborat-Decahydrat CD Circular dichroism, Zirkulardichroismus C-Terminus Carboxy-Terminus diC18:1PC 1-Stearoyl-2-oleoyl-sn-glycero-3-phosphocholine, DOPC DLPC 1,2-Dilauroyl-sn-glycero-3-phosphocholine, diC12:0PC DLPE 1,2-Dilauroyl-sn-glycero-3-phosphoethanolamine DLPG 1,2-Dilauroyl-sn-glycero-3-phosphoglycerol DPG Diphosphatidylglycerol E. coli Escherichia Coli EDTA Ethylendiamintetraessigsäure ETM Einzeltryptophanmutante g gefaltet HEPES 2-(4-(2-Hydroxyethyl)-1-piperazinyl)-ethansulfonsäure IM Innere Membran IMP Innenmembranproteine KTSE Kinetics of tertiary structure formation by electrophoresis LD Lipiddoppelschicht LD⋅[BamA] Lipiddoppelschicht mit gefaltetem und inseriertem BamA LDAO Lauryldimethylaminoxid LPS Lipopolysaccharid LUV Large unilamellar vesicles NaCl Natriumchlorid NRMSD Normalized root mean square deviation N-Terminus Amino-Terminus OM Outer membrane OMP Outer membrane protein OmpA Outer membrane protein A MES 2-(N-Morpholino)ethansulfonsäure MOPS 3-(N-Morpholino)propansulfonsäure 1
PC Phosphatidylcholin PCR Polymerase chain reaction PD periplasmatische Domäne PE Phosphatidylethanolamin PG Phosphatidylglycerol POTRA Polypeptide transport associated Präproteine Präkursor-Proteine SDS Sodium lauryl sulfate SS Signalsequenz TMD Transmembrandomäne TMP Transmembranproteine TPR Tetratricopeptide repeat Tris Tris(hydroxymethyl)aminomethan Trp, W Tryptophan u ungefaltet WaF, CaA Tryptophan gegen Phenylalanin (F), Cystein (C) gegen Alanin (A) substituiert wt Wildtyp Wmβn Mutanten mit W an Position m, Lokalisiert in β-Strang der Nummer n XmWLn Mutanten mit Aminosäure X an Position m getauscht gegen W, Lokalisiert in Loop (extrazelluläre Schleife) der Nummer n Formelsymbole und Einheiten Å Ångström Af Relativer Anteil der schnellen Faltungsphase c Konzentration cm Zentimeter cps Counts per second °C Grad Celsius h Stunde (hour) kDa Kilo Dalton kf Geschwindigkeitskonstante des schnellen Faltungsprozesses (fast) ks Geschwindigkeitskonstante des langsamen Faltungsprozesses (slow) L Liter 2
l Schichtdicke der Küvette Mcps Millionen counts per seconds mM Millimolar min Minute M Molar ml Milliliter Mr Relative Molarmasse n Anzahl der Aminosäuren nm Nanometer pI Isoelektrischer Punkt pKs dekadischer Logarithmus der Säurekonstante Ks rpm round per minute t Zeit XFP Anteil gefaltetes Protein Θ Elliptizität λ Wellenlänge λmax Wellenlängenposition des Fluoreszenzintensitätsmaximums Δλmax Differenz der Wellenlängenpositionen der Fluoreszenzintensitätsmaxima µM Mikro Molar 3
1. Einleitung Für das Leben essentiell sind biologische Membranen. Membranen sind Strukturelemente die die Bildung von Kompartimenten ermöglichen und sind damit Voraussetzung für die Differenzierung komplexer Organismen. So gibt es Organismen wie Gram negative Bakterien, die von zwei Membranen umgeben sind (siehe Abbildung 1). Das Cytoplasma ist von der inneren Membran (IM) umgeben. Diese ist eine Doppellipidschicht und besteht bei Escherichia coli (E. coli) aus 70-75 % Phosphatidylethanolamin (PE), 18-22 % Phosphatidylglycerol (PG) und 6-8 % Diphosphatidylglycerol (DPG) (Lugtenberg und Peters 1976, Morein et al., 1996). Die IM ist mit dem zwitterionische PE und dem anionische PG/DPG leicht negativ geladen. Inseriert in die IM sind typischerweise α-Helicale Proteine. Diese können aus einer einzelnen oder mehreren α-Helices bestehen (von Heijne und Gavel 1988). Die Insertion erfolgt mit Hilfe des SecYEG Translokasekomplexes und/oder der YidC Insertase (Pugsley 1993, Houben et al., 2000, Samuelson et al., 2000). Die Sec Translokasemaschinerie ist gleichermaßen für die Translokalisation vieler Außenmembranproteine (OMPs, outer membrane proteins) und periplasmatischer Proteine verantwortlich (siehe Cranford-Smith und Huber 2018). An die IM schließt sich das Periplasma an. Dieses Kompartiment schließt eine Vielzahl von Enzymen z.B. Proteasen und Phosphatasen und Funktionen wie Sekretion, Osmoseregulation und Protein Qualitätskontrolle ein. Weiterhin enthalten ist die sogenannte Peptidoglycanschicht bestehend aus vernetzten Zuckern und Aminosäuren (siehe Miller und Salama 2018). Umgeben ist das Periplasma von der Außenmembran (OM, outer membrane). Die OM ist im Gegensatz zur IM eine asymmetrische Membran, bei der die äußere Schicht meistens aus einer Lipopolysaccharid- (LPS) Schicht besteht, während die innere Schicht aus Phospholipiden aufgebaut ist (Kamio und Nikaido 1976, Bos et al., 2007). Bei E. coli besteht die innere zum Periplasma gerichtete Membranschicht aus 75-80 % PE, 15-20 % PG und 2-6 % DPG (Lugtenberg und Peters 1976, Morein et al., 1996). Damit zeigt sie einen leicht erhöhten PE und einen leicht erniedrigten PG Anteil gegenüber der IM. Ebenso ist der Anteil an gesättigten Lipid-Kohlenstoffketten im Vergleich zur IM minimal erhöht (Morein et al., 1996). Im Gegensatz zur IM ist an der OM, also im Periplasma, keine Energiequelle in Form von ATP verfügbar (Wülfing und Plückthun 1994). Ein geregelter Transport von Nährstoffen oder anderen Molekülen über die OM wird durch inserierte Proteine ermöglicht. Diese OMPs sind charakteristisch als β-Fass gefaltet. Bislang sind Proteine mit mindestens acht (z.B. OmpA) bis hin zu 26 (LptD) antiparallel angeordneten β-Strängen bekannt (siehe Kleinschmidt 2015). Wie alle Proteine werden auch OMPs im Cytoplasma synthetisiert und besitzen für den gezielten 4
