B-Zellen und Antikörper - Monika Raulf-Heimsoth - Vorlesung 11.05.2011
←
→
Transkription von Seiteninhalten
Wenn Ihr Browser die Seite nicht korrekt rendert, bitte, lesen Sie den Inhalt der Seite unten
B-Zellen und Antikörper
Monika Raulf-Heimsoth
Institut für Prävention und Arbeitsmedizin der
Deutschen Gesetzlichen Unfallversicherung,
Institut der Ruhr-Universität Bochum (IPA)
Vorlesung 11.05.2011
Themenkomplexe
Antikörper
1) Aufbau
2) Entstehung
3) Funktion
2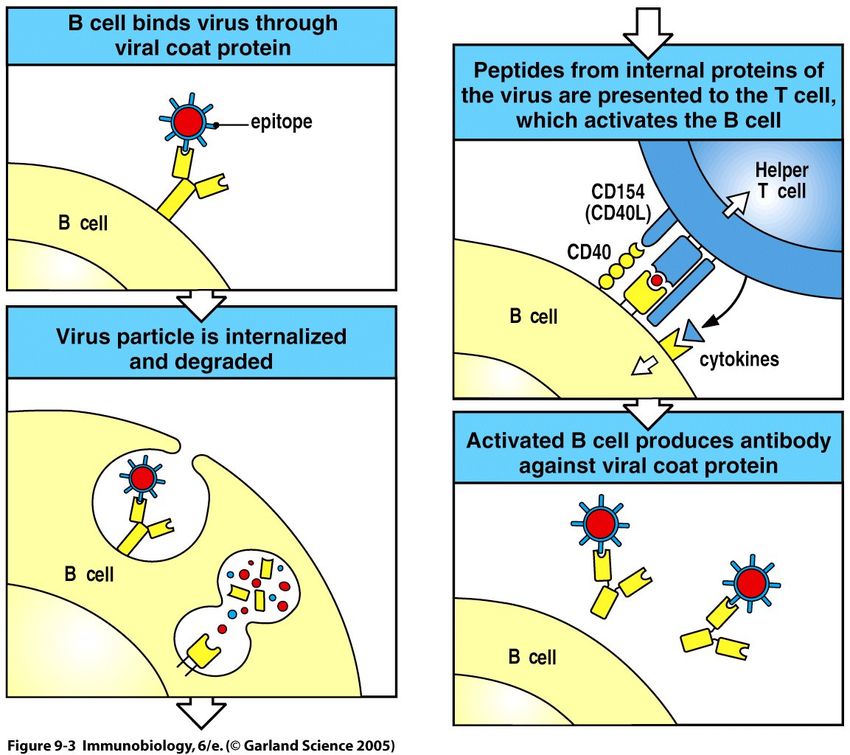
Was sind Antikörper?
Ein Antikörper ist ein Protein, das spezifisch an eine
bestimmte Substanz binden kann, d.h. an sein Antigen.
Aufgrund seiner einzigartigen Struktur kann jedes
Antikörpermolekül das entsprechende Antigen
spezifisch binden. Alle Antikörper haben jedoch
dieselbe Gesamtstruktur, und man fasst sie unter der
Bezeichnung Immunglobuline (Ig) zusammen.
Antikörper werden als Reaktion auf eine Infektion oder
Immunisierung von Plasmazellen erzeugt. Sie binden
und neutralisieren Krankheitserreger oder bereiten sie für
die Aufnahme und Zerstörung durch Phagozyten vor.
