3.1 Körpereigene, elektromagnetische Biosignale - nt.tuwien ...
←
→
Transkription von Seiteninhalten
Wenn Ihr Browser die Seite nicht korrekt rendert, bitte, lesen Sie den Inhalt der Seite unten
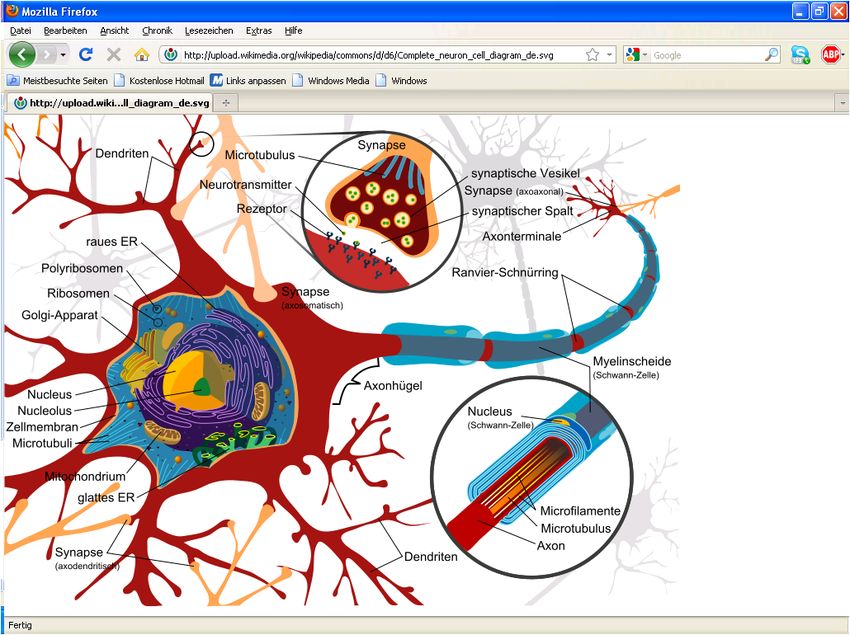
3.1 Körpereigene, elektromagnetische Biosignale 3.1.1 Funktionsweise von Nervenzellen, zum Thema „Nervenreizleitung“ Aufbau der Nervenzelle Eine Nervenzelle (Neuron) ist eine auf Erregungsleitung spezialisierte Zelle, vgl. Bild 3.1/1. Das menschliche Gehirn besteht schätzungsweise aus 1011 bis 1012 Nervenzellen. Eine typische Säugetier-Nerven- zelle ist aus Dendriten, dem Zell- körper und einem Axon aufge- baut. Dieser Zellfortsatz kann sehr lang sein und ermöglicht eine Erregungsleitung über weite Stre- cken. Dabei läuft ein elektrisches Signal durch das Axon. Es wird erzeugt, indem bestimmte Ionen gezielt durch die Zellmembran durchgeschleust werden. Das Axonende steht über Synapsen, an denen das Signal chemisch (seltener elektrisch) weitergege- ben wird, mit anderen Nervenzel- len oder Empfängerzellen in Ver- bindung. Bild 3.1/1: Nervenzelle Der Zellkörper (Soma) enthält neben dem Zellkern diverse Organellen. Das Soma hat in Abhängigkeit vom Nervenzelltypus eine Größe zwischen 5 und mehr als 100 µm. Es produ- ziert die Neurotransmitter für die Reizweiterleitung. Die an Dendriten eintreffenden Signale werden im Soma weiterverarbeitet. Das geschieht durch räumliche und zeitliche Summation von Änderungen des Membranpotentials. Vom Soma wachsen Fortsätze aus, die Dendriten. Das sind feine plasmatische Verästelun- gen des Zellkörpers, die über Synapsen den Kontakt zu anderen Nervenzellen herstellen. Sie empfangen Aktionspotentiale von anderen Neuronen durch deren Axone. Der Dendriten- baum einer einzigen menschlichen Nervenzelle kann mit 100.000 bis 200.000 Fasern ande- rer Neuronen im Kontakt stehen (die Potenziale aller Eingänge werden summiert). Dies ermög- licht eine extrem hochgradige Parallelisierung der Signalisierung zwischen den Nervenzellen. An das Soma angesetzt ist der Axonhügel. Das Schwellenpotential des Axonhügels ist stark reduziert. Es wirkt dadurch als Initialsegment. Von hier aus werden die Aktionspotentiale an das Axon weitergeleitet. Durch das niedrige Schwellenpotential und die Nähe des Axon- hügels zum Zellkörper ist sichergestellt, dass bei einer Erregung der Zelle das Aktionspoten- tial nur an einem Ort entsteht und weitergeleitet wird. Das ist wichtig für die gerichtete Erre- gungsleitung, da eine Nervenzelle bei ausreichender Reizintensität an jeder Stelle erregt werden kann und Aktionspotentiale in jede Richtung leitet. Das Axon ist ein langer Fortsatz der Nervenzellen, der am Axonhügel entspringt. Es ist in der Regel mehr oder weniger stark verzweigt und mündet in synaptische Endigungen (Sy- napsen). Ein Axon kann je nach Typ der Nervenzelle von 1 µm bis 1 m und länger (!!) sein. Es weist eine Dicke von 0,5 bis 10 μm auf. Das Axon ist von mehreren aufeinander folgenden Myelinscheiden umhüllt. Das Axon ist zuständig für die Übertragung des Aktions- potentials einer Nervenzelle und leitet dieses zu den Synapsen und damit an andere Ner- venzellen weiter. Des Weiteren wandern die Stoffe, die im Soma gebildet werden (Neuro- transmitter, Enzyme), durch das Axon zur Synapse. Auch in umgekehrter Richtung, also von der Synapse in Richtung Zellkörper, findet Stofftransport statt. Dr. Garn Elektromagnetische Felder und der Mensch Seite 3.1.1
Die Synapse stellt eine Schnittstelle zwischen Neuronen dar, über die Informationen che- misch übertragen werden können, vgl. Bild 3.1/2. Am Ende des Axons befinden sich soge- nannte Endknöpfchen, welche die Kontaktstelle zwischen zwei Nervenzellen bilden. Die Sy- napse enthält die präsynaptische Membran der "Senderzelle", welche die Neurotransmitter nach einer vorausgegangenen Erregung in den synaptischen Spalt entleert. Ihr gegenüber liegt die postsynaptische Membran der "Empfängerzelle", welche mit Rezeptoren bestückt ist, an der die Neurotransmitter binden, um dort sogenannte Ionenkanäle zu öffnen. Zwi- schen den beiden Membranen liegt der etwa 30 nm breite synaptische Spalt, welcher mit einer plasmatischen Lösung aufgefüllt ist. Ein Neuron bildet 1 bis 200.000 Synap- sen mit anderen Neuronen. Das mensch- liche Gehirn weist insgesamt etwa eine Billiarde Synapsen auf. Synapsen von Nervenzellen verbinden sich auf diese Weise indirekt untereinander zu einem neuronalen Netzwerk. Bild 3.1/2: Chemische Synapse Elektrochemische Funktionsweise der Nervenzelle Im Inneren des Axons wie auch außerhalb der Membran existieren Anionen und Kationen. Sie bestimmen Ladung und elektrische Leitfähigkeit des Mediums. Die Biomembran des A- xons sorgt nun dafür, dass zwischen Innen und Außen verschiedene Konzentrationen der Ionen bestehen. Das bedeutet, dass an der Außenwand des Axons eine andere elektrische Ladung anliegt als innen („Polarisation“, Ruhepotential). Die ungleiche Verteilung der Ionen innerhalb und außerhalb der Zelle sowie selektive Per- meabilitäten der Membrankanäle erzeugen das Membranpotential: Während die Zelle viele K+-Ionen und wenige Na+- und Cl--Ionen enthält, sind außerhalb der Zelle die