Transport zur OM eine meist 21 Aminosäure lange N-terminale Signalsequenz. Diese wird zum Beispiel von der Sec Translokasemaschinerie erkannt und nach Translokalisation abgespalten (siehe Cranford-Smith und Huber 2018). Im Periplasma werden Präproteine (Präkursor- Proteine) von Chaperonen wie SurA oder Skp gebunden (siehe Kleinschmidt 2007, Sklar et al., 2007b). Diese verhindern eine Aggregation der hydrophoben Präproteine in der wässerigen Umgebung des Periplasmas. Von den Chaperonen werden die Präproteine zur OM transportiert. Dort müssen sie in die Membran gefaltet und inseriert werden. In Gram negativen Bakterien gibt es dafür den β-barrel assembly machinery (BAM) Komplex (Wu et al., 2005, Sklar et al., 2007a). Dieser katalysiert die Faltung und Insertion der OMPs in die OM (siehe Ricci und Silhavy 2012, Kleinschmidt 2015, Ricci und Silhavy 2019). Ähnlich zu den Insertionsmaschinerien der IM gibt es auch bei der Insertionsmaschinerie der OM Homologe in anderen Organellen wie Chloroplasten und Mitochondrien (siehe Schleiff und Soll 2005, Knowles et al., 2009, Ricci und Silhavy 2012, Tsukazaki 2019). Im Gegensatz zur IM ist in der OM bislang nur ein Komplex, der BAM Komplex, für die Insertion von OMPs bekannt. Extrazellularraum β-Fass ? BamA OM BamC Periplasma OMP BamB BamD BamE Skp SurA Peptidoglycan IMP IMP α- α- Sec Helical YidC Helical IM Cytoplasma Abbildung 1: Transport und Einbau von Transmembranproteinen bei Gram negativen Bakterien. Außenmembranproteine (OMPs) und Innenmembranproteine (IMPs) werden im Cytoplasma synthetisiert. Die Prä-IMPs werden anhand ihrer Signal-Ankersequenz von der Sec Translokasemaschinerie oder der YidC Insertase erkannt und in die innere Membran (IM) gefaltet und inseriert. Die Prä-OMPs werden anhand ihrer Signalsequenz erkannt und mit Hilfe der Sec Translokasemaschinerie über die IM translokalisiert. Im Periplasma verhindern Chaperone wie z.B. Skp und SurA die Aggregation der Prä-OMPs und transportieren sie zur Außenmembran (OM). Dort werden diese vom BAM Komplex erkannt und in die OM gefaltet und inseriert (verändert nach Kleinschmidt 2015). 5
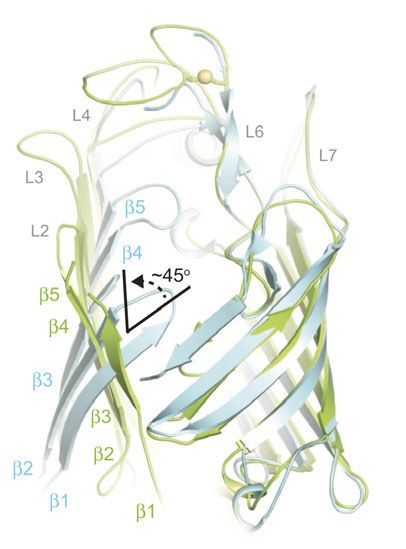
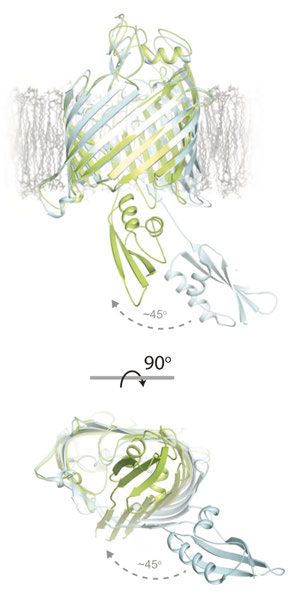
1.1 Faltung von β-Fass Proteinen In E. coli sind alle bekannten β-Fass Proteine aus einer geraden Anzahl an antiparallel angeordneten β-Strängen aufgebaut, die über lange extrazelluläre und kurze periplasmatische Schleifen verbunden sind. Das β-Fass ist durch Wasserstoffbrückenbindungen zwischen dem ersten und letzten β-Strang geschlossen. Die einzelnen Stränge zeigen eine alternierende Abfolge an hydrophoben und polaren Aminosäureresten. Hydrophobe Reste sind zur Membran hin orientiert und polare Reste in das Innere des β-Fasses (siehe Kleinschmidt 2015, Konovalova et al., 2017). Daher sind β-Fass Transmembranproteine (TMP) im Durchschnitt weniger hydrophob als α-Helicale TMP (Vogel und Jähnig 1986) und lassen sich bereits in 8 M Harnstoff entfaltet lösen und denaturieren (Schweizer et al., 1978). Helicale Proteine hingegen sind in entfalteter Form nur löslich in z.B. 88 % Ameisensäure oder Trifluoressigsäure (Huang et al., 1981). Eine Rückfaltung aus hitzedenaturiertem Zustand in LPS und Triton X-100 konnte für das β-Fass OMP OmpA (outer membrane protein A) bereits 1978 von Schweizer et al. gezeigt werden. 1.1.1 Modellprotein für OMP-Faltungsstudien: OmpA OmpA ist eines der häufigsten OMPs in E. coli (Chai und Foulds 1977) und ist ein Ionen- durchlässiges Porin, welches ebenso eine Rolle in der Strukturgebung der Zellen spielt. Weiterhin dient es als Rezeptor für viele Bakteriophagen und ist überexprimiert in E. coli Biofilmen (siehe Confer und Ayalew 2013). Es besteht aus einem achtsträngigen β-Fass und einer etwa gleich großen periplasmatischen Domäne. Die Kristallstruktur des 8-strängigen β- Fasses wurde bereits 1998 von Pautsch und Schulz veröffentlicht (Abbildung 2 A), wohingegen die etwa gleichgroße C-Terminal gelegene periplasmatische Domäne erst 16 Jahre später aufgelöst werden konnte (Abbildung 2 B) (Ishida et al., 2014). Sie besteht aus zwei langen α- Helices und vier β-Strängen die zusammen als βαβαββ-Formation erscheinen. Außerdem zeigt sich eine große Ausbuchtung in Form von zwei kleinen α-Helices, die über eine Disulfidbrücke verbunden sind. Die C-terminale Domäne bindet nicht kovalent an Peptidoglycan und stellt damit eine Verbindung zwischen der OM und dem Periplasma her (Koebnik et al., 2000, Ried et al., 1994). 6
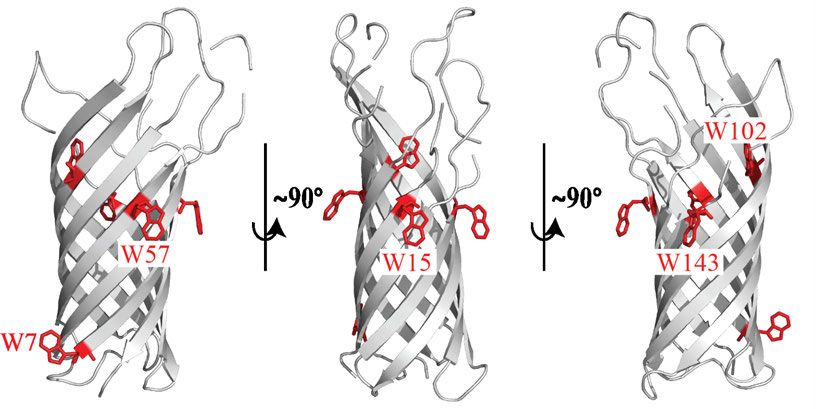
A B Abbildung 2: Kristallstrukturen der Transmembran- und periplasmatischen-Domäne von OmpA aus E. coli. (A) Die Transmembrandomäne besteht aus acht antiparallel angeordneten β-Strängen, die verbunden durch lange extrazelluläre und kurze periplasmatische Schleifen ein β-Fass ausbilden. In Rot sind die fünf nativen Tryptophanreste W7, W15, W57, W102 und W143 dargestellt (PDB Struktur: 1bxw, Pautsch und Schulz 1998). Die Verbindung der Transmembrandomäne mit der periplasmatischen Domäne (B) ist in Blau dargestellt. In Grün sind die beiden Cysteinreste dargestellt, die eine Disulfidbrücke ausbilden (PDB Struktur: 2mqe, Ishida et al., 2014). OmpA entwickelte sich über die Jahre als das Modellprotein für Faltungsstudien von OMPs. Bislang war die Rückfaltung von OmpA in über 60 Detergenzien und in Lipiden mit unterschiedlichsten Kohlenstoffkettenlängen (7-18) erfolgreich. Eine Notwendigkeit dabei ist das Überschreiten der kritischen Mizellkonzentration (Kleinschmidt, Wiener et al., 1999, Kleinschmidt 2006). Erst in Anwesenheit einer ausgebildeten Lipiddoppelschicht (LD) oder von Lipid- oder Detergensmizellen kann das OmpA β-Fass gebildet werden. Bei Rückfaltungen in LD aus einem in 8 M Harnstofflösung denaturierten Zustand konnte neben der ausgebildeten Sekundärstruktur ähnlich der nativen Sekundärstruktur ebenfalls eine orientierte Insertion von OmpA beobachtet werden (Surrey und Jähnig 1992). Orientiert bedeutet, dass die Transmembrandomäne in die Membran inseriert ist und die periplasmatische Domäne immer außerhalb der Vesikel liegt. Dies konnte anhand von Proteolysen gezeigt werden. Bei einer erfolgreichen Insertion der Transmembrandomäne ist diese durch die Lipide vor der Proteolyse geschützt, während die frei zugängliche periplasmatische Domäne hydrolysiert wird. Die geschützte und nicht hydrolysierte Transmembrandomäne kann später mittels 7
