Die Effekte des NSM-Inhibitors GW4869 auf Affektivität und Trinkverhalten - im Verhaltensmodell an der Maus - OPUS 4
←
→
Transkription von Seiteninhalten
Wenn Ihr Browser die Seite nicht korrekt rendert, bitte, lesen Sie den Inhalt der Seite unten

Psychiatrische und Psychotherapeutische Klinik des Universitätsklinikums Erlangen [Direktor: Prof. Dr. med. Johannes Kornhuber] Die Effekte des NSM-Inhibitors GW4869 auf Affektivität und Trinkverhalten – im Verhaltensmodell an der Maus Der Medizinischen Fakultät der Friedrich-Alexander-Universität Erlangen-Nürnberg zur Erlangung des Doktorgrades Dr. med. vorgelegt von Anna-Lisa Eberle aus Braunschweig
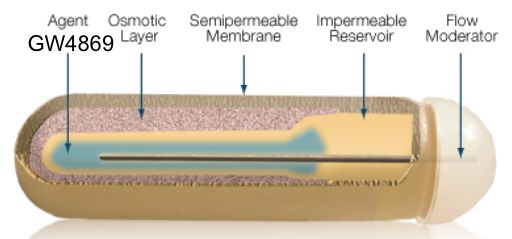
Als Dissertation genehmigt von der Medizinischen Fakultät der Friedrich-Alexander-Universität Erlangen-Nürnberg Vorsitzender des Promotionsorgans: Prof. Dr. med. Markus Neurath Gutachter: Prof. Dr. Christian P. Müller Gutachter: Prof. Dr. Johannes Kornhuber Tag der mündlichen Prüfung: 30. November 2021
Diese Dissertation ist meinen Eltern und meinem Verlobten gewidmet, die mich während meiner gesamten Studienzeit und in allen Lebenslagen unterstützt haben.
Inhaltsverzeichnis Zusammenfassung ....................................................................................... 1 Summary ........................................................................................................ 3 Einleitung ................................................................................................ 5 1.1 Sphingolipide und Sphingomyelinasen ......................................... 5 1.1.1 Übersicht .................................................................................................... 5 1.1.2 Sphingolipide in der Zellmembran.............................................................. 5 1.2 Die Familie der Sphingomyelinasen .............................................. 7 1.2.1 Saure Sphingomyelinase ........................................................................... 7 1.2.2 Neutrale Sphingomyelinase 2 .................................................................... 8 1.3 Alkoholkonsum und Alkoholsucht ............................................... 10 1.3.1 Übersicht .................................................................................................. 10 1.3.2 Alkohol und der Sphingolipidstoffwechsel ................................................ 10 1.3.3 NSM und Alkohol ..................................................................................... 12 1.4 Depression ..................................................................................... 12 1.4.1 Übersicht .................................................................................................. 12 1.4.2 Depression und der Sphingolipidstoffwechsel ......................................... 14 1.4.3 NSM und Depression ............................................................................... 15 1.5 Der NSM-Inhibitor GW4869 ........................................................... 15 Zielsetzung und Hypothesen der vorliegenden Arbeit ..................... 18 Material und Methoden ........................................................................ 19 3.1 Versuchstiere und Versuchsbedingungen .................................. 19 3.1.1 Tierhaltung ............................................................................................... 19 3.1.2 Versuchsgruppen ..................................................................................... 19 3.2 Versuchsablauf .............................................................................. 21 3.3 Osmotische Minipumpen .............................................................. 22 3.3.1 Aufbau der osmotischen Minipumpen ...................................................... 22 3.3.2 Funktionsprinzip der osmotischen Minipumpen ....................................... 23 3.3.3 Berechnung gemäß der Anleitung von ALZET ........................................ 24 3.3.4 Befüllung der osmotischen Minipumpen .................................................. 24 3.3.5 Operative Implantation der osmotischen Minipumpen ............................. 25 3.4 Experimentelle Tierversuche: Verhaltenstests ........................... 25 3.4.1 Emotionale Verhaltensparadigmen .......................................................... 25 3.4.2 Open Field Test........................................................................................ 26 3.4.2.1 Versuchsaufbau und -durchführung ................................................. 26