Na+- und Cl-- Konzentrationen hoch und die K+-Konzentrationen niedrig. Aufgrund der unterschiedlichen Ionenkonzentrationen diffundieren die in der Zelle hoch konzentrierten K+-Ionen durch die K+- Kanäle hinaus. Dabei entfernen sie positive Ladungen aus der Zelle, das Zellinnere wird ne- gativ aufgeladen. Dies wirkt der Diffusion der K+-Ionen entlang ihres Konzentrationsgradien- ten entgegen. Die negative Aufladung des Zellinneren kommt zum Stillstand, wenn sie den „Diffusionsdruck“ des Konzentrationsgradienten für K+ gerade aufhebt. Im Zustand des Fließgleichgewichts passieren auswärts und einwärts gleich viele K+-Ionen die Kanäle. Für Körpertemperatur (T = 310 K) und ein Verhältnis der Ionenkonzentrationen von [K+]i/[K+]a = 39 wird daraus z. B. das K+-Gleichgewichtspotential von -97 mV. Die Herstellung und Aufrechterhaltung dieses Potentials erfolgt durch die Natrium-Kalium-Pumpe. Eine Nervenzelle erhält ein Signal, indem Neurotransmitter an spezielle Rezeptoren in der postsynaptischen Membran in den Dendriten oder auch des Somas der zu erregenden Zelle anbinden. Ist die Erregung auf diese Weise übertragen, wird sie über die Dendriten an das Soma der Nervenzelle und von dort zum Axonhügel weitergeleitet. Jede der eingehenden Depolarisationen an den verschiedenen Synapsen der Nervenzelle verändert dabei das Membranpotential an der axonalen Membran. Je näher eine Synapse am Soma ansetzt, desto stärker ist ihr Einfluss auf die Nerven- zelle, je länger der Weg, den die Erregung zurücklegen muss, desto schwächer wird der Einfluss. Eine stärkere Reizung eines Dendriten (Fortsatzes vom Soma, der Ner- venzelle) resultiert also in einer stärkeren Depolarisierung. Nahezu gleichzeitig einlau- fende Reize addieren sich in ihrer Wirkung, was bedeutet, dass sich innerhalb der Zelle und am Axonhügel ein Erregungspotential aufbaut (Summation). Dr. Garn Elektromagnetische Felder und der Mensch Seite 3.1.2
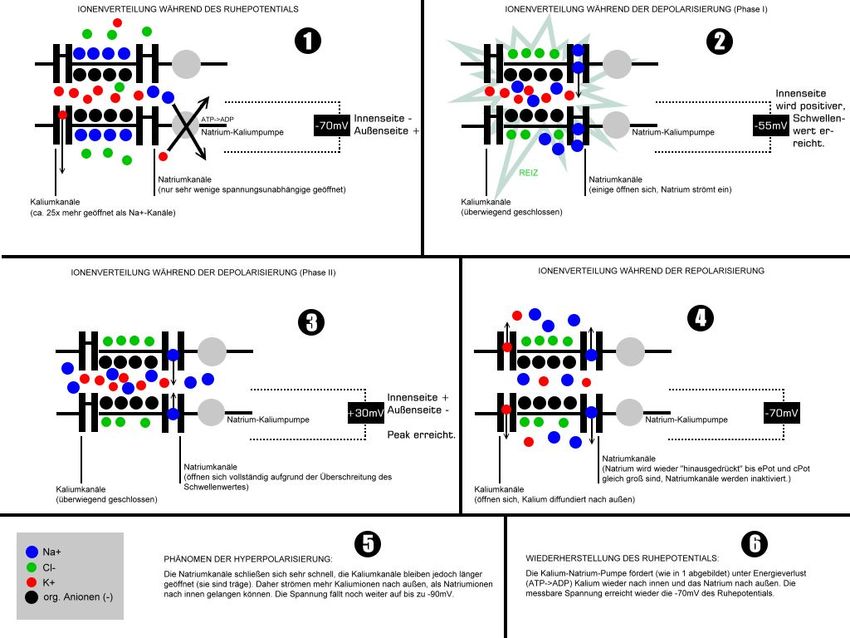
Wird am Axonhügel das Schwellenpotential erreicht und überschritten, so wird durch Depola- risation ein Aktionspotential entlang des Axons ausgelöst. Bild 3.1/3 zeigt den Verlauf der Ionenverteilung. Bild 3.1/3: Verlauf der Ionenverteilung während des Aktionspotentials Die spezifischen Ionenkanäle für Natrium- und Kalium-Ionen öffnen sich in Abhängigkeit vom Membranpotential, d. h. sie sind spannungsaktiviert. So ist beispielsweise ein spannungsab- hängiger Natriumkanal beim Ruhemembranpotential geschlossen und aktivierbar. Bei Depo- larisation über einen kanalspezifischen Wert wird der Kanal durchlässig für Ionen und geht in den Zustand offen über. Der Kanal bleibt aber trotz anhaltender Depolarisation nicht offen, sondern wird innerhalb weniger Millisekunden unabhängig vom Membranpotential wieder geschlossen. Das geschieht meist durch einen im Zytoplasma liegenden Teil des Kanalpro- teins, die Inaktivierungsdomäne, die sich gleich einem „Stöpsel“ in den Kanal setzt und die- sen verstopft. Nicht alle Kanäle öffnen sich gleichzeitig bei ein und demselben Wert des Membranpotentials. Vielmehr ist die Wahrscheinlichkeit eines Kanals, in einen bestimmten Zustand überzugehen, spannungsabhängig. Aus der rein statistischen Verteilung stellt sich ein Gleichgewicht ein, so dass eine größere Zahl von Kanälen in der Summe sehr gut das beschriebene Modell erfüllt. Auch ist der Zeitaufwand, um von einem Zustand in den anderen überzugehen, kanalspezi- fisch. Im geschilderten Natriumkanal läuft die Konformationsänderung von geschlossen nach offen in weniger als einer Millisekunde ab, während ein vergleichbarer Kaliumkanal Zeit in der Größenordnung von 10 ms benötigt. (Elektrisches Aktionspotential pflanzt sich entlang des Axons fort) Wandert nun ein Aktions- potential durch die Änderung des Konzentrationsgefälles der Ionen innerhalb des Axons am Axon entlang bis zum Endknöpfchen, so stößt dieser elektrische Impuls am Ende des Axons an eine Grenze, da eine Übertragung des elektrischen Signals durch den synaptischen Spalt zwischen den beiden Zellen nicht möglich ist. Der Reiz wird chemisch über die Synapsen weitergeleitet und mittels Neurotransmitter auf eine andere Zelle übertragen. Sobald ein Ak- Dr. Garn Elektromagnetische Felder und der Mensch Seite 3.1.2
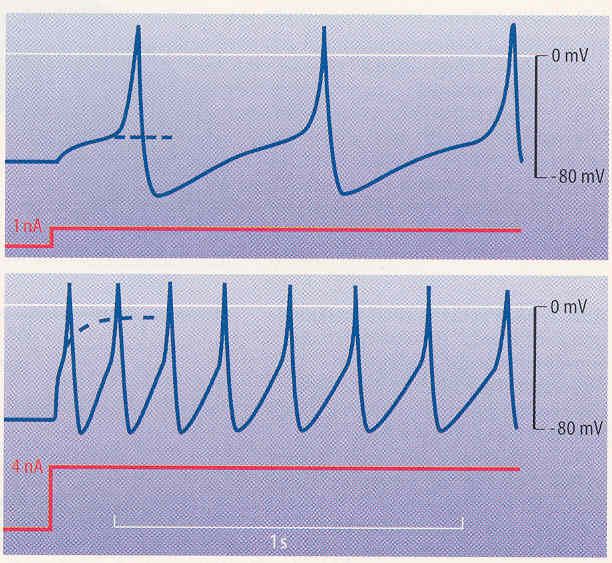
tionspotential ausgelöst wurde, braucht die Zelle Zeit (Refraktärphase), um das Membranpo- tential wiederaufzubauen (Repolarisation), vgl. Bild 3.1/4. Während der Refraktärphase kann kein neues Ak- tionspotential ausgelöst werden. Wenn also von nacheinander einlaufenden Reizen einer so stark ist, dass die Zelle ein Aktionspotential bildet und der nachfolgende Reiz während der Refraktärzeit einläuft, bildet die Zelle dafür kein neues Aktionspo- tential aus. Je mehr Aktionspotentiale die Zelle pro Sekunde abfeuert (je nach Zelltyp bis zu 500 Mal), desto stärker ist der Reiz. Bild 3.1/4: Verlauf des Aktionspotentials Signalcodierung („Puls-Zahl / Pulsfrequenz-Codierung“) Alle Informationen, die die Nerven übertragen sollen, müssen als Frequenz von Aktionspo- tentialen „codiert“ werden. Langdauernde Depolarisationen lösen Serien von Aktionspoten- tialen aus, in denen die Frequenz der