Gelelektrophorese nachgewiesen werden. Diese orientierte Insertion konnte für weitere OMPs wie z.B. BamA beobachtet werden (Patel und Kleinschmidt 2013). Die Faltungsausbeute von OMPs ist abhängig von den Kohlenstoffkettenlängen der verwendeten Lipide, von den Lipidkopfgruppen und auch vom pH-Wert der umgebenden Lösung (siehe Kleinschmidt 2006). Faltungsfördernd ist eine geringe Kohlenstoffkettenanzahl wie zum Beispiel 12 Kohlenstoffatome (diC12:0PC). Dies beruht am wahrscheinlichsten auf der hydrophoben Schichtdicke der LD die mit 19,5 ± 1 Å (Lewis und Engelman 1983) eher der hydrophoben Region vieler OMPs mit 22-25 Å entspricht (siehe Lee 2003), als die hydrophobe Schichtdicke von diC18:1PC mit 27 ± 1 Å (de Planque et al., 1998). Bei Verwendung der zwitterionischen PC (Phosphatidylcholin) Kopfgruppe zeigte sich bei vielen OMPs eine höhere Faltungsausbeute (Burgess et al., 2008), wohingegen die ebenfalls zwitterionische PE Kopfgruppe deutliche inhibierende Faltungseffekte zeigte (Patel und Kleinschmidt 2013, Gessmann et al., 2014). Der pH-Wert kann aufgrund der pKs-Werte von zahlreichen protonierbaren Aminosäureseitenketten die Ladung des OmpA sowie die Faltung und Aggregation des OmpA beeinflussen. Die Faltungsausbeute von OmpA (pI 5,6) kann zum Beispiel mit einer neutralen Kopfgruppe wie PC und gleichzeitiger negativer Proteinladung durch pH 10 erhöht werden. Im Vergleich dazu ist die OmpA Faltungsausbeute bei pH 7 geringer (Surrey und Jähnig 1995). Ein Vorteil von OmpA ist, dass es im SDS-Gel ohne vorhergehende Hitzedenaturierung in gefaltetem Zustand bei ca. 30 kDa migriert, wohingegen das ungefaltete bei ca. 35 kDa migriert (Schweizer et al., 1978). Dies beruht auf der kompakteren Form des gefalteten Zustandes, die weiteres migrieren durch die Gelmatrix ermöglicht. Gleichzeitig mit dieser Gelverschiebung kann eine nativ ausgebildete Sekundärstruktur und eine volle Funktionsfähigkeit des OmpA gezeigt werden (siehe Schweizer et al., 1978, Vogel und Jähnig 1986, Surrey und Jähnig 1992, Rodionova et al., 1995, Kleinschmidt und Tamm 1996, Kleinschmidt, den Blaauwen et al., 1999, Arora et al., 2000). Dies stellt eine einfache Methode zur Überprüfung der erfolgreichen Faltung und Insertion von OmpA und dessen Funktionalität ähnlich der nativen Form dar. Andere OMPs wie z.B. FomA und FhuA zeigen ähnliche Gelverschiebungen im gefalteten Zustand, bezeichnet als „heat modifiability“ (Schweizer et al., 1978, Kleinschmidt 2006, Hagan et al., 2013, Noinaj et al., 2015). 1.1.2 Faltungsmechanismus – Schrittweise Faltung und Insertion Einhergehend mit der erfolgreichen spontanen Rückfaltung von Proteinen in Modellmembranen in vitro kam die Frage nach dem dahinterstehenden Faltungsmechanismus auf. Bei α-Helicalen Membranproteinen bestehend aus mehreren α-Helices (helical bundle), 8
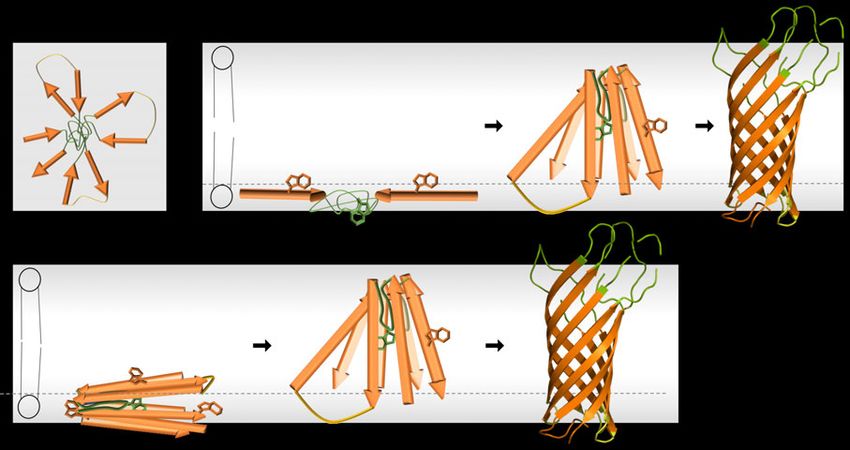
wird ein zweistufiger Faltungsmechanismus vermutet. Dabei falten und inserieren die einzelnen hydrophoben Helices autonom in die Membran und erst danach interagieren die einzelnen Helices lateral miteinander und bilden die Tertiärstruktur des Proteins aus (Popot et al., 1987, Popot und Engelman 1990). Solch ein Mechanismus erscheint für β-Fass TMP nicht möglich, da die einzelnen β-Stränge durch die alternierende Abfolge der hydrophoben und polaren Aminosäurereste amphipathisch sind. Diese Eigenschaft und die Notwendigkeit der Bildung von Wasserstoffbrückenbindungen zwischen den polaren Amid- und Carbonylgruppen der Polypeptidkette innerhalb der hydrophoben Region der Membran schließen die Membraninsertion einzelner β-Stränge aus (Kleinschmidt 2015). In den letzten Jahrzenten erfolgten einige Faltungsstudien die den Faltungs- und Insertionsmechanismus von β-Fass OMPs untersuchten. Neben einem einzigen, in einigen Fällen aus mehreren Schritten bestehenden Faltungsprozess (Kleinschmidt und Tamm 1996, Huysmans et al., 2012) zeigten unter bestimmten experimentellen Bedingungen mehrere Kinetikstudien verschiedenster OMPs auch zwei parallele Faltungsprozesse bzw. Faltungswege (Pocanschi, Apell et al., 2006, Patel et al., 2009, Huysmans et al., 2012). Faltungswege, die in mehreren Schritten verliefen, zeigten dabei in den ersten Schritten membranadsorbierte Faltungsintermediate und in den nachgeordneten Schritten die Ausbildung des β-Fasses. Die Membranadsorption erfolgte immer sehr schnell, wohingegen die Fassausbildung sehr viel länger dauerte. Daher ist der zweite Faltungsschritt geschwindigkeitslimitierend (siehe Kleinschmidt 2015). Der erste Faltungsschritt kann durch die Temperatur, aber auch durch die Wahl der Phospholipide und damit die Schichtdicke und Flexibilität der LD (Rawicz et al., 2000), sowie dem verwendeten Phospholipidüberschuss beeinflusst werden (Kleinschmidt 2006, Pocanschi, Apell et al., 2006, Huysmans et al., 2012). Studien mit kleinen unilamellaren Vesikeln aus Phospholipiden mit einer langen Kettenlänge von 18 Kohlenstoffatomen (diC18:1PC) zeigten einen genaueren Einblick in den Insertionsmechanismus von OmpA. Mittels Quenching von Tryptophanfluoreszenz durch an die Lipide gebundenes Brom konnte die Lokalisation der Tryptophanreste der Einzeltryptophanmutanten (ETM) des OmpA bestimmt werden. Die ETM besaßen jeweils ein Tryptophan an den nativen Tryptophanpositionen W7, W15, W57, W102 oder W143 die bis auf W7 alle im extrazellulären aromatischen Gürtel liegen (Abbildung 2 A). Dabei zeigte sich zunächst eine membranadsorbierte Form bei 2 °C, die für ca. eine Stunde stabil war. Die Tryptophanreste lagen hier 15-16 Å vom Zentrum der LD entfernt. Bei schrittweiser Temperaturerhöhung konnte eine synchrone Translokalisation der verschiedenen Tryptophanreste (außer W7) über die LD beobachtet werden (Abbildung 3). Dabei zeigten sich Entfernungen zum LD-Zentrum von zunächst 10-12 Å bis hin zu 0-7 Å. Nach erfolgreicher 9
Faltung lagen die Reste 9-10 Å vom LD-Zentrum entfernt. Bei dem Tryptophanrest W7 konnte entsprechend seiner Position im periplasmatischen aromatischen Gürtel keine Translokalisation über die Membran beobachtet werden, sondern ein konstanter Abstand von 10-11 Å zum Zentrum der LD (Kleinschmidt und Tamm 1999). In weiteren Studien konnte ebenfalls eine β- Strang Assoziation während des Insertionsprozesses innerhalb der LD gezeigt werden (Kleinschmidt et al., 2011). Für ein weiteres E. coli OMP, PagP (PhoPQ-activated gene P), konnte ebenso ein membranadsorbierter Übergangszustand (transition state) beobachtet werden, der relativ zur Membransenkrechten in einem Winkel geneigt ist, und zu einer sogenannten tilted insertion führt (Huysmans et al., 2010). Insgesamt deuten die Daten entsprechend Abbildung 3 auf eine synchrone Translokalisation der β-Stränge mit gleichzeitiger β-Fass Ausbildung hin. Diese Insertion könnte dabei, wie für PagP beobachtet, über einen geneigten Übergangszustand stattfinden. Ob dieser Faltungsmechanismus auch für größere OMPs zutrifft und inwiefern sich der Mechanismus durch Anwesenheit einer Insertionsmaschinerie ändert ist bislang unklar und wird zahlreich diskutiert. Der hohe Anteil an PE, der in vitro auch in kleinerem Anteil (30 %) stark inhibierend auf die spontane Faltung und Insertion von OMPs wirkt (Patel et al., 2009), zeigt aber bereits die Notwendigkeit einer Insertionsmaschinerie in Gram negativen Bakterien. Abbildung 3: Modell zur spontanen Faltung und Insertion von OmpA. Nach einer zunächst membranadsorbierten Form (links) erfolgt eine schrittweise synchrone Insertion der bereits ausgebildeten β-Stränge in die Membran bis hin zum vollständig ausgebildeten β-Fass (rechts) (entnommen aus Kleinschmidt 2003). 