3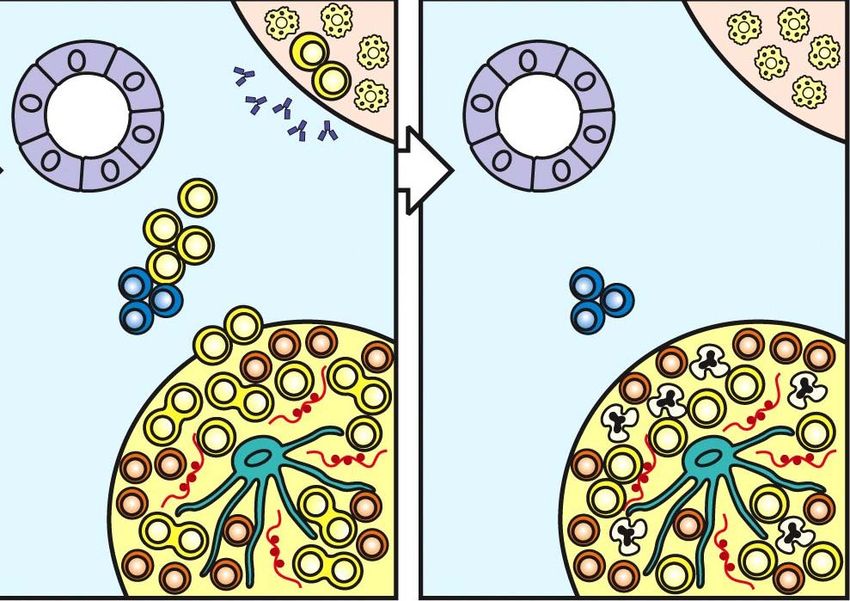
Die Struktur eines typischen Antikörpermoleküls I
2 Schwere Ketten (CH), 4-5 Domänen
2 leichte Ketten (CL), 2 Domänen
Die Ketten sind untereinander (CH-CH und CH-
CL) durch Disulfid-Brücken verbunden.
Jede Kette besitzt eine variable (N-terminal)
und 1-4 konstante (C-terminal) Domäne.
Die variablen Domänen bilden die
Antigenbindungsstelle aus (2 pro Ak).
Die konstanten Domänen vermitteln die
Antikörperfunktion (z.B. Rezeptorbindung)
4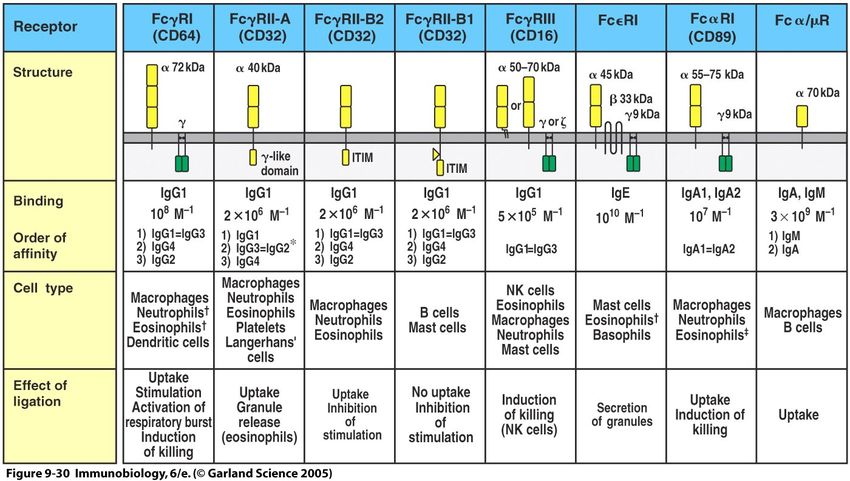
Die Struktur eines typischen Antikörpermoleküls II
Die einzelnen Domänen Die einzelnen Domänen
können zusätzlich und Ketten sind
Kohlenhydratstrukturen umeinander verdrillt.
