Einfluss warmer Frühjahre auf die Ei-Größe von - Parus major
←
→
Transkription von Seiteninhalten
Wenn Ihr Browser die Seite nicht korrekt rendert, bitte, lesen Sie den Inhalt der Seite unten

Einfluss warmer Frühjahre auf die Ei-Größe von
Parus major
Lena Gonther MS-OE-VÖK SoSe 2020
Einleitung Einleitung
Methoden Der Klimawandel führt in Deutschland zu einem langfristigen Temperaturanstieg im Frühjahr
Ergebnisse und Sommer, welcher eine Verschiebung der phänologischen Phasen in Pflanzen zur Folge
hat.3,9 Diese phänologischen Phasen haben Auswirkungen auf den Aktivitätszyklus von
Diskussion
Schmetterlingslarven und anderen Insekten, die für die Kohlmeise Parus major ab März zur
Brutzeit die wichtigste Nahrungsquelle darstellen.11 Bei P. major kann, neben einer
genetischen Veranlagung, das Gewicht des Weibchens in der Zeit vor und während der
Eiablage die Größe der Eier positiv beeinflussen, da schwerere Weibchen mehr Energie in die
Eiproduktion investieren können.6 Relativ warme Frühjahre könnten daher zu besonders
guten Umweltbedingungen durch eine hohe Nahrungsverfügbarkeit für Weibchen und somit
größeren Eier führen in Gelegen führen (Abb.2).
Abb. 1: Gelege von P. major.
Hypothese
Ei- Fitness/Gewicht Temperatur
Die Frühjahre 2014 und 2018 gehören zu den Wärmsten seit
des Weibchens Im Frühjahr
Beginn der Wetteraufzeichnungen.3 Die Eier in diesen Jahren sind Größe
aufgrund möglicher erhöhter Nahrungsverfügbarkeit und damit
einer potentiell gesteigerten Fitness der Weibchen größer, als in
Abb. 2: Schemadarstellung. Die Frühjahrstemperatur beeinflusst
den anderen untersuchten Jahre.
das Gewicht der Weibchen und wirkt dadurch indirekt auf die Ei-
Größe.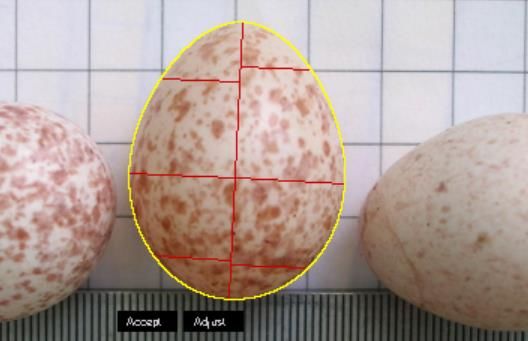
Einfluss warmer Frühjahre auf die Ei-Größe von
Parus major
Lena Gonther MS-OE-VÖK SoSe 2020
Einleitung Datenaufnahme
Methoden • Untersuchte Art : Kohlmeise Parus major
• Untersuchte Standorte wurden in drei Habitate aufgeteilt: Stadt (in Gießen),
Ergebnisse
Wiese (nahe Gießen) und Wald (wurde bei der Analyse nicht berücksichtigt da
Diskussion keine Daten für 2014 vorlagen)
• Datenaufnahme in den Jahren 2013 bis 2019
• Regelmäßige Untersuchung der Nistkästen und Fotodokumentation der Eier
• Vermessung der Eier digital mit dem Programm Image J, unter Verwendung des
Plug-In‘s Eggtool: Mithilfe von Egg Measurement wurde Volumen, Breite, Länge,
und Ei-Form bestimmt (Abb.3)
• Als Parameter für die Ei-Größe wurde das Volumen in cm³ verwendet
• Zur Einordnung der Ergebnisse Verwendung von Wärmesummen Temperatur
der des 1.März bis 20.April für die untersuchten Jahre von Wetterstation Linden