Impulse zur Stärke der Depolarisation proportional ist. Bild 3.1/5 zeigt, wie eine Zelle auf das Einschalten eines Reizstromes von 1 nA oder von 4 nA antwortet. Der geringe Strom von 1 nA führt zu einer langsam ansteigenden, elektrotoni- schen Depolarisation, die in Fortsetzung der strichlierten Kurve ihren Endwert finden würde. Vor dem Einstellen des Endwertes erreicht jedoch die Depolarisation die Schwelle und löst ein Aktionspotential aus. Dieses hyperpolarisiert nach der Repolarisation über das Ruhepo- tential hinaus, dann folgt eine langsame Depolarisation, nach etwa 0,5 s wird die Schwelle erreicht und ein weiteres Aktionspotential ausgelöst. Dieser Zyklus kann sich wiederholen, solange der depolarisierende Strom fließt. Die Dauerdepolarisation wird somit in eine rhyth- mische Aktionspotentialauslösung mit etwa 2 Hz umgesetzt. Beim stärkeren Strom von etwa 4 nA erfolgt grundsätzlich das gleiche wie bei 1 nA, nur die Steilheit und die Amplitude der (strichlierten) Dauerdepolarisation sind größer und entspre- chend die Frequenz der erzeugten Aktionspotentiale höher: Sie liegt anfänglich bei 7 Hz und nimmt auf 4 Hz ab. Diese langsame Abnahme einer Frequenz bei gleichbleibendem Reiz- strom wird „Adaption“ genannt. Insgesamt ist also die Amplitude des Reizstromes bzw. der Depolarisation in entsprechende Aktionspotentialfrequenzen umcodiert worden. a) Der depolarisierende Strom von 1 nA in ein Neuron erzeugt ein elektrotonisches (durch gleichblei- benden Stromfluss ausgelöstes) Potential, das in eine Dauerdepolarisation von etwa – 20 mV münden würde (strichliert), wenn nicht die Schwelle zur Auslösung eines Aktionspotentials über- schritten würde. Die Aktionspotentiale wiederholen sich rhythmisch, solange der Stromfluss an- hält. b) Der Strom von 4 nA erzeugt ein elektrotonisches Potential, das fast 0 mV erreichen würde (strich- liert). Es wird jedoch eine Serie von Aktionspotentialen mit einer höheren Wiederholrate ausgelöst als bei 1 nA. Bild 3.1/5: Rhythmische Impulsbildung, ausgelöst durch einen andauernden Reizstrom Dr. Garn Elektromagnetische Felder und der Mensch Seite 3.1.3
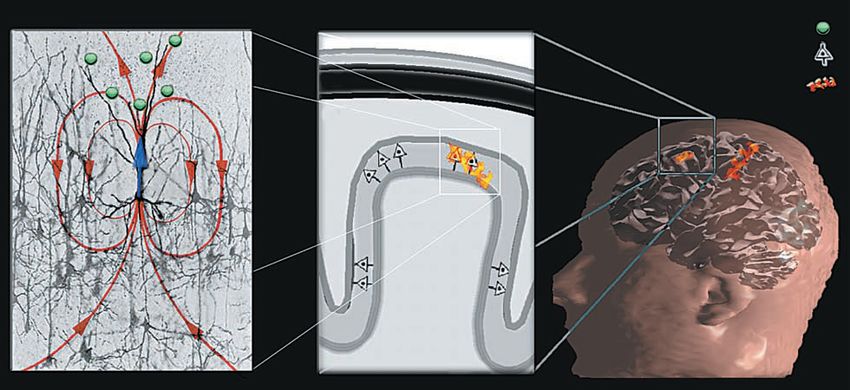
3.1.2 Elektrische und magnetische Felder im Gehirn Entstehung makroskopisch messbarer Felder In der Großhirnrinde (graue Masse) sitzen kortikale Pyramidenzellen (besonders große Ner- venzellen, die beim Menschen 85 % aller Nervenzellen ausmachen). Sie haben verlängerte Dendriten, die systematisch säulenförmig aneinander ausgerichtet sind und im rechten Win- kel zur Kortexschichte stehen, vgl. Bild 3.1/6 (Mitte). Diese räumliche Anordnung und die simultane Aktivierung großer Populationen dieser Zellen erzeugen die räumliche und zeitli- che Superposition der elementaren Aktivität jeder Zelle. Das Feld eines einzelnen Neurons ist zu schwach, um messbar zu sein. Die im EEG detektierten Signale stammen vielmehr aus Volumenbereichen mit untereinander verbundenen Neuronen, deren synaptische Strö- me zeitlich korreliert sind. Dies resultiert in einem Stromfluss und entsprechenden Potentia- len, die mit der Elektro-Enzephalographie (EEG) und der Magneto-Enzephalographie (MEG) gemessen werden können. Die kortikalen Nervenzellen bilden funktionale Netzwerke, die über verschiedene Gehirnregi- onen verteilt sein können. Sie sind die mutmaßlichen Generatoren der EEG u. MEG Signale. Stimulierende postsynaptische Potentiale entstehen am Scheitel des Dendritenbaums einer kortikalen Pyramidenzelle und triggern die Erzeugung eines Stromflusses durch das Gehirn- volumen von der nicht erregten Membran des Somas zu den Enden des Dendritenbaums. Der Strom fließt durch den Dendriten (Primärstrom im Bild 3.1/6 links, in blau) und verteilt durch den Kortex zurück (Sekundärströme, in rot). Bild 3.1/6: Netzwerke kortikaler Neuronengruppen als Erzeuger der EEG-Signale Näherungsweise Berechnung elektrischer und magnetischer Felder im Kopf Die gesamte Stromdichte im Kopf besteht aus zwei Komponenten: Einem primären (oder treibenden) Stromfluss JP(r´), der der tatsächlichen Aktivität der Neuronen entspricht, und dem im Volumen fließenden Strom JV(r´), der von der Wirkung des elektrischen Feldes auf die extrazellulären Ladungsträger herrührt. J(r´) = JP(r´) + JV(r´) = JP(r´) + s(r´).E(r´) = JP(r´) - s(r´).ÑV(r´), (3.1-1) wobei s … Leitfähigkeit des Gewebes im Kopf V … elektrisches Potential E = - grad V Die Divergenz des Gesamtstromes muss (im quasistatischen Fall, der hier gegeben ist) null sein (1. Maxwellgleichung mit ¶D/¶t = 0, div rot = 0 à div J = 0). Also DV = ÑJP/s (3.1-2) Die Lösung dieser Poissongleichung führt auf das elektrische Potential am Aufpunkt r. Inte- griert wird über das Volumen G, das die Quelle(n) enthält. Für den unendlich ausgedehnten, homogenen Raum würde (3.1-3) gelten: Dr. Garn Elektromagnetische Felder und der Mensch Seite 3.1.2
1 r - r´
4ps ò
V (r ) = J P
(r´) 3
dr´ (3.1-3)
G r - r´
Betrachten wir die Quellen, die sich in einem vergleichsweise kleinen Volumenbereich am
Ort r0 befinden. Der Beobachtungspunkt r ist ein Stück davon entfernt. Die primäre Strom-
dichte JP kann in diesem Fall gut durch einen äquivalenten „Stromdipol“ als Punktquelle re-
präsentiert werden.