tragen
5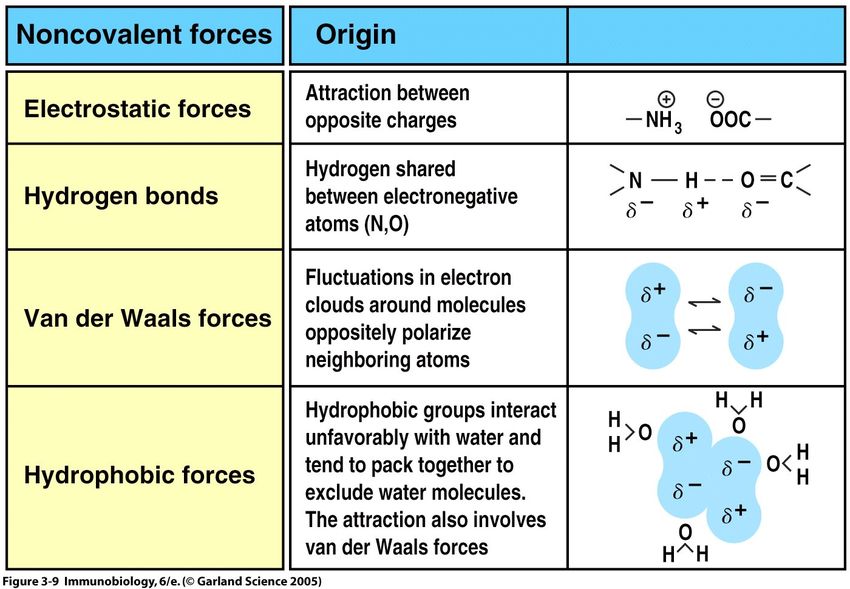
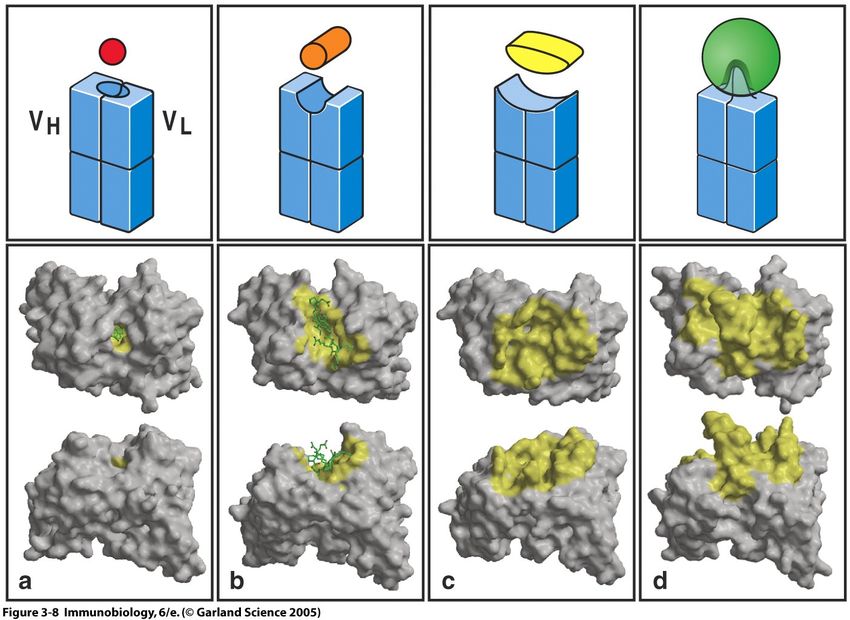
Die Bindung eines Antigens I
Antikörperbindung immer zwischen
Epitop (Antigen) und Paratop (Antikörper)
6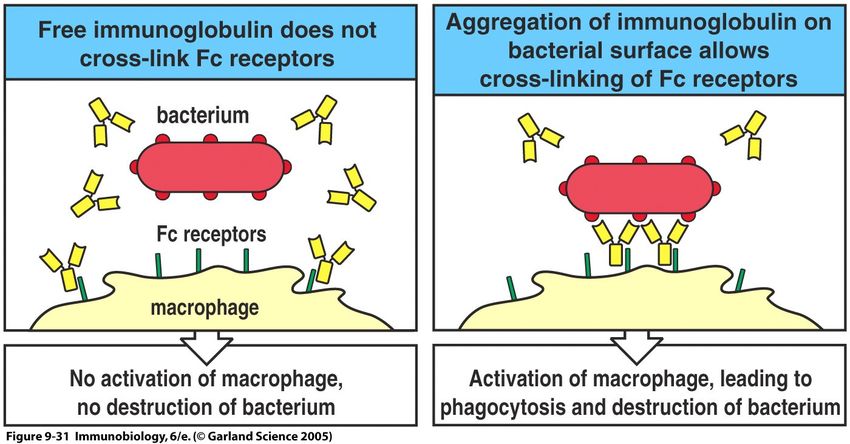
Die Bindung eines Antigens II
Die Bindung eines Antigens durch einen Antikörper ist nie kovalent.