1.2 Der BAM Komplex In der IM von Gram negativen Bakterien sind bislang zwei Insertionsmaschinerien (Sec und YidC) bekannt, von denen Homologe in zahlreichen Organismen existieren (siehe Tsukazaki 2019). In der OM hingegen ist bislang nur ein Komplex, der BAM Komplex, bekannt (Wu et 10
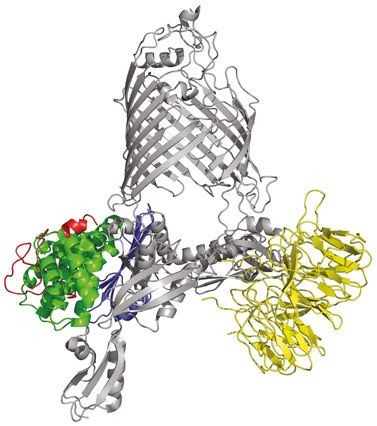
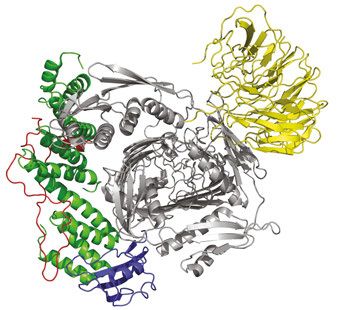
al., 2005). Aber auch hier existieren homologe Komplexe in anderen Organellen wie Chloroplasten (TOC-Komplex) (Schleiff und Soll 2005) und Mitochondrien (SAM-Komplex) (Kozjak et al., 2003, Paschen et al., 2003). Diese Maschinerien zum Einbau von OMPs in die OM von Gram negativen Bakterien, Chloroplasten und Mitochondrien bestehen alle aus einem TMP aus der Omp85 Familie und meistens mehreren Lipoproteinen. In E. coli besteht der BAM Komplex aus BamA und den vier Lipoproteinen BamB bis BamE, die direkt oder indirekt an BamA binden (Wu et al., 2005, Kim et al., 2007, Sklar et al., 2007a). Der Komplex weist in E. coli ein Molekulargewicht von ca. 208 kDa auf (Abbildung 4) (Ruiz et al., 2005, Bakelar et al., 2016b, Gu et al., 2016, Han et al., 2016). A B ~90° Abbildung 4: Kristallstruktur des E. coli BAM Komplexes. Der BAM Komplex bestehend aus BamA (grau), BamB (gelb), BamC (rot), BamD (grün) und BamE (blau) in Frontansicht (A) und um 90 ° gedreht (B), sodass das BamA β-Fass von der periplasmatischen Seite einsehbar ist und die Ringstruktur die von den POTRA (polypeptide transport associated) Domänen und den Lipoproteinen um das BamA β-Fass gebildet wird sichtbar ist (PDB Struktur: 5d0O, Gu et al., 2016). 1.2.1 Die Hauptkomponente des BAM Komplexes: BamA BamA auch bekannt als YaeT (E. coli) (Doerrler und Raetz 2005) oder Omp85 (Neisseria meningitides) (Voulhoux et al., 2003), gehört zur Omp85 Familie die essentiell ist für die Proteinbiogenese (Wu et al., 2005). Proteine der Omp85 Familie sind evolutionär konserviert und kommen neben Bakterien auch in Mitochondrien als Sam50 (Kozjak et al., 2003, Kutik et al., 2008)/ Tob55 (Paschen et al., 2003) und in Chloroplasten als Toc75 (Moslavac et al., 2005) vor. Charakteristisch sind die Proteine der Omp85 Familie aus einer C-terminalen β-Fass 11
Domäne mit 16 antiparallel angeordneten β-Strängen (Albrecht et al., 2014) und N-Terminal gelegenen POTRA (polypeptide transport associated) Domänen (Kim et al., 2007) in unterschiedlicher Anzahl aufgebaut. Sam50 besitzt eine POTRA Domäne, Toc75-V drei und BamA aus E. coli fünf (siehe Knowles et al., 2009). Die POTRA Domänen bestehen aus den Sekundärstrukturelementen β-α-α-β-β (Kim et al., 2007, Gatzeva-Topalova et al., 2008). Die im Periplasma gelegenen POTRA Domänen des BamA weisen in unterschiedlichen Kristallstrukturen unterschiedliche Konformationen auf. Dabei zeigen die POTRA Domänen zum einen eine „geschlossene“ und eine „offene“ Konformation (Abbildung 5 B). In der „geschlossenen“ Konformation verschließen die POTRA Domänen durch ihre Position das β- Fass von der periplasmatischen Seite. Zudem interagiert die POTRA Domäne 5, die am nächsten am β-Fass liegt, mit mehreren periplasmatischen Schleifen des BamA β-Fasses. Nach einer Drehung der POTRA Domänen, erscheint das β-Fass vom Periplasma her zugänglich, sodass dies als „offene“ Konformation beschrieben wird. Weiterhin zeigen sich in dieser Konformation keine Interaktionen zwischen POTRA 5 und den periplasmatischen Schleifen (Noinaj et al., 2013, Bakelar et al., 2016b, Gu et al., 2016). Diese zwei Zustände werden als möglicher Regulationsmechanismus für den Substratzugang zum BamA β-Fass angesehen. Weiterhin wird den POTRA Domänen eine wichtige Rolle in der Proteinassemblierung aufgrund von Interaktionen mit zum Beispiel der Chaperone SurA zugesprochen (Sklar et al., 2007b, Knowles et al., 2008, Bennion et al., 2010). Auch bei der 16-strängigen β-Fass Domäne des BamA sind zwei unterschiedliche Konformationszustände bekannt (Abbildung 5 A). Genauso wie bei den POTRA Domänen zeigte sich eine „offene“ und eine „geschlossene“ Konformation des β-Fasses (Abbildung 5) (Bakelar et al., 2016b, Gu et al., 2016). Durch eine Verschiebung der N-terminalen β-Stränge ist die Anzahl der Wasserstoffbrückenbindungen zwischen dem ersten und letzten β-Strang verringert und das β-Fass erscheint geöffnet. Ob die laterale Öffnung des BamA β-Fasses notwendig für die Faltung und Insertion von OMPs ist, ist derzeit umstritten. Eine Studie in der das BamA β-Fass mittels disulfide cross-linking verschlossen wurde, zeigte eine letale Wirkung (Noinaj et al., 2014). Andere Studien hingegen, bei denen ebenfalls mittels disulfide cross- linking das β-Fass geschlossen gehalten wurde, zeigten keine Veränderung der BamA bedingten Faltungs- und Insertionseffizienz bei kleinen OMPs wie OmpX und tOmpA (8 β- Stränge) (Doerner und Sousa 2017, Schiffrin et al., 2017). Daher wird eine β-Fass Öffnung nur bei Faltung und Insertion von größeren OMPs vermutet. Ob diese Öffnung dann zur Translokalisation der extrazellulären Schleifen des OMP oder aber zur Faltung und Insertion der β-Stränge dient, ist unklar. 12