JP(r´) = m.d(r´-r0), (3.1-4)
wobei d(r) … Diracfunktion
ò
Moment m º J (r´)dr´ .
P
(3.1-5)
Damit wird
1 r - r0
V (r ) = m (3.1-6)
4ps r - r0
3
(VGL.: PRECHTL KAP. 3.1)
Das inverse Problem führt auf die Berechnung der Dipolstromquellen, die die Potentiale an
der Kopfoberfläche erzeugen. Diese Potentiale werden im EEG gemessen.
Die Berechnung beinhaltet selbstverständlich Ungenauigkeiten:
Ø Die gemessenen Signale sind von Rauschen überlagert.
Ø Das Modell für die Kopplung zwischen den Stromquellen im Kortex und den gemessenen
Potentialen ist zu grob.
Ø Unterbestimmtheit bzw. Mehrdeutigkeit des inversen Problems bewirkt begrenzte Auflö-
sung in der Schätzung.
Dieses Verfahren wird zur Quellenanalyse eingesetzt. Es liefert sowohl in der Neurologie als
auch in der Gehirnforschung wertvolle Aufschlüsse über die Topographie der Gehirnaktivität.
Quellenanalyse (Low resolution brain electromagnetic tomography, LORETA)
Die primären Generatoren der im EEG registrierten Wellenvorgänge und Potentialschwan-
kungen sind synaptische Ströme. Diese Stromflüsse über die neuronalen Membranen fun-
gieren als winzige Stromquellen und –senken. Für jedes Neuron muss der gesamte Fluss
von Ladungen (hinein und hinaus) in Summe null ausmachen.
Bezogen auf die Schichtdicke haben die EEG-Messpunkte großen Abstand. Daher können
die Stromverteilungen innerhalb eines Stückes Kortex durch kleine, dipolförmige Stromquel-
len repräsentiert werden, die normal zur jeweiligen Kortexfläche ausgerichtet sind und deren
Stärke zeitlich variiert. Die Kopplung zwischen den Dipolstärken und den an der Kopfoberflä-
che gemessenen elektrischen Feldern kann damit einfach als Summe bzw. Integral über alle
Volumenbereiche berechnet werden. Mittels Quellenanalyse kann man aus den am Kortex
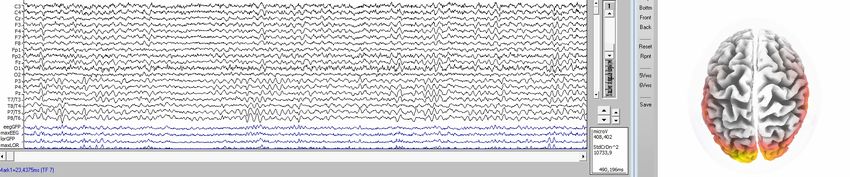
gemessenen Oberflächenpotentialen auf die Quellen zurückrechnen, Bild 3.1/7.
Bild 3.1/7: EEG-Aufnahme nach dem 10-20 System (Koordinatensystem der EEG-
Messelektroden-Punkte auf der Schädeldecke), LORETA-Quellenanalyse
Dr. Garn Elektromagnetische Felder und der Mensch Seite 3.1.3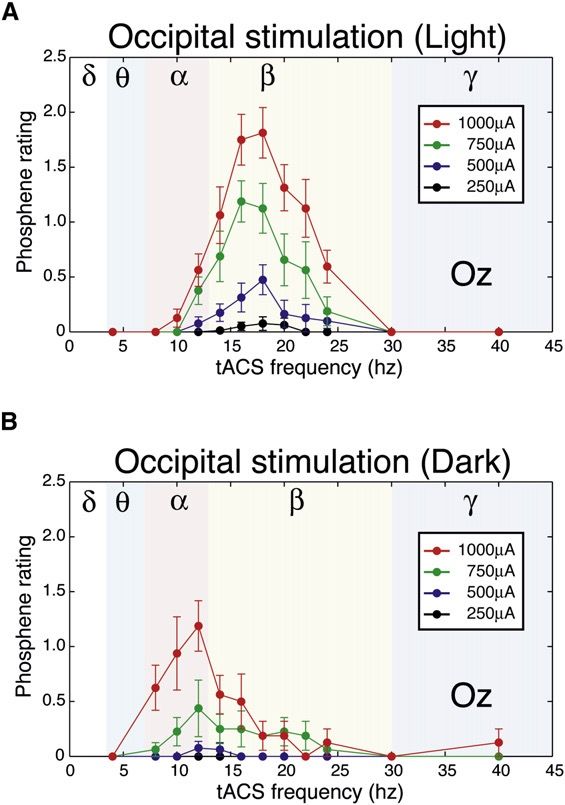
3.2 Nichtthermische Wirkungen niederfrequenter Felder
3.2.1 Wirkungen auf Nervenzellen
Reizwirkungsmodell
Bild 3.2/1 zeigt das Ersatzschaltbild der Zellmembran. Die über die Zellmembran fließende
Stromdichte Jm ist durch (3.2-1) gegeben. Die Leitwerte für die Ionen sind nichtlineare, kom-
plexe Funktionen, die aus Differentialgleichungen berechnet werden.
dU
J m = cm + ( J K + J Na + J L ) , (3.2-1)
dt
cm Membrankapazität
gNa, gK nichtlineare Leitwerte für Na+ und
K+
gL linearer Leitwert für den Leck-
strom
JK, JNa Ionenstromdichten;
JL Leckstromdichte;
EK, ENa, EL Nernstpotentiale.
Bild 3.2/1: Ersatzschaltbild der Zellmembran (nach Hodgkin-Huxley)
An der Zellmembran wirken hohe elektrische Kräfte: Bei einem Membranruhepotential von
- 90 mV und einer Membrandicke von etwa 10 nm beträgt die Spannung 9 kV/mm (!).
(Durchschlagsfestigkeit trockener Luft: 2-3 kV/mm; Glas: 10 kV/mm.) Die Leitfähigkeiten der
Membran für die Ionenströme sind eng mit dem elektrischen Feld verknüpft. Änderungen des
chemischen Ruhezustands bewirken Veränderungen der elektrischen Eigenschaften. Diese
Änderungen verursachen die funktionalen Reaktionen der Nerven und Muskeln.