7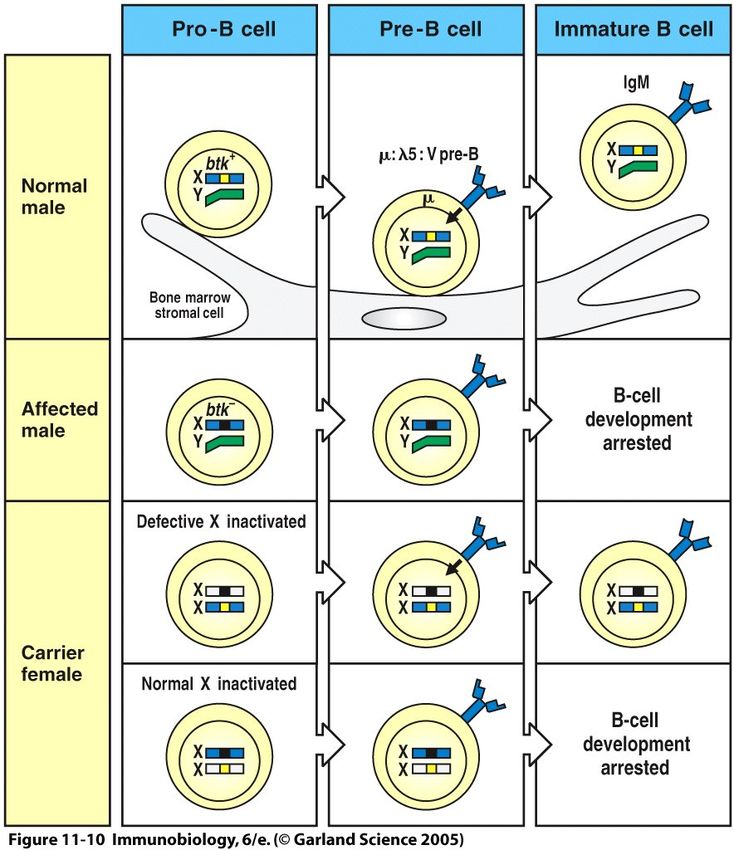
Entstehung der Antikörper-
vielfalt I
Möglichkeiten:
1) Keimbahntheorie: Für jeden Antikörper gibt es ein
eigenes Gen
Aber: es gibt 1x1011 verschiedene Antikörper
Der menschliche Körper besteht aus ca. 1x1014
(100 Billionen) Zellen, davon sind 3,5x1013
Gewebszellen (u.a. 1010 Neurone (Nervenzellen),
Rest Blutzellen
2) Somatische Diversifikation: Wenige vorhandene
Gensequenzen verändern sich und schaffen
dadurch die notwendige Vielfalt
8
Entstehung der Antikörper-
vielfalt II
C J D V
9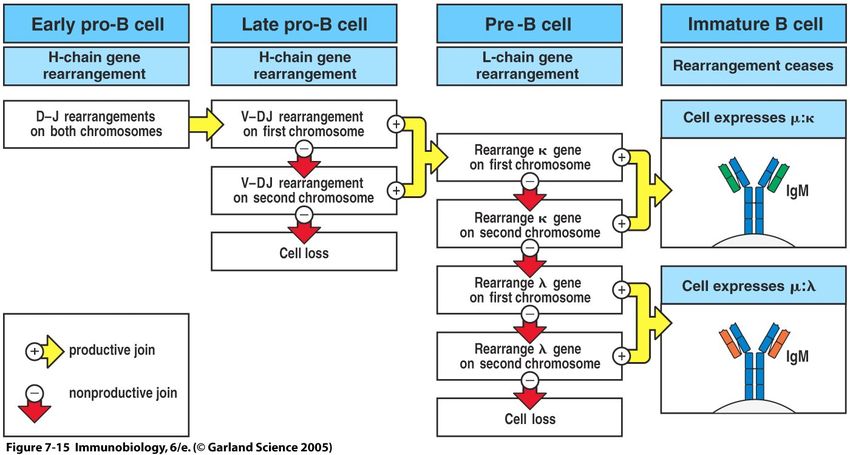
Somatische Rekombination I
Kombinatorische Vielfalt:
Leichte Kette κ: 40 VL-Segmente
5 JL-Segmente
40 x 5 = 200
Leichte Kette λ: 29 VL-Segmente
4 JL-Segmente
29 x 4 = 116
= 316 leichte Ketten
10Somatische Rekombination II
Kombinatorische Vielfalt:
Schwere Kette:
51 VH-Segmente
27 DH-Segmente
6 JH-Segmente
51 x 27 x 6 = 8262
8262 schwere Ketten
11Somatische Rekombination III
316 leichte Ketten x 8.262 schwere Ketten = 2,5 x 106
12Junktionale Vielfalt
An den Verbindungsstellen der
Gensegmente werden weitere Nucleotide
eingefügt oder entfernt
(P- und N-Nucleotide).