Neben der destabilisierten Bindung des ersten und letzten β-Stranges, zeigen die Kristallstrukturen von BamA eine asymmetrische Form des β-Fasses. Die Fassregion um das C-terminale Ende weist eine geringere Höhe auf, als die gegenüberliegende Seite (Abbildung 4 A, 5 A). Gleichzeitig konnte eine verringerte Schichtdicke der umliegenden Membran in der Region des C-terminalen Endes bestimmt werden (Noinaj et al., 2013, Sinnige et al., 2014). Die Hauptfunktion des BamAs könnte daher darin liegen die Aktivierungsenergie zur Faltung von OMPs durch Verjüngung der LD Schichtdicke herabzusetzen. A B P5 “geschlossen” P5 “offen” P5 “geschlossen” P5 “offen” Abbildung 5: BamA β-Fass zeigt unterschiedliche Konformationszustände. Gezeigt sind die unterschiedlichen Konformationszustände des β-Fasses (A) und der POTRA Domäne 5 (B). Die PDB Struktur 4C4V (Albrecht et al., 2014) in blau dargestellt und die PDB Struktur 5EKQ (Bakelar et al., 2016a) in grün. Während die Struktur in grün dargestellt ein geöffnetes β-Fass zeigt, ist gleichzeitig eine Bewegung der POTRA 5 vor die periplasmatische Öffnung des β- Fasses zu beobachten wodurch sich ein zum Periplasma geschlossenes β-Fass zeigt. Die Struktur in blau zeigt genau die gegenteiligen Zustände (entnommen aus Bakelar et al., 2016b). 1.2.2 Lipoproteine des BAM Komplexes Die vier Lipoproteine des E. coli BAM Komplexes binden direkt oder indirekt an die POTRA Domänen des BamA. BamC (NlpB) und BamE (SmpA) bilden mit BamD (YfiO) einen Subkomplex und sind nur indirekt über BamD an BamA gebunden. BamB (YfgL) bindet unabhängig von diesem Subkomplex an BamA (Abbildung 4) (Wu et al., 2005, Hagan et al., 13
2010). BamB-BamE zählen genauer zu den Lipidanker Proteinen, sodass sie mittels N- terminalem Lipidanker in die innere Schicht der OM verankert sind (Sankaran und Wu 1994). Zwar sind nur BamA und BamD essentiell (Malinverni et al., 2006, Kim et al., 2007), jedoch arbeitet der BAM Komplex nur in Anwesenheit aller Bam-Lipoproteine am effektivsten (Wu et al., 2005, Charlson et al., 2006, Sklar et al., 2007a). BamB ist mit 41,9 kDa das größte Lipoprotein des BAM Komplexes. Es bildet mit acht Blättern, bestehend aus je vier antiparallel angeordneten β-Strängen, eine propellerartige Struktur (Heuck et al., 2011). BamB bindet an BamA POTRA 2 – 5 (Kim et al., 2007, Chen et al., 2016) und zeigt zu BamD überlappende Funktionen (Heuck et al., 2011). BamB sowie BamD spielen eine wichtige Rolle in der BamA Faltung (Tellez und Misra 2012). Ebenso trägt BamB zur effizienten Faltung und Insertion verschiedener OMPs bei (Ureta et al., 2007, Palomino et al., 2011). BamC (36,8 kDa) zeigt zwei definierte Helix-Griff Domänen (helix-grip), die als Protein-Protein Interaktionsstellen dienen könnten (Warner et al., 2011). Zusammen mit BamE spielt es eine Rolle in Dynamik und Funktion von BamA durch die Stabilisierung der BamA-BamD Verbindung (Malinverni et al., 2006, Rigel et al., 2012). Das an POTRA 5 bindende (Kim et al., 2007) essentielle Lipoprotein BamD (27,2 kDa) ist aus 10 α-Helices aufgebaut, die wiederum in fünf tetratricopeptide repeat (TPR) unterteilt werden können. Ein TPR Motiv besteht aus zwei benachbarten α-Helices, die durch eine Schleife verbunden sind. Das Motiv ist bekannt als Protein-Protein Interaktionsstelle und besitzt zudem mehrere Furchen für die Bindung von Liganden (Albrecht und Zeth 2011, Sandoval et al., 2011, Dong et al., 2012). BamD dient daher wahrscheinlich als OMP Rezeptor. Das sogenannte β-Signal, eine C-terminale Peptidsequenz die bei allen OMPs in Prokaryoten und Eukaryoten vorkommt (siehe Ricci und Silhavy 2019), dient dabei als Erkennungssequenz für BamD (Hagan et al., 2015). BamD erkennt und bindet neben anderen OMPs auch ungefaltetes BamA an dessen β-Signalsequenz und beeinflusst Bindung und Lokalisation der OMPs in der OM (Hagan et al., 2015, Ricci und Silhavy 2019). Im gebildeten BAM Komplex induziert BamD eine BamA Konformationsänderung, die die Faltung und Insertion von OMPs erleichtert (McCabe et al., 2017, Lee et al., 2018). Das kleinste Lipoprotein des BAM Komplexes, BamE (12,3 kDa) ist aus zwei N-Terminal gelegenen α- Helices und einem C-Terminal gelegenen β-Faltblatt bestehend aus drei antiparallel angeordneten β-Strängen aufgebaut (Kim et al., 2011). Neben der stabilisierenden Wirkung auf die BamA-BamD Bindung ist außerdem bekannt, dass es PG bindet (Knowles et al., 2011). 14
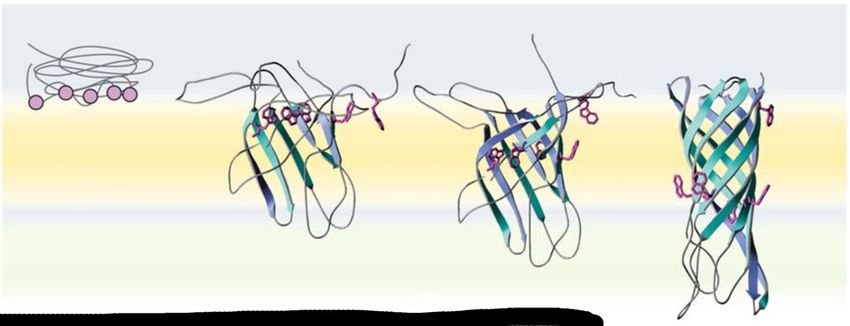
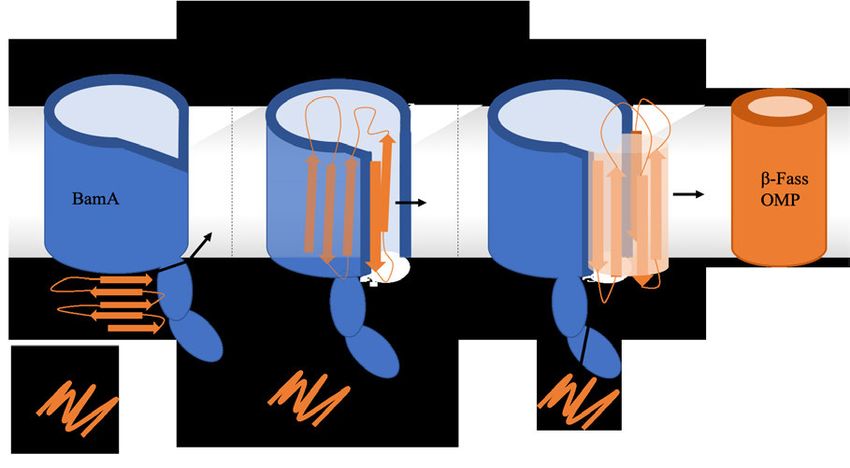
1.3 Modelle zum BamA-abhängigen Faltungsmechanismus von OMPs Für die BamA katalysierte Faltung und Insertion von OMPs gibt es mehrere Modelle, die hauptsächlich versuchen, die Frage der Translokation und Faltung der β-Stränge der OMPs zu beantworten. Die als das ‚budding model‘ bekannte Theorie erklärt diese Vorgänge zum Beispiel so, dass sich das BamA β-Fass öffnet und die β-Stränge 1 und 16 als Ausgangspunkt/Vorlage für die Bildung von β-Strängen des OMP dienen. Daraufhin wird das BamA β-Fass durch die OMP β-Stränge erweitert und es entsteht ein hybrid BamA-OMP Fass. Nachdem alle β-Stränge des OMP β-Fasses ausgebildet sind wird das Fass von BamA in die OM abgestoßen und das OMP ist in die Membran inseriert (Abbildung 6 C) (Noinaj et al., 2014). Ein weiteres Modell besagt, dass eine Bindung des N-terminalen Endes des OMP an die BamA β-Fass Öffnung entsteht während sich die OMP-β-Stränge im Inneren des BamA β- Fasses bilden. Dabei wirkt das innere des BamA β-Fasses durch Bindung einzelner OMP β- Stränge unterstützend. Eine Paarung des N-terminalen Endes mit dem C-terminalen Ende des OMP schließt getrieben durch OMP Schleifeninteraktionen das β-Fass (Abbildung 6 B) (Albrecht et al., 2014, Lee et al., 2019). Bei Toc75, das BamA Homolog aus der Omp85 Familie aus Chloroplasten wird ebenfalls eine β-Fass Öffnung vermutet (Ganesan und Theg 2019). Weiterhin gibt es das sogenannte ‚assisted model‘. In dieser Theorie ist das BamA lediglich passiv beteiligt. Die Schichtdicke der OM ist am C-terminalen Ende des BamAs (β-Fass Öffnung) deutlich verringert (Sinnige et al., 2014), sodass dadurch die Aktivierungsenergie zur Faltung und Insertion von OMPs in die OM herabgesetzt sein könnte. Die OMPs falten und inserieren dabei getrieben durch thermodynamische Prozesse selbstständig (ΔG° < 0, spontane Faltung) in die OM (Abbildung 6 A) (Noinaj et al., 2013, Patel und Kleinschmidt 2013, Gessmann et al., 2014, Sinnige et al., 2014) so wie dies für die Faltung von verschiedenen OMPs wie OmpA oder PagP in Abwesenheit von BamA für Modellmembranen experimentell vielfach bestätigt wurde (siehe Kleinschmidt 2015). Unterstützt wird dieses Modell durch experimentelle Studien zur Funktion des BAM-Komplexes bei der Faltung des 26-strängigen β-Fasses LptD, des LPS-Transporters (Lee et al., 2016). Für Sam50, der Hauptkomponente des SAM Komplexes aus Mitochondrien, wurden die gleichen Modelle für den Sam50-abhängigen OMP Faltungsmechanismus postuliert, wie hier für BamA vorgestellt (Hohr et al., 2015, Hohr et al., 2018). 15