3.4.2.2 Fragestellung und Messparameter ................................................... 27 3.4.3 Elevated Plus Maze ................................................................................. 27 3.4.3.1 Versuchsaufbau- und durchführung ................................................. 27 3.4.3.2 Fragestellung und Messparameter ................................................... 28 3.4.4 Novelty Suppressed Feeding ................................................................... 28 3.4.4.1 Versuchsaufbau- und durchführung ................................................. 29 3.4.4.2 Fragestellung und Messparameter ................................................... 29 3.5 Experimentelle Tierversuche: Alkoholtrinkstudie ...................... 29 3.5.1 Zwei-Flaschen-Wasser-Alkohollösung-Wahlparadigma .......................... 29 3.5.2 Versuchsablauf ........................................................................................ 30 3.5.3 Messparameter ........................................................................................ 30 3.6 Tiertötung und Hirnentnahme ...................................................... 30 3.7 Statistische Auswertung ............................................................... 31 Ergebnisse ............................................................................................ 32 4.1 Open Field Test .............................................................................. 32 4.1.1 Aufenthaltsdauer im Zentrum ................................................................... 33 4.1.2 Zurückgelegte Distanz ............................................................................. 34 4.1.2.1 Zurückgelegte Distanz im Zentrum................................................... 34 4.1.2.2 Zurückgelegte Distanz in der Peripherie .......................................... 35 4.1.3 Eintritte ins Zentrum ................................................................................. 36 4.1.4 Latenz bis zum Eintritt ins Zentrum .......................................................... 37 4.2 Elevated Plus Maze ........................................................................ 37 4.2.1 Aufenthaltsdauer im EPM ........................................................................ 38 4.2.1.1 Aufenthaltsdauer auf den proximalen/distalen EPM-Armen ............. 40 4.2.2 Zurückgelegte Distanz im EPM ................................................................ 42 4.2.2.1 Zurückgelegte Distanz auf den proximalen/distalen EPM-Armen .... 43 4.2.3 Eintritte ins EPM....................................................................................... 45 4.2.3.1 Eintritte auf den proximalen/distalen EPM-Armen ............................ 47 4.3 Novelty Suppressed Feeding ........................................................ 49 4.3.1 Latenz bis zum ersten Fressen ................................................................ 49 4.3.2 Zurückgelegte Distanz bis zum ersten Fressen ....................................... 50 4.4 Alkoholtrinkstudie ......................................................................... 51 4.4.1 Alkoholkonsum ......................................................................................... 51 4.4.2 Wasserkonsum ........................................................................................ 54 4.4.3 Alkoholpräferenz ...................................................................................... 57 4.4.4 Gesamter Flüssigkeitskonsum ................................................................. 60