Abb. 3: Vermessung von Eiern mit ImageJ und Eggtool
Statistische Analyse I Statistische Analyse II
• Deskriptive Statistik: Prüfung der Normalverteilung des Ei-Volumens durch One- • Da auch die Habitate und die Gelegegröße1 das Ei-
sample Kolmonogorov-Smirnoff-Test, Bestimmung des Mittelwertes und der Volumen beeinflussen könnten, wurde ein
Standardabweichung generalisiertes lineares Modell für normalverteilte Daten
• Induktive Statistik: Unterschiede zwischen den Jahren für Habitate Stadt und Wiese ( und ein lineares Modell für normalverteilte Daten
hier ohne 2013 da hier keine Daten vorlagen) zusammen und getrennt geprüft erstellt und Effekte der Habitate, der Gelegegröße und
• Nicht normalverteilte Daten wurden mithilfe des Wilcoxon-Test geprüft. Dafür aller Jahre auf das Volumen überprüft (Anova und chi2
wurden die Unterschiede des Volumens der Eier der Jahre 2014 und 2018 mit Test)
wilcox.test(x,y, alternative =„greater“), dabei x = Ei-Volumen der Jahre 2014 oder • Durchführung der statistischen Analyse mit R Studio12
2018 und y = Ei-Volumen der restlichen Jahre, mit den Volumen in anderen Jahren Berechnung der Wiederholbarkeit r nach LESSELS & BOAG
(2013 – 2019) einzeln untersucht, um die Volumen für 2014 und 2018 als größer zu (1987) 8 für das Ei-Volumen
testen, für normalverteilte Daten das gleiche Vorgehen mit Student‘s t-Test
Einfluss warmer Frühjahre auf die Ei-Größe von
Parus major
Lena Gonther MS-OE-VÖK SoSe 2020
Einleitung Habitate zusammengefasst
Methoden Die Wiederholbarkeit der Messungen des Eivolumens lag bei 97,8%
Ergebnisse (r = 0.9778).
Das mittlere Eivolumen in den Untersuchungsgebieten betrug
Diskussion
insgesamt 2.16 ± 0.17 cm³ (Range 0.9 – 4.37) (Abb. 4). Die Daten
zeigten eine Normalverteilung ( p = 0.058, D = 0.069).
Abb. 5: Mittleres Eivolumen und Standardabweichung für die
Habitate Stadt und Wiese in den Jahren 2013 bis 2019
Habitate Stadt und Wiese
Da ein lineares Modell für beide Habitate zeigte, dass es einen
signifikanten Unterschied der Volumina zwischen Jahren und
Habitaten gab (lm( Volume ~ Jahr*Habitat) Zielvariable: Volumen,
Effekte des Prädiktors Jahr*Habitat F(1,366) = 26.92, p < 0.001) ,
wurde die Hypothese für beide Habitate im weiteren Verlauf
getrennt untersucht. Im Stadthabitat lag das mittlere Eivolumen bei
Abb. 4: Verteilung der des Eivolumens der Jahre 2013 bis 2019 2.19 ± 0.3 cm³ (Range = 0.9 – 4.37)(Abb.5). Im Habitat Wiese betrug
für die Habitate Stadt und Wiese zusammengefasst mit Median, das mittlere Eivolumen 2.16 ± 0.26 cm³ (Range = 1.2 – 4.18)(Abb.5).