2/3 der Nucleotid-Einschübe sind
unproduktiv.
13Somatische Hypermutation
Affinitätsreifung der Antigen-Bindungsstellen findet in den
sekundären lymphatischen Organen statt (nur in aktivierten B-
Zellen, die bereits ein Antigen gebunden haben).
Mutationen treten in der gesamten variablen Region des
Antikörpers auf .
Mutationen treten in bestimmten Regionen gehäuft auf
(hot spots).
Antikörper-produzierende Zellen, deren Antikörper durch die
Mutationen negativ verändert werden, sterben durch
Apoptose.
Der Mechanismus der somatischen Hypermutation ist
weitestgehend unbekannt.
14Zusammenfassung
Antikörper können gegen fast jede beliebige Substanz
hergestellt werden
Um die dafür notwendige Antikörpervielfalt bereitzustellen,
gibt es drei Mechanismen:
1) Kombinatorische Vielfalt
(Somatische Rekombination)
2) Junktionale Vielfalt
(ungenaue Verknüpfung)
3) Somatische Hypermutation
(Affinitätsreifung in Keimzentren)
15Antikörperklassen - Isotypen I
Immunglobuline unterscheiden sich
• in der Anzahl der konstanten Domänen
• in Position und Anzahl der Disulfidbrücken
• in Anzahl und Lage der Kohlenhydrat-Seitenketten
• im Vorhandensein einer Gelenkregion
16Antikörperklassen - Isotypen II
IgM bildet Pentamere
• Erhöhung der Affinität
Æ Avidität
17Affinität - Avidität
IgM gleicht eine niedrige Affinität durch hohe Avidität aus.
Affinität = Bindungsstärke zwischen einer Antigenbindungsstelle
und dem Antigen.
Avidität = Bindungsstärke zwischen mehreren Antigenbindungs-
stellen und einem multivalenten Antigen.
Assoziation Dissoziation
Dissoziation
Assoziation
Avidität
Affinität
18Antikörperklassen - Isotypen III
IgA bildet Dimere
• Sekretion des Antikörpers
(Garland Science 2005)
19Antikörperklassen - Isotypen IV
cμ = IgM, cδ = IgD, cγ = IgG, cε = IgE, cα = IgA
Ψ = Pseudogen
Die Expression der Gene der konstanten Region
wechselt während der Reifung der B-Zelle =
Klassenwechsel (Isotyp-Switch).
20Antikörperklassen - Isotypen V
Beim Switch werden die
zwischen den Schalter-
regionen liegenden
Genabschnitte
herausgenommen
Æ Reifung nur in eine
Richtung
21Antikörperklassen - Isotypen VI
• Spezifische DNA-Sequenzen (Switch-Regionen)
regulieren die Rekombination
• Der Isotyp-Switch findet im Lymphknoten
(Keimzentrum) statt
• Er erfordert ein spezifisches Milieu aus Zytokinen, die
ihrerseits die Switchfaktoren
(Transkriptionsfaktoren) aktivieren
22Isotyp-Switch
Schwere Kette
JH1-6
Cμ Cδ Cγ 3 Cγ1 ωCε Cα1 Cγ2 Cγ4 Cε Cα2
IL-4
IL-5
IFN-γ
TGF-β
= induziert = steigert Synthese
= hemmt
23Isotyp-Switch
Im Laufe jeder B-Zell-Entwicklung findet ein Isotyp-Switch statt.
Primär wird IgM gebildet, später weitere Ig-Klassen, die dann
auch eine Affinitätsreifung durchmachen.