Abbildung 6: Modelle der BamA-abhängigen Faltung und Insertion von β-Fass OMPs. (A) Die Sekundärstrukturausbildung des Prä-OMP erfolgt an der Membran. Die Insertion erfolgt durch die BamA induzierte geringere Schichtdicke der Membran („assisted model“). Durch die geringer Schichtdicke, wird die Aktivierungsenergie herabgesetzt und es kommt zu einer spontanen Faltung. (B) Die Sekundärstrukturausbildung erfolgt innerhalb des BamA β- Fasses und die Insertion in die Membran wird durch die laterale BamA β-Fass Öffnung möglich. (C) Mit dem ersten und letzten β-Strang des BamA β-Fasses als Vorlage werden die β-Stränge des Prä-OMP ausgebildet, wodurch ein hybrid β-Fass entsteht. Nach vollständiger Ausbildung der β-Fass Struktur wird das OMP lateral in die Membran entlassen („budding model“). 1.4 Motivation und Ziel der Arbeit OMPs sind der erste Kommunikationspunkt zwischen Zellen und deren Umgebung. Sie können als Kontaktpunkte z.B. in medizinischen Fragestellungen (Entwicklung neuer Antibiotika) eine bedeutende Rolle spielen. Das Verständnis des Mechanismus der Faltung und Insertion in die OM ist daher von zentraler Bedeutung, aber bislang ist nur sehr wenig darüber bekannt. In vitro kann eine spontane Faltung von E. coli OMPs in Modellmembranen beobachtet werden und verschiedene Studien geben Hinweise auf den dahinterstehenden Faltungs- und Insertionsmechanismus. In E. coli ist der BAM Komplex involviert in die Proteinbiogenese. Ob und wie sich der Faltungsmechanismus von OMPs in Anwesenheit des BAM Komplexes im Vergleich zur spontanen Faltung in Abwesenheit des BAM Komplexes in vitro verändert wird rege diskutiert, konnte aber noch nicht experimentell aufgeklärt werden. Bisher durchgeführte Studien dazu nutzten Methoden wie disulfide cross-linking, wodurch eine Interaktion zwischen BamA und einem Klienten-OMP erzwungen wird. 16
In dieser Arbeit wird dazu ein komplett neuer Ansatz zur Untersuchung des Faltungsmechanismus von Membranproteinen des β-Fass Typs vorgestellt. Dieser besteht aus einer Kombination von molekularbiologischen und biophysikalischen Verfahren mit der Konzeption und Erprobung neuer Modellmembranen für Faltungsstudien. Die spezifischen Ziele waren: 1. Die Etablierung eines Systems, indem BamA Wechselwirkungen mit entfalteten OMPs untersucht werden können ohne massiv in die Interaktionen einzugreifen, so wie dies z.B. durch chemische Reaktionen (z.B. disulfide cross-linking) der Fall ist. Für die lokale Untersuchung von Konformationsänderungen in Proteinen oder Untersuchungen (auch des zeitlichen Verlaufs) von lokalen Protein-Protein-Wechselwirkungen bieten sich hoch sensitive fluoreszenzspektroskopische Verfahren in Kombination mit ortsgerichteter Mutagenese an. Diese Technik erlaubt sowohl das Studium lokaler Änderungen in der Umgebung des Fluorophors im Protein als auch die lokale Untersuchung von intermolekularen Wechselwirkungen zwischen verschiedenen Proteinen, wie z.B. zwischen BamA und einem (ungefalteten oder teilgefalteten) Klienten-OMP. Mit geeigneten Paaren von Fluorophoren lassen sich auch Abstandsmessungen durch Messung des Fluoreszenz- Resonanzenergietransfers durchführen. Die Fluoreszenzmarker (meist kommerziell erhältliche Fluoreszenzlabel) lassen sich z.B. durch proteinchemische Reaktionen kovalent z.B. an die Sulfhydrylgruppe einer Einzelcysteinmutante des Proteins koppeln. Auch kann die intrinsische Fluoreszenz der Aminosäure Tryptophan ausgenutzt werden, wozu ETM des Proteins präpariert werden müssen. Für diese Arten von ortsgerichteter Fluoreszenzspektroskopie muss zunächst getestet werden, ob es möglich ist, die natürlich vorhandenen zwei Cystein- bzw. 13 Tryptophanreste im E. coli BamA konservativ, d. h. ohne Verlust der Funktion des BamA, gegen möglichst ähnliche Aminosäuren auszutauschen. Die Trp- und Cys- freie Mutante des BamAs kann dann als Ausgangsplattform verwendet werden, um fluoreszenzspektroskopisch nutzbare Punktmutanten zu präparieren. 2. Neben dieser BamA Mutante soll ein Modellmembransystem, aufgebaut aus Dilauroylphospholipiden (12 Kohlenstoffatomen) mit verschiedenen Kopfgruppen, entwickelt werden. In solche dünnen und flexiblen Modellmembranen falten und inserieren erfahrungsgemäß viele verschiedene OMPs, wahrscheinlich weil die hydrophobe Schichtdicke am ehesten der hydrophoben Transmembranregion vieler OMPs entspricht. Bislang gelang es aber nicht Faltungsintermediate in solch einer Membran zu stabilisieren und zu analysieren. Dieses System soll durch geeignete Auswahl von Lipiden und physikalischen Bedingungen 17
eine Verringerung der Faltungsreaktionsgeschwindigkeit von OMPs in Ab- und Anwesenheit von BamA gewährleisten, bei gleichzeitiger Stabilisierung von OMP Faltungsintermediaten, die dann analysiert werden können. 3. Mit Hilfe der in den Meilensteinen 1 und 2 entwickelten Verfahren sollen schließlich die BamA-abhängige OMP-Faltung und der Membraneinbau anhand von Faltungsintermediaten, hier am Beispiel von OmpA, analysiert werden, um eine Aufklärung des Faltungsmechanismus in Anwesenheit von BamA zu erreichen. Dabei werden OmpA ETM des OmpA verwendet, die an verschiedenen Positionen im Protein wie z.B. in β-Fasssträngen und extrazellulären Schleifen einen Tryptophanrest aufweisen. Damit soll eine ortspezifische fluoreszenzspektroskopische Analyse der Faltung und Insertion in Abhängigkeit von BamA erfolgen. 18