Standardabweichung und Ausreißern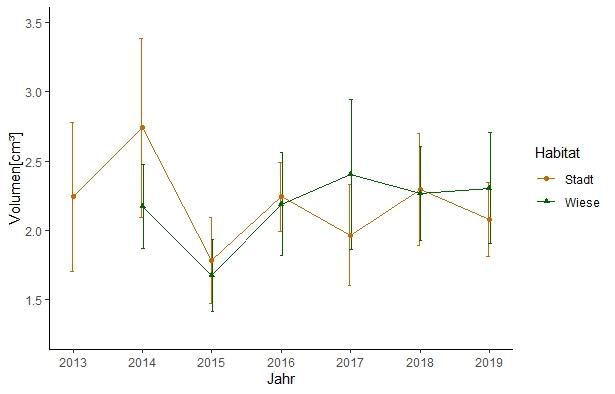
Einfluss warmer Frühjahre auf die Ei-Größe von
Parus major
Lena Gonther MS-OE-VÖK SoSe 2020
Einleitung Stadtgebiet Tab 2: Statistische Parameter t, df und p des Vergleich des Eivolumens der Jahre
2014 und 2018 mit den Jahren 2015 bis 2019 (t.test( y, x, alternative = „greater“))
Methoden Die Daten des Stadtgebietes wiesen eine Habitat:
Ergebnisse Normalverteilung auf (p = 0.02, D = 0.11). Die Eier aus Wiese Vergleich Vergleich
2014 2018
dem Jahr 2014 waren im Vergleich zu allen anderen
Diskussion
Jahren signifikant größer (wilcox.test(Volumen2014, x, Jahr t df p-Wert t df p-Wert
alternative = „greater“) siehe Tab. 1). Im Jahr 2018 2015 5.69 29.82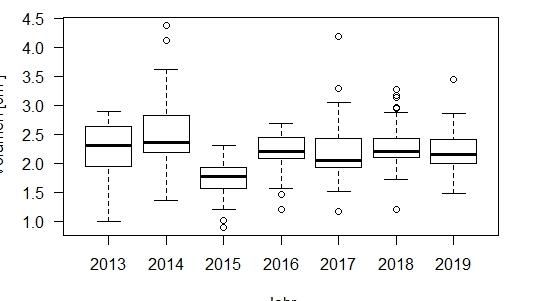
Einfluss warmer Frühjahre auf die Ei-Größe von
Parus major
Lena Gonther MS-OE-VÖK SoSe 2020
Einleitung I. Vorteil der Eigröße III. Einordnung der Ergebnisse
Methoden Für Weibchen kann es sich lohnen in besonders guten Jahren, in denen sie genug Da urbane Habitate vermutlich durch geringeres Invertebraten-
Energiereserven nutzen können, größere Eier zu produzieren, denn die Ei-Größe kann vorkommen häufiger zu geringerem Nestlingsgewicht und höherer
Ergebnisse sich positiv auf die Schlupfrate auswirken. 7 Zudem wird durch die Ei-Größe oft das Sterberate durch verhungern führen, 2 kann dieses Ergebnis darauf
Gewicht der Schlüpflinge bestimmt, da größere Eier meist größere bzw. schwerere Küken hindeuten, dass der Effekt der Frühjahrs Temperatur auf die Ei-
Diskussion hervorbringen. 10 Dies kann besonders bei folgendem Nahrungsmangel von Vorteil sein, Größe in qualitativ schlechteren Habitaten stärker ausgeprägt sein
da Küken mit mehr Körpergewicht häufiger Hungertage überleben. 13 könnte. Zudem könnte der Wärmeeffekt durch die
Es ist bereits aus vorherigen Studien bekannt, dass größere Eier tatsächlich in Ausnahme
Umgebungseigenschaften noch verstärkt werden, da in urbanen
Jahren auftreten können. 4
Gebieten die Wärme besser gehalten und Strahlung stärker
reflektiert werden kann, 5 wodurch insbesondere diese Habitate
II. Temperatur
stärker durch steigende Temperaturen beeinflusst werden könnten.
Das Gelege wird signifikant von den Temperaturen der Zeit des 1. März bis 20. April Die zu Beginn aufgestellte Hypothese kann demnach nur zum Teil
beeinflusst und ein Zusammenhang zwischen der Wärmesumme dieses Zeitraums kann und nur für bestimmte Habitate bestätigt werden. Für das
beobachtet werden.11 Dies kann durch die Einordnung der Ergebnisse mithilfe der Wiesenhabitat konnte zusätzlich ein Effekt der Gelegegröße auf die
. Wärmesummen des relevanten Zeitraums der untersuchten Jahre zum Teil bestätigt Ei-Größe zwischen Jahren nachgewiesen werden. Studien haben
werden. Im Vergleich der Wärmesummen, zeigt sich , dass der relevante Zeitraum im Jahr gezeigt, dass die Energie eines erhöhten Nahrungsvorkommens
2014 mit einer Summe von 758,8 °C wärmer war, als die restlichen Jahre (411,5 – 685,6 ebenso in die Produktion größerer Gelege mit kleineren Eiern
°C). Für das Stadthabitat konnte nachgewiesen werden, dass die Eier aus dem Jahr 2014 investiert werden könnte.1 In weiteren Studien sollte daher
ein signifikant höheres Volumen hatten. Auch im Jahr 2018 waren konnte im Vergleich untersucht werden, inwiefern sich Habitatqualität und -
mit drei von fünf Jahren ein signifikant höheres Volumen festgestellt werden. Für das eigenschaften auf die Sensibilität von Individuen bezüglich
Wiesenhabitat war das Volumen der Eier in diesen beiden Jahren, außer im Vergleich mit klimatischer Umwelteinflüsse, auswirkt und was die Bedingungen
2015 (mit der geringsten Wärmesumme von 411,5°C), nicht signifikant höher. dafür sind, ob in größere Gelege oder größere Eier investiert wird.