24Verlauf von Antikörpertitern
Kontakt Krankheitsbeginn
Inkubationszeit Akute Phase Rekonvaleszenz
Frühe Antikörper Spät- bzw. Schutz-Antikörper
IgG
IgM
IgA
Die Inkubationszeit von Erregern ist unterschiedlich. Sie kann zwischen
2 und 6 Tagen (Influenza) und 60-180 Tagen (Hepatitis B) betragen.
25Antikörperklassen - Funktion I
1) Neutralisierung (Viren, Bakterien, Toxine):
IgG und IgA
26Antikörperklassen - Funktion II
2) Opsonierung (unterstützt Phagozytose): IgG1 und IgG3
27Antikörperklassen - Funktion III
3) Komplement-Aktivierung (klassischer Weg): IgG und IgM
28Antikörperklassen - Funktion IV
Die Verteilung der Antikörperrezeptoren auf den verschiedenen Zellen
des Immunsystems/der Gewebe bestimmt den Ort der Wirkung des
Antikörpers.
Funktion IgM IgD IgG IgA IgE
Neutralisierung + - ++ ++ -
Opsonierung - - ++ + -
Mastzell-
- - - - +++
Sensibilisierung
Komplement-
+++ - + + -
Aktivierung
Transport durch
+ - - +++ -
Epithelien
Transport durch
- - +++ - -
Plazenta
Diffusion ins
+/- - +++ ++ +
Gewebe
Serumspiegel
1,5 0,04 13,5 2,1 0,005
[mg/ml]
29Antikörperklassen - Funktion V
• IgG kann durch die Plazenta an
den Embryo weitergegeben werden
• IgG und IgM sind die dominie-
renden Ak-Klassen im Plasma
• IgA wird in allen Schleimhäuten
sezerniert (Æ Schutzbarriere)
• IgA kann mit der Muttermilch an
den Säugling weitergegeben
werden
• IgE findet man auf Zellen in der
Haut und den Schleimhäuten
30Antikörperklassen - Funktion VI
Vorübergehender Immundefekt (Immunglobulinmangel)
nach der Geburt Æ hohe Infektanfälligkeit von Säuglingen
31Antikörper-Rezeptoren I
Die Rezeptorfunktion bestimmt ebenfalls die Antikörper-Wirkung
32Antikörper-Rezeptoren II
Die Rezeptorfunktion bestimmt ebenfalls die Antikörper-Wirkung
33Cytolytische Mechanismen von NK-Zellen IV
ADCC (antibody-dependent cellular cytotoxicity): Lyse von
Antikörper-markierten Zellen durch Apoptose und Nekrose
(Granzyme und Perforin). NK-Zellen tragen den Fcγ-Rezeptor (CD16)
34Antikörper-Rezeptoren III
Die Rezeptorfunktion bestimmt ebenfalls die Antikörper-Wirkung
35Themenkomplexe
B-Zellen
• Entstehung
• Aktivierung
36B-Zell-Entstehung I
• B-Zellen entstehen laufend im
Knochenmark (bone marrow) aus
lymphatischen Vorläuferzellen
• Die Umgebung (Stroma-Zellen des
Knochenmarks) liefern das notwen-
dige Milieu (Oberflächenmoleküle
und Cytokine) für die Entwicklung
• Die Immunglobulingene werden
umgeordnet; dieser Prozess läuft
unabhängig von Antigenen ab
37B-Zell-Entstehung II
• Die gebildete schwere Kette wird mit einer Ersatz-L-Kette z.T. an der Oberfläche der
großen Prä-B-Zelle exprimiert (Prä-B-Zell-Rezeptor).
• Nach der erfolgreichen Expression der schweren Kette teilt sich die große Prä-B-
Zelle aktiv, bevor die Umlagerung der leichten Kette beginnt
38B-Zell-Entstehung III
• Beginn der V-J-Umlagerung am κ-Locus, später folgt bei Bedarf der
λ-Locus
• Die Entwicklung zur reifen B-Zelle findet außerhalb des KM statt
39B-Zell-Entstehung – Zusammenfassung I-III
40B-Zell-Defekte I, XLA
Angeborene Immunschwächekrankheiten beruhen auf
rezessiven Gendefekten
Viele der häufigsten Immunschwächekrankheiten
beruhen auf defekten Genen auf dem X-Chromosom.