Den Mechanismus der Reizwirkung elektrischer Ströme auf Nervenzellen kann man nach
dem obigen Modell von Hodgkin-Huxley darstellen. Wird durch das äußere Feld ein Strom
eingeprägt, der über die hochohmigen Zellmembran fließt, so entsteht dort eine polarisieren-
de oder depolarisierende Spannung, vgl. Bild 3.2/2. Erreicht eine depolarisierende Spannung
den Wert des so genannten Aktionspotentials, so wird der Nervenreiz ausgelöst. Im gegen-
teiligen Fall (zusätzliche Polarisierung, d.h. noch stärker negatives Potential im Zellinneren),
wird die Erregbarkeit der Nervenzelle vermindert.
a) Strom fließt durch lang gestreckte Nervenzelle b) Vereinfachtes elektrisches Ersatz-
schaltbild
Bild 3.2/2: Modell für die Reizwirkung auf Nervenzellen
Aus Bild 3.2/2b kann man einfache Gleichungen für den Zeitverlauf von Ladung und Span-
nung an der Membran sowie für die Zeitkonstante der Aufladung herleiten: Betrachten wir
einen Strom i(t), der zum Zeitpunkt t = 0 nach einer Stufenfunktion von der Amplitude 0 auf 1
geschaltet wird. Dann gilt
Dr. Garn Elektromagnetische Felder und der Mensch Seite 3.2.1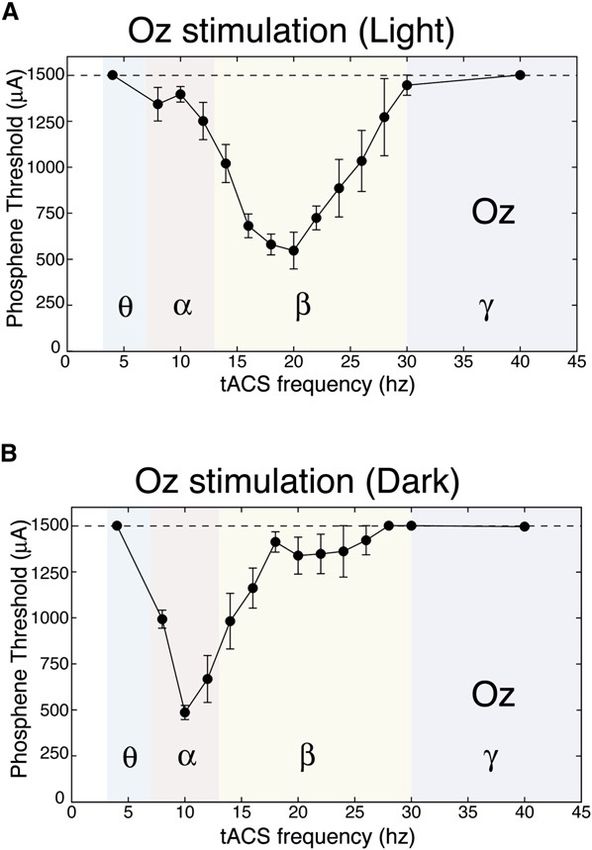
uM (t ) = i R (t ).R = I .R .( 1 - e
-t
M
) (3.2-2)
mit
tM = rm.cm .. Membran-Zeitkonstante
cm ………… Kapazität eines Membransegmentes
rm ……........ Widerstand eines Membransegmentes.
Für die Stromstärke IA eines Stromimpulses, der an der Membran innerhalb der Zeit t die
Spannung UA (Aktionspotential) erzeugen soll, gilt
UA
I A = R -t . (3.2-3)
1- e M
Für t ® ¥ wird IA = UA/R.
Die Ladung QA, die erforderlich ist, um an der Membran die Spannung UA zu erzeugen, ist
UA t
Q A = I A .t = . -t . (3.2-4)
R 1- e M
Bild 3.2/3 verdeutlicht den Zusammenhang zwischen Strom und Ladung an der Membran.
Weiters sieht man den Energieverbrauch zur Erzeugung einer Stimulation in Abhängigkeit
von der auf eine gewebespezifische Konstante normierten Impulsdauer.
Bild 3.2/3: Berechneter (normali-
sierter) Zusammenhang von
Strom, Ladung und Energiever-
brauch (Vertikalachse) für die Errei-
chung der Stimulationsschwelle an
einer Zellmembran bei Anregung
mit einem rechteckförmigen, uni-
polaren Stromimpuls,
(Horizontalachse: Pulsdauer auf die
Membranzeitkonstante normiert)
Kompliziertere Rechenmodelle existieren, liefern jedoch qualitativ gleiche Ergebnisse wie
das einfache Modell. Experimentell bestimmte Werte der Membran-Zeitkonstanten hän-
gen stark von der Gewebeart ab. Ein typischer Mittelwert für
periphere Nervenzellen ist 0,3 ms, für den Herzmuskel 3 ms.
Sehsinn, Gehör und Tastsinn benötigen kleine Zeitkonstanten für schnelle Reaktionen, der
Herzmuskel hingegen soll gerade nicht auf Kurzzeitereignisse reagieren und hat daher eine
größere Zeitkonstante!
Dr. Garn Elektromagnetische Felder und der Mensch Seite 3.2.2Schwellwerte der Stromstärke/Stromdichte bzw. Gewebefeldstärke für die Stimulation von
Nervenzellen bei Niederfrequenz und bei Impulsen („Wahrnehmungsschwellen“ und ihre
zeitlichen Abhängigkeiten)
Bei markscheidenhaltigen Nervenfasern im peripheren Nervensystem, zu Muskeln und zu
Sinneszellen, des Menschen liegt die Reizschwelle bei einer Gewebefeldstärke von 4-6 V/m.
Bei stärkerem Stimulus entstehen Unbehagen und dann Schmerz. Die niedrigste Schwelle
für eine nicht mehr tolerierbare Nervenreizung setzt man 20 % über dem Mittelwert der
Wahrnehmungsschwelle an (ICNIRP).
Für die Reizschwelle im Cortex werden für gepulste Felder etwa 10 V/m angegeben. Dies
bestimmt man durch transkranielle magnetische Stimulation.
Oberhalb von ca. 1 kHz steigen die Schwellwerte an, weil die Zeit, die für die Akkumulation
von Ladung zur Verfügung steht, immer kürzer wird. Unterhalb von ca. 10 Hz steigen sie
ebenfalls an, weil sich die Nervenzelle an den langsamen Depolarisationsstimulus anpasst.
Muskelzellen sind wesentlich unempfindlicher als Nervenzellen. Besondere Beachtung ver-
dient hier der Herzmuskel, bei dem jede anormale Funktion lebensgefährlich sein kann. Die
Fibrillationsschwellen liegen jedoch noch um zumindest einen Faktor 50 über den Reiz-
schwellen für Muskelfasern. Hier beginnen die Schwellen bereits oberhalb von ca. 120 Hz
anzusteigen, aufgrund der wesentlich längeren Zeitkonstanten der Muskelzellen im Vergleich
zu markscheidenhaltigen Nervenfasern.
Körperbewegungen werden vom Motorcortex (Zentrallappen) über motorische Nervenfasern
ausgelöst, die auf den Muskelfasern enden. Die Endplatten dieser Nerven übertragen Akti-
onspotentiale an die Muskelfasern. Die Kraftwirkung des Muskels wird durch die Bewegung
von Aktin- und Myosinfilamenten ausgelöst, die sich ineinanderschieben. Dabei bindet je-
weils ein Myosinkopf abwechselnd, so dass quasi „ein Fuß nach dem anderen“ vorgesetzt
wird. Die Bewegung erfolgt gerichtet, da Myosin auf dem Aktinfilament nur in eine Richtung
wandern kann. Die Motoraktivität wird durch den sogenannten Querbrückenzyklus beschrie-
ben, vgl. Bild 3.2/4.
Phase 1 Phase 2 Phase 3 Phase 4
Myosin bindet an Aktin Myosinköpfchen spalten Myosinköpfchen lösen Myosin im Ruhezustand
ATP zu ADP und Phos- sich unter Aufnahme von
phat und führen dabei ATP vom Aktin.
ihren Kraftschlag aus.