Diskussion ............................................................................................ 63 5.1 Alkoholtrinkstudie ......................................................................... 63 5.1.1 Zusammenfassung und Interpretation der Ergebnisse...................... 63 5.1.1.1 GW4869 führt zu verändertem Alkoholtrinkverhalten, deutlicher Alkoholpräferenz- und Alkoholkonsumanstieg bei den Weibchen ...................... 63 5.1.2 Die Rolle der NSM bei Alkoholtrinkverhalten ...................................... 64 5.2 Verhaltensparadigmen .................................................................. 66 5.2.1 Zusammenfassung und Interpretation der Ergebnisse...................... 66 5.2.1.1 GW4869 führt zu ängstlichem Verhalten im OFT, anxiogener Effekt bei den Weibchen ................................................................................................ 66 5.2.1.2 GW4869 führt zu ängstlichem Verhalten im EPM, kein Geschlechtereffekt ....................................................................................... 66 5.2.1.3 GW4869 führt zu depressivem Verhalten im NSF, depressiver Effekt bei den Männchen ............................................................................................... 67 5.2.2 Die Rolle der NSM bei Emotionalität (Angst/Depression) .................. 67 5.3 Die Rolle der NSM im Zusammenhang von Affektivität und Trinkverhalten .......................................................................................... 69 5.4 Stärke der vorliegenden Arbeit ..................................................... 72 5.5 Limitation der vorliegenden Arbeit .............................................. 73 5.6 Ausblick .......................................................................................... 73 Literaturverzeichnis ............................................................................. 74 Anhang .................................................................................................. 88 7.1 Abbildungsverzeichnis .................................................................. 88 7.2 Tabellenverzeichnis ....................................................................... 90 7.3 Abkürzungsverzeichnis ................................................................. 91 Danksagung .......................................................................................... 93
Zusammenfassung Hintergrund und Ziele. Der schädliche und unangemessene Konsum von Alkohol ist ein weltweites Problem und gewinnt in der heutigen Zeit und in den westlichen Industrieländern zunehmend an Bedeutung (Heath, 2000). Er kann neben einer Alkoholkonsumstörung und Alkoholsucht (European Monitoring Center for Drugs and Drug Addiction (EMCDDA), 2012) zu vielen somatischen Erkrankungen, wie zum Beispiel Erkrankungen des Herz-Kreislaufsystems oder Tumorerkrankungen führen. Schließlich können auch psychiatrische Erkrankungen wie die Depression und Angststörungen auftreten oder sich als Folge dieser entwickeln (Grant and Harford, 1995, Müller et al., 2017, Schuckit et al., 1997). Ein Erklärungsansatz für diese Komorbiditäten liegt in der Regulation des Sphingolipidhaushalts durch das Enzym Neutrale Sphingomyelinase 2 (NSM). In der vorliegenden Arbeit wurde im pharmakologischen Modell der NSM-Inhibition durch GW4869 das Alkoholtrinkverhalten von Mäusen sowie deren Emotionalität untersucht und im Hinblick auf die Regulation der Sphingolipidhomöostase durch die NSM analysiert. Die Hypothese lautete in diesem Zusammenhang, dass der NSM- Inhibitor GW4869 das Trinkverhalten und das emotionale Verhalten in entsprechen- den Tests verändert und diese Effekte geschlechtsabhängig sind. Methoden. Für die Experimente wurden 31 naive C57Bl6-Mäuse verwendet, jeweils 16 Männchen und 15 Weibchen. Grundvoraussetzungen für die Durchführung der Experimente war es, der Hälfte der Tiere (sowohl Männchen als auch Weibchen) den NSM-Inhibitor GW4869 kontinuierlich mit einer Dosis von 2 mg/kg/Tag mittels osmo- tischer Minipumpen zu verabreichen, um so die Effekte der pharmakologischen Herunterregulierung der NSM-Aktivität auf Trinkverhalten und Emotionalität zu untersuchen. Der Kontrollgruppe wurde Saline (0,9 % NaCl) verabreicht (n = 8 bzw. 7 Mäuse/Versuchsgruppe). Für die Alkoholtrinkstudie wurde ein Zwei-Flaschen-Wasser-Alkohol-Wahlparadigma benutzt und für die Verhaltensexperimente dienten der Open Field Test (OFT), das Elevated Plus Maze (EPM) und das Novelty Suppressed Feeding (NSF). Ergebnisse In der hier durchgeführten Alkoholtrinkstudie konnte herausgefunden werden, dass die pharmakologische Hemmung der NSM durch GW4869 bei Mäusen zu einem veränderten Trinkverhalten führt und dieser Effekt geschlechtsabhängig ist. Es zeigten sowohl GW4869-Weibchen als auch GW4869-Männchen einen höheren Alkoholkonsum gegenüber der mit Saline behandelten Kontrollgruppe. Insbesondere 1