Männer erkranken, Frauen bleiben meistens gesund.
Beispiele:
X-gekoppelte Agammaglobulinämie (XLA):
keine B-Zellen
41B-Zell-Defekte II, XLA
Niedrige Antikörper-
konzentrationen führen zu
häufigen Infektionen mit
extrazellulären Bakterien
XLA (X-linked Agamma-
globulinämie) = defekte
Tyrosinkinase (btk), auf dem
X-Chromosom codiert.
btk überträgt Signale des Prä-
B-Zell-Rezeptors (B-Zell-
Reifung).
42B-Zell-Entstehung IV
Im Knochenmark:
Selektion auf Selbst-Toleranz
• Kontakt zu Antigenen in der
Umgebung.
• Unreife B-Zellen, die stark an diese
Antigene binden, sterben ab
(Apoptose), binden sie nur
schwach, werden sie inaktiviert
(Anergie).
• Eine Möglichkeit, der Deletion zu
entkommen, ist „Rezeptor-Editing“.
43B-Zell-Entwicklung
• Selbst-tolerante, naive B-Zellen wandern aus dem
Knochenmark aus.
• Naive B-Zellen zirkulieren durch das Blut in die
sekundären lymphatischen Organe.
• Treffen die B-Zellen in den sek. Lymphorganen auf ihr
passendes Antigen, werden sie aktiviert
(Umwandlung zur Antikörper-sezernierenden
Plasmazelle).
• Ohne Antigenkontakt zirkulieren sie wieder zurück mit
der Lymphe in die Blutbahn.
• Lebensdauer einer nicht-aktivierten B-Zelle:
ca. 3 Tage.
44Themenkomplexe
B-Zellen
• Entstehung
• Aktivierung
45B-Zell-Aktivierung I
Naive B-Zellen benötigen 2 Signale zur Aktivierung:
Das erste Signal liefert die Bindung eines passenden
Antigens an den B-Zell-Rezeptor (BCR) = Oberflächen-
Immunglobulin
Das zweite Signal kann auf zwei Wegen geliefert werden:
1) Thymus-abhängig (durch bereits aktivierte T-Helfer-
Zellen) = TD-Antigene (thymus dependent)
2) Thymus-unabhängig (nur durch Antigen aktiviert) =
TI- Antigene (thymus independent)
46TI-B-Zell-Aktivierung I
TI-1-Antigene aktivieren
neben dem BCR einen
weiteren Rezeptor auf der TI-2-Antigene aktivieren die B-
Oberfläche, z.B. CD14 gegen Zelle über gleichzeitige
LPS oder einen Vernetzung des BCR über viele
Komplementrezeptor = gleichförmige Antigene, keine Co-
co-stimulatorisches Signal Stimulation
47TI-B-Zell-Aktivierung II
TI (1+2) -Aktivierung : B-Zelle Æ Plasmazelle
¾ kein Keimzentrum
¾ keine Gedächtniszellen
¾ keine Affinitätsreifung
¾ switch bis IgG3, IgG2 (wodurch?)
48TD-B-Zell-Aktivierung I
49TD-B-Zell-Aktivierung II
Linked recognition = T-Zelle und B-Zelle erkennen das
gleiche Antigen
CD4
T-Zell-Rezeptor
Peptid
MHC II
50TD-B-Zell-Aktivierung II
CD4
T-Zell-Rezeptor
Peptid
MHC II
Eine T-Zelle exprimiert die co-stimulatorischen Signale erst, wenn sie
ihrerseits durch eine professionelle Antigen-präsentierende Zelle
(APC) spezifisch aktiviert worden ist (dendritische Zellen, Makrophagen).