2. Materialien und Methoden 2.1 Materialien Alle nicht extra aufgeführten Materialien wurden von den Firmen Carl Roth (Karlsruhe, Deutschland) und Sarstedt (Nümbrecht, Deutschland) bezogen. 2.1.1 Bakterienstämme Escherichia coli: - BL21(DE3) omp8 fhuA [F ̄, ompT hsdSB (rB− mB−) gal dcm (DE3) ΔlamB ompF:: Tn5 ΔompA ΔompC ΔfhuA] - XL10-Gold TetR D(mcrA)183 D(mcrCB-hsdSMR-mrr)173 endA1 supE44 thi-1 recA1 gyrA96 relA1 lac Hte [F´ proAB lacIqZDM15 Tn10 (TetR) Amy CamR] 2.1.2 Oligonukleotide Die Oligonukleotide wurden mittels der Software SnapGene (GSL Biotech LLC, San Diego, USA) designt und von Eurofins Scientific (Luxemburg, Luxemburg) bezogen. Tabelle 1: Liste der Oligonukleotide die zur Sequenzierung oder Mutagenese (Restriktionssequenz in fett gedruckt) der OmpA Mutanten verwendet wurden. Beschreibung Sequenz T7_pET22b fw 5'-TAATACGACTCACTATAGGG-3' (Sequenzierung) pET-RP_pET22b rev 5'-CTAGTTATTGCTCAGCGG-3' (Sequenzierung) OmpA W7 NdeI fw 5'-CACATATGGCTCCGAAAGATAACACCTGGTAC-3' OmpA F7 NdeI fw 5'-CACATATGGCTCCGAAAGATAACACCTTCTAC-3' OmpA XhoI rev 5'-CTGCTCGAGTTAAGCCTGCGGCTGAGTTACA-3' 19
2.1.3 Plasmide Alle verwendeten Vektoren enthalten zur Selektion einen Genabschnitt für die Ampicillinresistenz. Tabelle 2: Aufgelistet sind die verwendeten Plasmide unter Angabe der Vektoren, eingefügter Genabschnitt, sonstige Informationen und die Bezugsquelle. (SS = Signalsequenz) Vektor eingefügtes Gen Information Bezugsquelle pET22b(+) C-terminaler His-Tag Novagen pJET blunt ends Thermo Fisher Scientific, Waltham, USA pET15b bamA - SS + N-terminaler His-Tag pET15_EcOMP85 (Patel und Kleinschmidt 2013) pET15b bamA WaF CaA - SS + N-terminaler His-Tag Trenzyme GmbH, Konstanz, WaF CaA Deutschland pET22b ompA wt - SS Sonja Braun 2013 pET22b ompA WaF CaA - SS WaF CaA pET22b185 (Qu et al., 2009) pJET ompA W7 - SS WaF diese Arbeit pET22b ompA W7 - SS WaF diese Arbeit, Sonja Kasten pJET ompA W15 - SS WaF diese Arbeit pET22b ompA W15 - SS WaF diese Arbeit, Sonja Kasten pJET ompA W102 - SS WaF diese Arbeit pET22b ompA W102 - SS WaF diese Arbeit, Sonja Kasten pJET ompA W143 - SS WaF diese Arbeit pET22b ompA W143 - SS WaF diese Arbeit, Sonja Kasten pET22b ompA I24W - SS WaF CaA pET24 (Qu et al., 2009) pET22b ompA V67W - SS WaF CaA pET67 (Qu et al., 2009) pET22b ompA V110W - SS WaF CaA pET110 (Qu et al., 2009) pET22b ompA I153W - SS WaF CaA pET153 (Qu et al., 2009) pTRC99A ompA W7 + SS WaF pET1102 (Kleinschmidt, den Blaauwen et al., 1999) pTRC99A ompA W15 + SS WaF pET1115 (Kleinschmidt, den Blaauwen et al., 1999) pTRC99A ompA W102 + SS WaF pET186 (Kleinschmidt, den Blaauwen et al., 1999) pTRC99A ompA W143 + SS WaF pET1103 (Kleinschmidt, den Blaauwen et al., 1999) 20
2.2 Methoden 2.2.1 Homologie Sequenzvergleich Um konservierte Aminosäuren in der BamA Proteinsequenz aus E. coli im Vergleich zu BamA Homologen anderer Gram negativer Bakterien (siehe Tabelle 3) zu bestimmen wurden die Proteinsequenzen mittels SnapGene (GSL Biotech LLC, San Diego, USA) über das Clustal Omega Sequenzvergleichsprogramm abgeglichen. BamA aus E. coli wurde als Vergleichssequenz genutzt und eine Grenzschwelle von 50 % angewendet. Tabelle 3: Gram negative Bakterienarten die für den Homologie Sequenzvergleich der BamA Homologe verwendet wurden. Aufgelistet ist die Bakterienart, Abteilung, Klasse, Familie und der NCBI Code das BamA Homolog aus der Bakterienart. uniprot, NCBI Bakterienart Abteilung Klasse Familie Datenbank Escherichia coli Proteobacteria Gammaproteobacteria Enterobacteriaceae P0A940 Shigella flexneri Proteobacteria Gammaproteobacteria Enterobacteriaceae P0A943 Salmonella typhimurium Proteobacteria Gammaproteobacteria Enterobacteriaceae Q8ZRP0 Legionella longbeachae Proteobacteria Gammaproteobacteria Legionellaceae D3HKZ9 Stenotrophomonas Proteobacteria Gammaproteobacteria Xanthomonadaceae B8L348 Pseudomonas aeruginosa Proteobacteria Gammaproteobacteria Pseudomonadaceae Q9HXY4 Helicobacter pylori Proteobacteria Epsilonproteobacteria Helicobacteraceae O25369 Neisseria gonorrhoeae Proteobacteria Betaproteobacteria Neisseriaceae P95359 Neisseria meningitidis Proteobacteria Betaproteobacteria Neisseriaceae Q9K1H0 Caulobacter vibrioides Proteobacteria Alphabroteobacteria Caulobacteraceae Q9A711 Borrelia turcica Spirochaetae Spirochaetes Spirochaetaceae A0A386PMY4 Pirellula sp. Planctomycea Planctomycea Planctomyceaceae A0A142XW14 Chlamydia pneumoniae Chlamydiae Chlamydiaceae Chlamydiaceae A0A0F7X3D0 2.2.2 Gezielte Punkt-Mutagenese Die gezielte Insertion, Deletion oder Substitution einzelner Basenpaare innerhalb einer Plasmid-DNA-Sequenz dient zur Änderung der Aminosäuresequenz oder zum Verschieben des Leserahmens. Hier wurde das QuikChange XL Site-Directed Mutagenese Kit (Agilent Technologies, Santa Clara, USA) genutzt um Einzelcystein- oder Einzeltryptophanmutanten 21
für Protein-Protein und Protein-Lipid Interaktionsstudien herzustellen. Die Mutagenese und anschließende Transformation in XL10-Gold ultrakompetente Zellen wurde nach Herstellerangaben des QuikChange XL Site-Directed Mutagenese Kit durchgeführt. Eine Änderung war das verwenden der PfuUltra Polymerase (Thermo Fisher Scientific, Waltham, USA) anstatt der im Kit verwendeten PfuTurbo Polymerase. Die PfuUltra weist einen dreifach niedrigeren mittleren Fehler auf als die PfuTurbo. Die Mutation wurde mit Hilfe von Nukleotid Sequenzierung überprüft (Eurofins Scientific, Luxemburg, Luxemburg). Alle verwendeten Oligonukleotide sind in Tabelle 1 aufgeführt. 2.2.3 Mutagenese zur Deletion der Signalsequenz von OmpA Die Signalsequenz von OmpA ist 20 Aminosäuren lang und führt nach erfolgter Translation zum Transport hin zur OM. Die Expression und Isolation von OMPs aus der OM führt meist zu kleinen und verunreinigten Mengen. Die Expression und Isolation aus cytosolischen Einschlusskörpern (inclusion bodies) hingegen führt zu größeren und reineren Mengen (siehe Übersichtsartikel dazu (Popot 2014). Für diese Art der Reinigung muss die Signalsequenz deletiert werden, damit das Protein im Cytoplasma verbleibt. Die ETM von OmpA (W7, W15, W102 und W143) von (Kleinschmidt, den Blaauwen et al., 1999) lagen im Vektor pTRC99A mit Signalsequenz vor. Daher wurden sie ohne Signalsequenz in den Expressionsvektor pET22b(+) kloniert. Dies wurde über eine PCR (polymerase chain reaction) mit synthetischen Oligonukleotiden erreicht, die die OmpA-Sequenz ohne Signalsequenz flankierten und jeweils eine unterschiedliche Restriktionsschnittstelle enthielten. Für das vorwärts gerichtete Oligonukleotid wurde die NdeI Schnittstelle gewählt und für das rückwärtsgerichtete die XhoI Schnittstelle (Tabelle 1). Nach der Amplifikation mittels PCR wurde das DNA-Fragment in