bei den GW4869-Weibchen gab es mit dem starken Anstieg des Alkoholkonsums und dem gleichzeitig starken Abfall im Wasserkonsums eine klare Verschiebung der Präferenz hin zum Alkohol bei hohen Alkoholkonzentrationen (12 und 16 Vol.-%). Bei den GW4869-Männchen lag die Alkoholpräferenz gegenüber Wasser bei den mittleren Alkoholkonzentrationen am höchsten. In der hier durchgeführten experimentellen Verhaltensstudie konnte außerdem beobachtet werden, dass die pharmakologische Hemmung der NSM durch GW4869 zu einem sichtbaren Effekt auf das Angst- und Depressionsverhalten führt und auch dieser geschlechtsabhängig ist. Der depressive Verhaltenseffekt war ausschließlich bedingt durch die Männchen mit einer signifikant höheren Latenzzeit bis zum ersten Fressen im NSF im Vergleich zur Kontrollgruppe. Hinsichtlich Angstverhalten zeigte sich insgesamt die Tendenz zum anxiogenen Effekt bei den GW4869-Weibchen im OFT. Schlussfolgerung. Insgesamt konnte mit den Ergebnissen meiner Arbeit bestätigt werden, dass die NSM bei neuronalen und psychiatrischen Pathologien eine bedeutende Funktion einnimmt. Die NSM scheint nach Zusammenschau der oben genannten Ergebnisse Einfluss auf die Regulation des Trink- und des Affekt- verhaltens zu nehmen. Eine Blockade der NSM durch GW4869 und damit eine Verringerung der NSM-Aktivität erhöht demnach das Konsumrisiko und die Konsummenge von Alkohol, insbesondere bei Frauen. Das wiederum bedeutet, dass der NSM-Inhibitor GW4869 ein essenzieller Modulator des Alkoholkonsums ist. Dieser Effekt ist stark weiblich dominiert und damit geschlechtsabhängig. Die NSM- Hemmung scheint geringere und dennoch sichtbare Einflüsse auf das Affekt- verhalten, in Form von depressiven Effekten bei Männern und anxiogenen Effekten bei Weibchen, zu haben. Insgesamt leistet diese Arbeit einen Beitrag zum besseren Verständnis der Regulationsmechanismen durch die NSM, mit dem Ziel psychiatrische Erkrankungen wie die Alkoholsucht, Depression und Angststörungen in Zukunft durch neue Behandlungsstrategien noch besser therapieren zu können. 2
Summary Background and aims. The harmful and inappropriate consumption of alcohol is a global problem and is becoming increasingly important in today's world and in the western industrialized countries (Heath, 2000). In addition to alcohol consumption disorders and alcohol addiction (European Monitoring Center for Drugs and Drug Addiction (EMCDDA), 2012), it can lead to many somatic diseases, such as diseases of the cardiovascular system or tumor diseases. Finally, psychiatric illnesses such as depression and anxiety disorders can also occur or develop as a result of them (Grant and Harford, 1995, Müller et al., 2017, Schuckit et al., 1997). One explanation for these comorbidities can be found in the regulation of the sphingolipid homeostasis by the enzyme neutral sphingomyelinase 2 (NSM). In the present work, the alcohol drinking behaviour of mice as well as their emotional behaviour was examined and analyzed with regard to the regulation of sphingolipid homeostasis by the NSM. For these experiments the pharmacological NSM inhibition with GW4869 was used. The hypothesis in this context was that the NSM inhibitor GW4869 changes drinking behaviour and emotional behaviour in corresponding tests and that these effects are gender dependent. Methods. 31 naive C57Bl6 mice were used, 16 males and 15 females each. The prerequisites for carrying out the experiments were to continuously administer the NSM inhibitor GW4869 to half of the animals (both males and females) by using osmotic mini-pumps at a rate of 2 mg/kg/day in order to investigate the effects of the pharmacological down-regulation of NSM activity on drinking behaviour and emotionality. The control group received saline (0,9 % NaCl) in the osmotic mini- pumps (n = 8 respectively 7 mice/test group). A two-bottle water-alcohol choice paradigm was used for the alcohol drinking study. The Open Field Test (OFT), the Elevated Plus Maze (EPM) and the Novelty Suppressed Feeding (NSF) were used as behaviour experiments. Results. In the alcohol drinking study it was observed that the pharmacological inhibition of NSM by GW4869 in mice leads to a change in drinking behaviour and that this effect is gender dependent. Both GW4869 females and GW4869 males showed a higher alcohol consumption compared to the control group treated with saline. Especially among the GW4869 females, there was a clear shift in preference towards alcohol with a strong increase in alcohol consumption and a simultaneous strong decrease in water consumption (12 % and 16 % alcohol). In the GW4869 males, the alcohol preference over water was highest at medium alcohol concentrations. 3