ATP: Adenosintriphosphat, Energieträger in Zellen
ADP: Adenosindiphosphat entsteht durch Hydrolyse aus ATP
Bild 3.2/4: Querbrückenzyklus (Myosin: gelb; Aktin: rosa)
Effekte, die unterhalb der Stimulationsschwelle auftreten
Sowohl bei in-vitro (außerhalb eines lebenden Organismus, „im Reagenzglas“) als auch bei
in-vivo (in einem lebenden Organismus) Experimenten findet man Effekte, die unterhalb der
Stimulationsschwellen auftreten. Für die inhibitorische Wirkung einer polarisierenden Gewe-
befeldstärke ist dies evident. Die Empfindlichkeit von Gehirnzellen des Hippocampus wurde
z.B. einem in-vitro Experiment gemessen. Abhängig von der polarisierenden Gewebefeld-
stärke veränderte sich die Spikeamplitude bei Reizung mit 25 bis 250 ms langen Pulsen, vgl.
Bild 3.2/5.
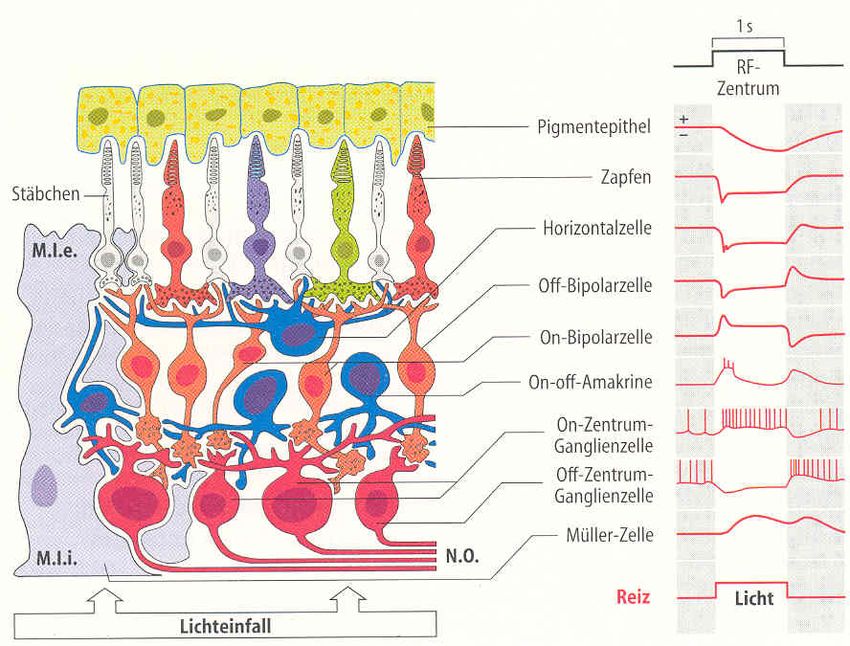
Dr. Garn Elektromagnetische Felder und der Mensch Seite 3.2.3Bild 3.2/5: Änderung der Spikeamplitude bei Gehirnzellen (Hippocampus) in Abhängigkeit von der Gewebefeldstärke (in-vitro Expe- riment). Dies entspricht einer „Arbeits- punkt-Verschiebung“ im Sinn einer IN/OUT-Kennlinie. Ein in-vivo nachgewiesener Effekt, der auch unterhalb der genannten Reizschwellen auftritt, ist die Induktion sogenannter Phosphene. Er bewirkt die Wahrnehmung flackernder Lichter- scheinungen an der Peripherie des Sehfeldes. Die minimale Flussdichte zur Auslösung von Magnetophosphenen beträgt bei 20 Hz ca. 5 mT und steigt sowohl bei niedrigeren als auch bei höheren Frequenzen an. Man nimmt an, dass das elektrische Feld hier direkt die Ner- venzellen in der Retina anregt. Die entsprechenden elektrischen Feldstärken liegen bei ca. 50-100 mV/m bei 20 Hz. Bild 3.2/6 zeigt den Aufbau der Netzhaut. Sie besteht aus fünf Schichten hoch spezialisierter Nervenzellen. Die Photosensoren sind die Stäbchen und Zäpfchen: Ø Stäbchen: Durchmesser 3 µm; 120 Millionen; sehr lichtempfindlich (1 Photon à 1 mV), Absorptionsmaximum bei 500 nm (à Sehen bei Mondlicht) Ø Zapfen: Durchmesser: 2 µm; 6 Millionen; rot-grün-blau; brauchen 200 Photonen Bild 3.2/6: Photosensoren in der Retina und Schema der Reaktion einzelner Neurone Bild 3.2/7a zeigt Ergebnisse eines Experimentes, in dem bei acht Versuchspersonen mittels Elektroden am Hinterkopf („transcranial alternating current stimulation“, tACS) Phosphene ausgelöst wurden. Beim stärksten Stimulationsstrom von 1000 µA betrug das Maximum der Stromdichte unter der Stimulationselektrode 83 µA/cm2. Das Intensitätsmaximum der Wahr- nehmungen trat in der Dunkelheit bei ca. 12 Hz auf, in beleuchteter Umgebung bei ca. 18 Hz. Bild 3.2/7b zeigt die Wahrnehmungsschwellen. Bild 3.2/8 gibt die von den Versuchsperson berichteten Wahrnehmungen wieder (Phosphene in grau). Dr. Garn Elektromagnetische Felder und der Mensch Seite 3.2.4
Bild 3.2/7: a) Frequenzverlauf der Phosphene b) Stimulationsschwellen für Phosphene Bild 3.2/8: Zeichnungen der von sechs Versuchspersonen wahrgenommenen Phosphene Bild 3.2/9 zeigt mittlere Schwellwerte für Stimulation durch Magnetfelder. Es wird Ganzkör- perexposition in einem konstanten Feld angenommen. Während Nervenzellen eine Depolari- sation von etwa 15 bis 20 mV benötigen, um ein Aktionspotential zu erzeugen, können sy- naptische Prozesse durch Änderung des präsynaptischen Membranpotentials bereits bei weniger als 1 mV ausgelöst werden. Relativ geringe Änderungen des präsynaptischen Po- tentials können damit eine prozentuell wesentlich stärkere Änderung des postsynaptischen Potentials verursachen. Die postsynaptische Zelle summiert die präsynaptischen Inputs meh- rerer Zellen. Hier kann sowohl eine Erregung als auch eine Inhibition (Hemmung) entstehen. Die Synapsen sind damit die empfindlichsten Punkte im Nervensystem. Dr. Garn Elektromagnetische Felder und der Mensch Seite 3.2.5
Bild 3.2/9: Mittlere Stimulationsschwellen bei Magnetfeld-Exposition. Horizontal: Frequenz der Stimulation in Hz, Vertikal: magnetische Flussdichte in mT (Effektivwert) MPE: Maximum Permissible Exposure (“general public” entspricht “Allgemeinbevölkerung”, “con- trolled environment” entspricht „berufliche Exposition“, dazwischen Schutzfaktor 5 ) Bild 3.2/10 gibt eine Übersicht der Stromdichte-Schwellwerte für verschiedene Wirkungen an Nerven- und Muskelzellen als Funktion der Frequenz. Die Mehrzahl der Stromdichte-Daten stammt aus Tierversuchen. Vor allem hinsichtlich des Frequenzverlaufes ist die Umlegung der Ergebnisse auf den Menschen mit Unsicherheiten behaftet. Die gezeigten Daten der Fib- rillationsschwellen beinhalten eine Unsicherheit um den Faktor 2 - 3. Bei herzkranken Perso- nen können auch bei niedrigeren Stromdichten bereits Störungen der Herztätigkeit auftreten. Das Bild stammt aus 1983, aber es zeigt recht übersichtlich die Größenordnungen und Fre- quenzabhängigkeiten. Zentrale Frage im Hinblick auf die Festlegung von Grenzwerten: welche äußeren Feld- stärken oder welche äußeren Ströme sind notwendig um im Inneren des Gewebes die Stromdichten in Bild 3.2/ zu erzeugen, bzw. entsprechende Gewebe-Feldstärken im Körperinneren? Dr. Garn Elektromagnetische Felder und der Mensch Seite 3.2.6
1µA/cm2 = 10 mA/m2
Bild 3.2/10: Schwellenwerte der elektrischen Stromdichte für verschiedene Wirkungen an
Nerven- und Muskelzellen
a1, a2 ….. Stimulation von Sinnesrezeptoren mit Elektroden (Wahrnehmung durch den Pro-
banden)
A ...…..... Alle Kurven oberhalb von Kurve A hängen mit der Erzeugung von Aktionspo-
tentialen zusammen, d. h. mit der Erzeugung von Reizwirkungen an Nerven
oder Muskeln durch elektrische Feldstärken bzw. Stromdichten im Körper
b1, b2-b5 . Störung der Herzaktion
b6 ……… Schwellwerte für Kammerflimmern (300 µA/cm2 bei 50 Hz)
b7 ……… Auslösung von Extrasystolen (80 µA/cm2 bei 50 Hz)
c1, c2 .…. Stimulation von Einzelzellen im Gehirn (Messungen mit Mikroelektroden)
d ………. Reizschwelle für die Nerv-Muskel Erregung nach der Reiztheorie von Schäfer, be-
zogen auf 50 µA/cm2 bei 50 Hz
e ………. Erzeugung von Membranspannungen von 10 mV durch elektrische Felder in der
Umgebung von Nervenzellen
g ………. Erzeugung von Membranspannungen von 0,1 mV durch elektrische Felder in der
Umgebung von Nervenzellen
h ..…….. Erzeugung von Phosphenen (Lichterscheinungen) im Auge durch Reizung mittels
Elektroden
i ……….. Flimmererscheinungen im Magnetfeld
j ……….. Änderung von Reaktionspotentialen durch Magnetfelder
EEG …... Aus EEG - Signalen berechnete, natürliche mittlere Stromdichten zufolge der
elektrischen Aktivität des Gehirns selbst
Zusammenfassung: Oberhalb von Kurve A: Bereich möglicher Schädigung bei Stromdicht-
en größer als ca. 20 bis 30µA/cm2 für Frequenzen bis 50 Hz.