51TD-B-Zell-Aktivierung II
Linked recognition = T-Zelle und B-Zelle erkennen das
gleiche Antigen
52Zusammenfassung: TD-B-Zell-Aktivierung
Signale für die B-Zell-Aktivierung:
1) Bindung eines Antigens über den BCR
(B-Zell-Rezeptor = membranständiges Immunglobulin)
2) Signale der T-Zelle, die das MHC-II-präsentierte Peptid
mittels TCR und Corezeptor CD4 erkennt:
• Stimulation des CD40 (B-Zelle) durch CD40-Ligand
(T-Zelle)
• Cytokine
• Weitere co-stimulatorische Signale
53TD-B-Zell-Aktivierung III
Cytokine aus der Th-Zelle bewirken
• Die Proliferation der B-Zelle (v.a. IL-5, IL-6)
• Den Isotyp-Switch (z.B. IFN-γ zum IgG-switch, IL-4)
B-Zellen werden nach der Aktivierung zu
• B-Gedächtniszellen
• Antikörper-sezernierenden Plasmazellen
54Primäre humorale Immunantwort I
T-Zellen werden von dendritischen Zellen in Lymphknoten gelockt und
aktiviert, wenn sie das präsentierte Antigen erkennen.
© Nature Immunology 7(2006), 329-332
grün = dendritische Zellen
rot = T-Zellen
55Primäre humorale Immunantwort II
Die primäre Aktivierung der naiven
B-Zelle durch eine aktivierte Th-Zelle
geschieht ebenfalls in den sekundären
Lymphorganen.
B-Zellen wandern über Venolen mit
hohem Epithel (HEV) in die
Lymphorgane ein und verlassen sie,
wenn sie auf kein spezifisches Antigen
treffen.
56Primäre humorale Immunantwort III
B-Zellen, die ein spezifisches Antigen treffen und in der T-Zell-Zone auf
passende aktivierte T-Zellen treffen bilden zusammen mit diesen T-
Zellen einen sog. Primärfocus, in dem beide Zelltypen proliferieren.
grün = T-Zellen
rot = B-Zellen
© Nature Immunology 7(2006), 329-332
57Primäre humorale Immunantwort III
Die B-Zellen wandern z.T. aus dem Primärfocus
in die Markstränge Æ Plasmazellen
(Antikörperproduktion, Isotyp-Switch).
58Sekundäre humorale Immunantwort I
Einige B- und T-Zellen verlassen die Primär-Foci und wandern in einen
primären Lymph-Follikel, wo sie Keimzentren bilden. Die B-Zellen
proliferieren rasch (Centroblasten) und durchlaufen die somatische
Hypermutation.
59Zusammenfassung:
sek. Lymphorgane I
Aktivierung der B-
Zelle durch T-Zelle
Lymphfollikel, Primäre Foci
3 Tage Mark-
Keimzentrum
stränge
6 Tage
Plasmazellen
Hypermutation,
Affinitätsreifung Ak-Freisetzung,
switch
Rezirkulation
Plasma- Gedächtnis- Langlebige
Apoptose
zellen zellen Plasma-
zellen
Sek. lymph. Organe
Ak-Freisetzung Knochenmark
60Zusammenfassung:
sek. Lymphorgane II
Gedächtnis- B-Gedächtniszellen (Memory B-Zellen)
zellen zirkulieren.
Sie benötigen Antigen zur
Antikörperproduktion und teilen sich
stetig in geringer Frequenz.
langlebige Langlebige Plasmazellen entstehen v.a.
Plasmazellen nach einem 2. Antigenkontakt
Sie wandern ins Knochenmark (Nische,
die das Überleben garantiert).
Sie benötigen kein Antigen zur
Antikörperproduktion.
Evtl. überleben sie Jahre/Jahrzehnte.
61Zusammenfassung:
sek. Lymphorgane III
B-Zellen benötigen bei der Affinitätsreifung
unbedingt 2 positive Signale
(sonst Æ Apoptose)
Signal 1: Antigen erkannt
Signal 2: T-Zell-Interaktion
62Woher stammen die Antigene
in den Keimzentren?
1) FDC = Follikulär Dendritische Zellen
• FDCs sind in den Keimzentren
und locken die B-Zellen über
Chemokine dort hin.
• FDCs präsentieren Antigene nicht
über MHC sondern Fc- und
Komplement-Rezeptoren. Sie
können keine T-Zellen aktivieren.
63Sie können auch lesen

















