In the behaviour tests it could also be observed that the pharmacological inhibition of NSM by GW4869 leads to an effect on anxiety and depression behaviour and that it is also gender dependent. The depressive behavioural effect was stronger in males. They showed a significantly higher latency until the first eating in the NSF compared to the control group. With regard to anxiety behaviour, GW4869 females showed a tendency towards the anxiogenic behaviour in the OFT. Conclusion. Overall, the results of my work could confirm that the NSM plays an important role in neuronal and psychiatric pathologies.I was able to confirm that the NSM plays an important role in neuronal and psychiatric pathologies. After looking at the obtained results, the NSM seems to have an influence on the regulation of the drinking behaviour and the emotional behaviour. An inhibition of NSM by GW4869 and thus a reduction in NSM activity accordingly increases the risk and the amount of alcohol consumption, especially in women. This means that the NSM inhibitor GW4869 is an essential modulator of alcohol consumption. This effect is strongly dominated by women and is therefore gender dependent. The NSM inhibition appears to have less, but still visible influences on emotional behaviour, in form of depressive effects in males and anxiogenic effects in females. Overall, my work contributes to a better understanding of the regulatory mechanisms by the NSM, with the aim of improving the medical treatment of psychiatric diseases such as alcohol addiction, depression and anxiety disorders. 4
Einleitung 1.1 Sphingolipide und Sphingomyelinasen 1.1.1 Übersicht Zu den Sphingolipiden gehören unter anderem Sphingomyelin, Ceramid, Sphingosin und Sphingosin-1-Phosphat. Diese sind neben ihrer Aufgabe als Bestandteile der Zellmembran auch an einer Vielzahl von grundlegenden zellulären Prozessen beteiligt, wie zum Beispiel an Apoptose, Zellproliferation und Zelldifferenzierung (Ohanian and Ohanian, 2001, Futerman and Hannun, 2004, Baumruker and Prieschl, 2002, Hannun and Obeid, 2008, Müller et al., 2015b) und spielen außerdem eine wichtige Rolle bei psychiatrischen Erkrankungen (Müller et al., 2015a, Schwarz et al., 2008), worauf der Fokus der vorliegenden Arbeit liegt. Sphingomyelinasen wiederum sind Enzyme, die die Hydrolyse von Sphingomyelin in Ceramid und Phosphorylcholin katalysieren (Kolesnick, 1994, Schneider and Kennedy, 1967). Sie unterscheiden sich in ihren pH-abhängigen Isoformen. Diese sind die saure Sphingomyelinase (Acid Sphingomyelinase, ASM), die neutrale Sphingomyelinase (NSM) und die alkalische Sphingomyelinase (alkSMase) (Goñi and Alonso, 2002). Ceramid entsteht entweder über die Sphingomyelinase-induzierte Hydrolyse von Sphingomyelin oder durch de novo Synthese aus Palmitoyl-CoA und Serin (Perry and Hannun, 1998). Ceramid wird vermehrt gebildet durch verschiedenste Reize wie Wachstumsfaktoren, UV-bedingten Stress oder Zytokine (Ohanian and Ohanian, 2001). Ein erhöhter zellulärer Ceramidspiegel kann als second messenger wiederum über CAPPs (Ceramid-aktivierte Protein-Phospha- tasen) zum Anhalten des Zellzyklus, über Caspasen zur Apoptose und über den Raf- MEK-MAP (mitogen-activated protein) -Kinase-Weg zu inflammatorischen Pro- zessen führen. Dies wird Sphingomyelin-Signaltransduktionskaskade genannt (Kolesnick, 1994, Kolesnick et al., 2000, Ohanian and Ohanian, 2001, Goñi and Alonso, 2002). 1.1.2 Sphingolipide in der Zellmembran Die Zellmembran besteht aus einer Lipiddoppelschicht, dessen Grundgerüst vor allem Phospholipide, Sphingolipide und Cholesterin bilden. Diese sind amphiphatisch und damit sehr flexibel und dynamisch in der Bildung von Membranen nach dem Flüssig-Mosaik-Modell (Singer and Nicolson, 1972). Sphingolipide leiten sich vom ungesättigten Alkohol Sphingosin ab, machen insgesamt bis zu 20 % der Membranlipide aus und bestehen aus zwei langkettigen Fettsäuren und einer hydrophilen Kopfgruppe (van Meer and Lisman, 2002). Besonders häufig kommen sie im Gehirn vor (Holthuis et al., 2001). Die Synthese 5