Unterhalb von Kurve B: Sicherer Bereich
Dr. Garn Elektromagnetische Felder und der Mensch Seite 3.2.73.2.2 Wahrnehmung von Wirkungen an der Körperoberfläche
Wahrnehmung niederfrequenter elektrischer Felder
Für niedrige Frequenzen kann die Körperoberfläche als elektrisch gut leitend angesehen
werden. Sie hat daher in guter Näherung überall Erdpotential. An der Körperoberfläche treten
Feldüberhöhungen von bis zu einem Faktor 20 auf.
Das elektrische Feld bewirkt eine mit der Frequenz wechselnde Aufladung der relativ
hochohmigen Behaarung, so dass zwischen den Haaren und der Hautoberfläche Kräfte
wirksam werden und eine Vibration des Haarschaftes anregen. Diese Vibration wird über die
Haarwurzel und die Haarzwiebel zu den Haarfollikeln hin übertragen, wo Berührungs- und
Vibrationsrezeptoren die Bewegung registrieren können, vgl. Bild 3.2/11. Das Haar wird mit
der doppelten Feldfrequenz aus seiner Ruhelage abgelenkt.
Die Wahrnehmungen sind daher:
- Haarvibrationen
- "Kribbeln" der Haut.
Bild 3.2/11: Vibration der Haare im
elektrischen Wechsel-
feld
Bei Untersuchungen an Personen, die mit erhobener Hand unter einer Hochspannungslei-
tung stehen, konnte bei 5 % der Probanden eine Wahrnehmung des Feldes ab 1 kV/m fest-
gestellt werden. Eine belästigende Wahrnehmung setzt bei 1 % der Probanden bei erhobe-
nem Arm ab 5,5 kV/m ein, vgl. Bild 3.2/12. Sticheln auf der Haut wird ab 7 kV/m, vibrierendes
Kopfhaar ab 9 kV/m als belästigend empfunden. Unterschiedliche Wetterbedingungen führen
zu keinen signifikanten Änderungen bei den Wahrnehmungen.
Die Frequenzabhängigkeit der Wahrnehmungseffekte ist in Bild 3.2/13 dargestellt.
Bild 3.2/12: Wahrnehmung elektri-
scher Felder unter Hoch-
spannungsleitungen in
1m Höhe
Dr. Garn Elektromagnetische Felder und der Mensch Seite 3.2.8Bild 3.2/13: Mittlere Wahrneh-
mungsschwellen für
Haarvibrationen als
Funktionen der Fre-
quenz
Feldstärke von ca. 10kV/m mit:
220kV-Leitung in 20m über
Grund, oder 110kV-Leitung in
10m über Grund oder (fiktiv)
11kV in ca. 1m über Grund
Experimentelle Ergebnisse verschiedener Autoren stimmen relativ gut überein. Man findet
stets Unterschiede zwischen den Schwellwerten für Männer, Frauen und Kinder. Allerdings
ist es schwierig, Reizschwellen für die Versuchspersonen exakt zu definieren. Weiters be-
stehen bei Versuchen unter Hochspannungsleitungen Täuschungsmöglichkeiten durch Au-
ßentemperatur, Wind und den Höreffekt ("Surren" der Leitung, d.s. Spitzenentladungen nahe
am Leitungskabel wo die Feldstärke am höchsten ist).
Elektrisierungen bei Berührungen von elektrisch leitenden Körpern in elmagn. Feldern
Große, elektrisch leitende Körper, die sich in einem elektromagnetischen Feld befinden, kön-
nen gegenüber Erde Spannung führen. Berührt man einen solchen Leiter, so können durch
die gegen Erde fließenden Ableitströme spürbare Elektrisierungen auftreten. Von besonders
sensiblen Personen können niederfrequente Ströme bei Dauerkontakt bereits ab 200 µA
wahrgenommen werden. Der Prozentsatz solcher "elektrosensibler" Personen könnte neue-
ren Untersuchungen zufolge 1 - 2 % der Bevölkerung ausmachen. Im allgemeinen wird an-
genommen, dass für die Durchschnittsbevölkerung die Wahrnehmungsgrenze im Frequenz-
bereich von 10 Hz - 100 Hz über 0,5 mA liegt. Bei IEC wird die 50 % - Wahrnehmungsgrenze
mit 1 mA angenommen.
Von Erde isolierte, elektrische Leiter, die bei Berührung Elektrisierungen auslösen können,
sind z. B. Dachrinnen, Baugeräte, PKW oder größere Kfz, Werkzeuge oder Weidezäune. Es
erfolgt zunächst ein kurzzeitiger Entladeimpuls, der unmittelbar bei der Berührung auftritt.
Anschließend fließt ein Dauerstrom, der bei weiter bestehen bleibendem Kontakt aus dem
Leiter über die Person gegen Erde fließt.
Ein Dauerstrom von 1 mA bei 50 Hz fließt durch eine aufrecht stehende Person, wenn die
ungestörte elektrische Feldstärke
Ø 70 kV/m beträgt, oder
Ø 9 bis ca. 11 kV/m beträgt und die Person z. B. einen im Feld stehenden PKW berührt,
vgl. Bild 3.2/14a.
In einer isolierten, plattenförmigen Anordnung entsteht ein Dauerstrom von etwa 3 µA/m2 pro
kV/m, d. h. 1 mA entsteht bei ca. 300 m2.kV/m.
Entladung einer Ladungsmenge von 0,5 µC:
Ø Isolierte Person in einem Feld von 10 kV/m entlädt sich über einen geerdeten Gegen-
stand
Ø Geerdete Person berührt einen isolierten Leiter von 40 m2 Fläche, Feldstärke 1 kV/m,
vgl. Bild 3.2/14b).