Literatur 7 KOUIDRI,
1 BAŃBURA, M., GLĄDALSKI, M., KALIŃSKI, A., MARKOWSKI, M., SKWARSKA, J., WAWRZYNIAK, J., & BAŃBURA, J. (2018). A consistent long-lasting pattern of spatial variation in egg size and M., ADAMOU, A. E., BAŃBURA, A., OUAKID, M. L., CHABI, Y., & BAŃBURA, J. (2015). High egg size variation in African blue tits Cyanistes caeruleus ultramarinus on the periphery of species range. Acta
shape in blue tits (Cyanistes caeruleus). Frontiers in zoology, 15(1), 34. Ornithologica, 50(2), 205-212.
2 CHAMBERLAIN, D. E., CANNON, A. R., TOMS, M. P., LEECH, D. I., HATCHWELL, B. J., & GASTON, K. J. (2009). Avian productivity in urban landscapes: a review and meta‐analysis. Ibis, 8 LESSELS, C. M., BOAG, P. T. (1987) . Unrepeatable repeatabilities: a common mistake. Auk 104:116-121.
9 MENZEL, A., SPARKS, T. H., ESTRELLA, N., KOCH, E., AASA, A., AHAS, R., & CHMIELEWSKI, F. M. (2006). European phenological response to climate change matches the warming pattern. Global change biology,
151(1), 1-18.
3 DEUTSCHER WETTERDIENST (DWD) (2020). Mittlere Tagesmitteltemperatur im Frühling in Deutschland 1881 bis 2019. Mitteilung vom 27.01.2020. 12(10), 1969-1976.
4 ENCABO, S. I., MONRÓS, J. S., & BARBA, E. (2001). Egg size variation in a Mediterranean Great Tit Parus major population. Ardeola, 48(1), 63-70. 10 OJANEN , M., ORELL, M., & VÄISÄNEN, R. A. (1981). Egg size variation within passerine clutches: effects of ambient temperature and laying sequence. Ornis Fennica, 58(3), 93-108.
5 HEINL, M., HAMMERLE, A., TAPPEINER, U. & LEITINGER, G. (2015). Determinants of urban–rural land surface temperature differences – A landscape scale perspective. Landscape 11 PERRINS, C. M. (1991). Tits and their caterpillar food supply. Ibis, 133, 49-54.
12 R CORE TEAM (2019). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria
and Urban Planning, 134, 33-42.
6 HÕRAK, P., MAEND, R., OTS, I., & LEIVITS, A. (1995). Egg size in the Great Tit Parus major: individual, habitat and geographic differences. Ornis fennica, 7(3), 97-114. 13 VAN NOORDWIJK, A. J., KEIZER, L. C. P., VAN BALEN, J. H., & SCHARLOO, W. (1981). Genetic variation in egg dimensions in natural populations of the Great Tit. Genetica, 55(3), 221-232.
e-Mail der Autorin: lena.gonther@bio.uni-giessen.de Bild Gelege lizenzfrei von https://pixabay.com/de/photos/kohlmeise-das-nest-zu-verlassen-5178281/ Bild Kohlmeise lizenzfrei von https://pixabay.com/de/photos/kohlmeise-meise-singvogel-3494874/
Sie können auch lesen

















