den pJET Vektor ligiert (nach Herstellerangaben, CloneJET PCR Cloning Kit, Thermo Fischer Scientific, Waltham, USA). Bei dem Vektor handelt es sich um einen blunt-end („glattes Ende“) Vektor, wodurch das PCR-Produkt ohne Restriktionsverdau ligiert werden kann. Nach der Hitzeschock-Transformation in XL10-Gold ultrakompetente Zellen, konnte die via MiniPrep (Thermo Fischer Scientific, Waltham, USA) isolierte DNA mit NdeI und XhoI (Thermo Fischer Scientific, Waltham, USA) verdaut werden. Der Restriktionsverdau wurde anhand eines Agarosegels überprüft, die DNA daraus ausgeschnitten und mit Hilfe des Gelextraction Kits (Quiagen, Hilden, Deutschland) gereinigt. Danach erfolgte die Ligation in den NdeI/XhoI verdauten pET22b Vektor und die Hitzeschock Transformation in XL10-Gold Bakterien (T4 Ligase, Thermo Fischer Scientific, Waltham, USA). Insofern die Sequenzierung eine erfolgreiche Mutagenese zeigte, wurde das Plasmid in die Expressionszellen BL21(DE3) omp8 fhuA mittels Hitzeschock transformiert. 22
2.2.4 Expression und Reinigung von BamA und OmpA Die Expression und Reinigung des wildtyp BamA (wt-BamA) und der Mutante WaF, CaA- BamA (siehe Tabelle 2) erfolgte entsprechend des Expressions- und Reinigungsprotokolls aus 2013 (Patel und Kleinschmidt 2013). Die Anionenaustauschchromatographie wurde mit einer Q-Sepharose Säule unter Verwendung einer ÄKTAprime plus (GE Healthcare Life Sciences, Freiburg) oder NGC (Bio-Rad Laboratories, Feldkirchen, Deutschland) durchgeführt. Lediglich der NaCl-Gradient zur Elution wurde auf 0-500 mM NaCl erhöht. Nach der Elution, wurden die BamA-enthaltenden Fraktionen gegen 15 mM Tris(hydroxymethyl)aminomethan (Tris), 8 M Harnstoff, pH 8,5 in Dialyseschläuchen (Spectrum Spectra/Por 1, MWCO 6000- 8000 Dalton, 50 mm Breite, Thermo Fisher Scientific, Waltham, USA) dialysiert um das NaCl zu entfernen. Der wildtyp OmpA (wt-OmpA) und alle Mutanten wurden entsprechend des Protokolls aus 2009 (Qu et al., 2009) exprimiert und gereinigt. Die Fällung, mittels Isopropanol, während der Isolation wurde ausgelassen, da sie keinerlei Einfluss auf die Reinheit des OmpAs zeigte. Die Anionenaustauschchromatographie wurde mit einer Q-Sepharose Säule unter Verwendung einer ÄKTAprime plus (GE Healthcare Life Sciences, Freiburg) oder NGC (Bio-Rad Laboratories, Feldkirchen, Deutschland) durchgeführt. Das Ankonzentrieren der Proteine aus den Fraktionen der Elution erfolgte mit Hilfe der Vivaspin 20 Ultrafiltration Unit (Sartorius, Göttingen, Deutschland) Zentrifugenkonzentratoren nach Herstellerangaben bei 10 °C. Die Konzentrationen wurden durch die Methode nach Lowry et al. aus 1951 bestimmt. Hierbei wurde BSA (bovines serum albumin) als Standard verwendet. 2.2.5 Präparation von Lipidfilmen und großen unilamellaren Vesikeln Für alle Faltungen wurden, neben wässerigen Lösungen von Detergensmizellen aus Lauryldimethylaminoxid (LDAO), Phospholipide mit einer Kettenlänge von 12 Kohlenstoffatomen verwendet. Dabei wurden 1,2-Dilauroyl-sn-glycero-3-phosphocholine (DLPC), 1,2-Dilauroyl-sn-glycero-3-phosphoethanolamine (DLPE), 1,2-Dilauroyl-sn-glycero- 3-phosphoglycerol (DLPG) (Avanti Polar Lipids, Alabama, USA) in einem molaren Verhältnis von 5:3:2 verwendet. Insgesamt wurden 10 bis 20 mg Phospholipide in braune Glasfläschchen eingewogen und unter einem Abzug in Methanol und Chloroform (Verhältnis 1:1 (v/v)) gelöst. Die organischen Lösungsmittel wurden unter einem Abzug mittels eines Stickstoffstroms verblasen. Daraufhin wurden die Lipidfilme in einem Exsikkator unter Vakuum für vier Stunden getrocknet. Zur Lagerung wurden sie unter Argon versetzt bei -20 °C gelagert. 23
Für die Herstellung von großen unilamellaren Vesikeln (LUVs, large unilamellar vesicles) wurde ein Lipidfilm in dem entsprechenden Puffer der geplanten Faltung durch Mischen mittels Vortexer gelöst (auf 10 mg/ml). Danach folgten sieben Schritte von Einfrieren in flüssigem Stickstoff (30 s) und Auftauen bei 55 °C 300 rpm im Thermoschüttler. Dabei entstehen unilamellare Vesikel unterschiedlicher Größe. Um eine einheitliche Größe in allen Experimenten zu gewährleisten, wurden die Vesikel mit Hilfe eines Extruders (Avanti Polar Lipids, Alabama, USA) und einer Membran mit einer Porengröße von 100 nm (Nucleopore Track-Etch-Membran PC, GE Whatman, Clifton, USA) auf eine einheitliche Größe von 100 nm Durchmesser gebracht. Die LUVs wurden direkt verwendet oder bei 4 °C gelagert. 2.2.6 Faltung in LUVs und LDAO Die Faltung der in 8 M Harnstoff denaturierten Proteine BamA und OmpA erfolgte jeweils über die Verdünnung des Harnstoffs auf maximal 0,6 M in Anwesenheit von LUVs oder LDAO- Mizellen. Faltung von BamA Zur Faltung von BamA in LUVs (LD bestehend aus DLPC, DLPE und DLPG in einem Verhältnis von 5:3:2, oder DLPE, DLPG in 4:1) wurde dieses in einem molaren Verhältnis Protein zu LD von 1 zu 1000 in 10 mM Borax (Dinatriumtetraborat-Decahydrat), pH 10 bei 4 °C, 300 rpm für 48 h inkubiert. Die finale Konzentration im Faltungsansatz betrug 6,3 µM BamA und 6,3 mM LUVs. Zur Faltung in LDAO-Mizellen wurden 10 µM BamA in 10 mM LDAO ebenfalls in 10 mM Borax, pH 10 bei 4°C, 300 rpm für 48 h inkubiert. Die Harnstoffkonzentration in den finalen Faltungsansätzen betrug maximal 0,6 M. OmpA Faltungskinetik in Ab- und Anwesenheit von BamA Kinetische Studien (KTSE, kinetics of tertiary structure formation by electrophoresis) zur OmpA-Faltung in LUVs wurden unter verschiedenen Bedingungen durchgeführt. Um den Einfluss von BamA auf die OmpA-Faltung zu untersuchen, wurden Faltungskinetiken in reine LD (DLPC, DLPE, DLPG, 5:3:2) und mit in die LD inseriertem BamA (LD⋅[BamA]) bei pH 8 (50 mM HEPES (2-(4-(2-Hydroxyethyl)-1-piperazinyl)-ethansulfonsäure), 2 mM EDTA (Ethylendiamintetraessigsäure)), 300 rpm und 30 °C (40 °C) parallel durchgeführt. Die finalen Konzentrationen in den Faltungsansätzen waren 7,1 µM OmpA, 2,13 µM BamA und 2,13 mM LUVs (LD bestehend aus DLPC, DLPE, DLPG in einem Verhältnis von 5:3:2) und maximal 0,3 M Harnstoff. Die OmpA-Faltung wurde von 2 bis 180 min in acht Zeitschritten (bei 40 °C, 1-90 min in acht Zeitschritten) aufgezeichnet. An jedem Zeitpunkt wurde eine Probe (16 µl) 24
Sie können auch lesen

















