Dr. Garn Elektromagnetische Felder und der Mensch Seite 3.2.9a) Dauerentladung durch eine Person: Erforderliche ungestörte Feldstärke für einen gerade
spürbaren Dauerstrom von 1 mA
b) Impulsentladung durch eine Person: Erforderliche ungestörte Feldstarke für einen gera-
de spürbaren Entladungsimpuls von 0,5 µC
Bild 3.2/14: Skizzen von Situationen, in denen spürbare Elektrisierungen bei 50 Hz auftreten
können
Man erkennt aus den in Bild 3.2/14 angegebenen Werten, dass die Feldstärke bzw. die Auf-
fangfläche, die notwendig ist, um einen spürbaren Effekt in einem Feld zu erzeugen, für Mik-
roschocks (spürbare Entladungen) um etwa eine Größenordnung geringer ist, als für einen
spürbaren Dauerstrom.
Ähnliche Entladungsimpulse können auch infolge elektrostatischer Ladungstrennung auftre-
ten. Zum Beispiel kann sich eine Person mit etwa 100 pF Kapazität beim Aufstehen aus ei-
nem Kfz-Sitz aus Kunststoff auf etwa 10 kV aufladen, entsprechend einer Ladungsmenge
von 1 µC.
Wirkungen niederfrequenter Ableitströme – Basis für Schutzkonzepte
Elektrische Ströme können je nach Stromstärke und Frequenz verschiedene Reaktionen
auslösen, die von einer Wahrnehmung des Stromes durch ein Gefühl der Wärme (Gleich-
strom) oder durch ein Prickeln (Wechselstrom) bis zu zwanghaften Muskelkontraktionen und
darüber zu Atemlähmungen und Kammerflimmern reichen.
Tabelle 3.2/1 enthält eine Zusammenstellung der Stromwirkungen, geordnet nach steigender
Stromstärke, für 60 Hz. Werte für 50 Hz unterscheiden sich davon nur geringfügig. Die Werte
gelten für Männer, die Schwellen für Frauen liegen etwa bei 2/3 der angegebenen Werte.
Effektivwert der Reaktion/Schwelle Prozentsatz der Probanden
Stromstärke (mA) (männliche Erwachsene)
0,13 Wahmehmung bei Berührung 1%
0,36 -"- 50 %
0,49 Wahrnehmung bei Griffkontakt 1%
1,10 -"- 50 %
9 Unwillkürliche Muskelkontraktionen (Loslass- 0,5 %
strom)
16 -"- 50 %
23 Atemschwierigkeiten 50 %
100 Kammerflimmern (Erwachsene mit 70 kg, 0,5 %
Einwirkdauer 3 Sek.)
Tab. 3.2/1: Reaktionsschwellen für 60 Hz-Ströme, vergleiche mit den Auslöseströmen von
Fehlerstromschutzschaltern /FI-Schalter)
Dr. Garn Elektromagnetische Felder und der Mensch Seite 3.2.10Für Kinder gelten noch niedrigere Werte, z. B. 4,5 mA als 0,5 % Schwelle für die Loslass-
stromstärke und 35 mA als 0,5 % Schwelle für Kammerflimmern (Kinder mit 20 kg).
Gleichströme werden erst bei höheren Stromstärken wahrgenommen (1 mA: 1 %; 5,2 mA:
40 %, Griffkontakt) als 50 (60) Hz Ströme. Für höhere Frequenzen steigen die Schwellwerte
ebenfalls an. Ströme, die zu einer möglicherweise gefährlichen Schreckreaktion bei der Be-
rührung von Metallteilen führen können, liegen zwischen 2,2 mA und 3,2 mA (50 % Frauen)
je nachdem, wie der Kontakt hergestellt wird. Diese Stromstärken stellen bereits eine gewis-
se Gefährdung dar. Kontinuierliche Ströme (60 Hz) werden ab 2 mA als unangenehm emp-
funden.
Wird die Stromstärke weiter gesteigert, so erreicht man die Schwelle der Loslassströme. Dort
kommt es zu unwillkürlichen Muskelkontraktionen, sodass ein einmal gefasster, Strom füh-
render Teil nicht mehr ausgelassen werden kann. Loslassströme stellen an sich noch keine
unmittelbare Gefahr dar, jedoch kommt es bei lang anhaltender Einwirkung von Strömen
über der Loslass- (let-go-) Schwelle zu Atmungsschwierigkeiten und schließlich zum Ersti-
ckungstod. Daher müssen Loslassströme bei langer Exposition als lebensgefährlich angese-
hen werden.
Die Loslassströme sind ähnlich wie die Wahrnehmungsschwellen frequenzabhängig und er-
reichen die niedrigsten Werte zwischen etwa 15 Hz und 300 Hz. Die 1 % Schwelle für
Gleichstrom liegt bei 60 mA, bei 74 mA sprechen schon 50 % der Probanden an.
Bei noch höheren Stromstärken werden Stromdichten erreicht, bei denen das Herz zu unko-
ordinierten Kontraktionen angeregt wird, so dass die Pumptätigkeit völlig ausbleibt. Wenn
dieses so genannte Kammerflimmern (Fibrillation) einmal eingesetzt hat, kehrt das Herz nur
selten von selbst in den normalen Rhythmus zurück. Erfolgt keine künstliche Defibrillation, so
tritt der Tod ein.
Aus Tierversuchen kann man schließen, dass die Fibrillationsschwellen dem Gewicht der
Tiere proportional sind und daraus entsprechende Werte für den Menschen ermitteln.
Bild 3.2/15: Loslassströme als
Funktion der Fre-
quenz
Wahrnehmung von Funkenüberschlägen
Greift eine Person auf ein elektrisch geladenes Objekt in einem niederfrequenten elektri-
schen Feld, so kommt es bereits vor dem Kontakt zu einem Funkenüberschlag. Die Entla-
dung wird ausgelöst, sobald in einem schmalen Spalt zwischen Finger und Leiteroberfläche
die elektrische Durchschlagfeldstarke überschritten wird. Diese Funkenüberschläge werden
wahrgenommen, selbst wenn sie einem Strom entsprechen, der weit unter der Empfindungs-
schwelle für Dauerkontaktströme liegt, da die lokale Stromdichte auf der Haut hohe Werte
annimmt. Berührt man z. B. den Türgriff eines kleinen PKW (Objektkapazität ca. 700 pF), so
Dr. Garn Elektromagnetische Felder und der Mensch Seite 3.2.11spürt man die Entladung ab ca. 600 V. Die Wahrnehmungsschwelle hängt auch von Lufttem-
peratur und Luftfeuchtigkeit ab und ist bei trockenem Wetter am höchsten. Die Schwellwerte,
bei denen die Empfindungen beginnen, unangenehm zu werden, liegen um einen Faktor 2 -
4 über den Schwellwerten für die Wahrnehmung.
Berührungsströme in Hochfrequenzfeldern
Auch bei Berührung elektrisch leitender Gegenstände, die sich in einem Hochfrequenzfeld
befinden, können Ströme über den Körper gegen Erde fließen. Diese Ströme können je nach
Stromstärke Schreckreaktionen, Schocks oder Verbrennungen hervorrufen. Bild 3.2/16 zeigt
experimentell ermittelte Wahrnehmungsschwellen für Berührungsströme, die heute für die
Grenzwertfestsetzung herangezogen werden.
Bild 3.2/16: Wahrnehmungs-
schwellen für
Berührungsströme im
Frequenzbereich von
10 kHz bis 10 MHz
2019 Fachvertiefung: Elektromagnetische Felder und Wellen Seite 1.2Sie können auch lesen

















