dieser findet im endoplasmatischen Retikulum und im Golgi-Apparat statt. Der Transport in andere Zellorganellen und in die Plasmamembran erfolgt über Vesikel. Sphingolipide können auch frei zwischen den Membranen diffundieren (Gault et al., 2010) und von der zytosolischen auf die entgegengesetzte Plasmamembranseite (sog. Flip-Flop) wechseln (Kubelt, 2004). Durch den ständigen Umbau der Zell- membran und durch die Stoffwechselprozesse der Sphingolipide, die wiederum selbst als bioaktive Metabolite dienen, kommt es in der Zellmembran zu zellbiologisch bedeutsamen Prozessen. Dadurch können sowohl katabole als auch anabole Prozesskaskaden in Gang gesetzt oder die Sphingolipidzusammensetzung selbst verändert werden (Gault et al., 2010). Sphingomyelin, das Vorläufermolekül von Ceramid und das häufigste komplexe Sphingolipid in menschlichen Zellen, gehört neben Phosphatidylcholin, Phosphatidyl- ethanolamin und Phosphatidylserin zu den vier Hauptklassen der Phospholipide. Zusammen machen diese mehr als 50 % aller Lipide in zellulären Plasmamembranen aus und zählen zu den häufigsten Lipiden in den Zellmembranen des Gehirns (Jain et al., 2014). Aufgrund der sperrigen Kopfgruppe ist ein Wechsel des Ortes nur mit Hilfe von spezifischen Transportmolekülen und Flippasen möglich. Das Sphingo- myelin befindet sich deshalb entweder auf der zum Lumen des Golgi-Apparats gerichteten Seite oder auf der extrazellulären Seite der Zellmembran nach vesi- kulärem Transport dorthin. Der Sphingomyelin-Abbau durch das Enzym Sphingo- myelinase ist mitbestimmend für die Aufrechterhaltung der Membranhomöostase (Gault et al., 2010, Goñi and Alonso, 2004, Holthuis et al., 2001). Die Sphingomyelinasen befinden sich auf der zytosolischen Seite der Zellmembran. Das durch Hydrolyse entstehende Ceramid kann den Ort, an dem es entstanden ist, nicht verlassen und ist auf die Plasmamembran beschränkt, in der es entstanden ist. Durch die zelluläre Abschottung ist es in der Interaktion mit anderen Proteinen stark eingeschränkt. Es kann jedoch, genauso wie andere Sphingolipide durch den Flip- Flop-Mechanismus zwischen beiden Membranseiten der Lipiddoppelschicht hin und her wechseln. Deshalb entscheidet die Lokalisation der Sphingomyelinase auch den Wirkort des dadurch entstehenden Ceramids (Hannun and Obeid, 2008). Die Zellmembran ist nicht an jeder Stelle gleich aufgebaut. Es können sich spezialisierte Membranmikrodomänen bilden, die reich an Cholesterin und Sphingo- myelin sind. Sie werden Lipid Rafts genannt (Simons and Ikonen, 1997, Futerman and Hannun, 2004, Baumruker and Prieschl, 2002) und haben unter anderem G- Protein-gekoppelte Rezeptoren und andere Signalproteine eingelagert. Sie dienen damit als Plattform für komplexe Signalkaskaden und Immunantworten sowie für Endozytose, Transzytose und für Eintritte von pathogenen Molekülen in die Zelle 6
sowie für den Transport von Cholesterin und kleineren Molekülen wie Vitaminen und Lipiden über Potozytose (Huang and Feigenson, 1999, Recktenwald and McConnell, 1981, Robertson, 2018, Slotte and Ramstedt, 2007, van Meer et al., 2008, Gulbins and Kolesnick, 2003, Ohanian and Ohanian, 2001, Veiga et al., 2000). Eine spezielle Form der Lipid Rafts sind Caveolae, die Ω-förmige Einbuchtungen der Plasmamembran sind (Lingwood and Simons, 2010). Sie dienen durch Abschnürung der Zellmembran der Endozytose und Transzytose. Verändert sich die Zusammensetzung der Lipid Rafts, zum Beispiel durch Aktivitätssteigerung der Sphingomyelinasen, so wirkt sich dies direkt auf die Funktionalität der Lipid Rafts und der Caveolae aus (Fantini and Barrantes, 2009, Gulbins and Kolesnick, 2003). Aus diesem Grund will man noch besser verstehen, wie die Zusammensetzung der Zellmembranen und damit der Lipid Rafts reguliert und kontrolliert wird. 1.2 Die Familie der Sphingomyelinasen Bei Säugetieren gibt es, wie bereits in Kapitel 1.1.1 "Übersicht" beschrieben, drei unterschiedliche Sphingomyelinasen (Goñi and Alonso, 2002). Die saure, alkalische und neutrale Spingomyelinase (Gault et al., 2010). Aufgrund ihres unterschiedlichen pH-Optimums sind die Enzyme dementsprechend im menschlichen Körper zu finden: Die ASM liegt als lysosomale Form in allen Geweben vor, wird aber gleichzeitig auch kontinuierlich sezerniert (Kornhuber et al., 2015) und die NSM befindet sich über- wiegend im zentralen Nervensystem (Mühle et al., 2013, Rao and Spence, 1976, Stoffel et al., 2018). Beide gelten als die wichtigsten Regulatoren im Sphingomyelin- Abbau (Jenkins et al., 2009). Die alkalische Sphingomyelinase ist vor allem im Gastrointestinaltrakt aktiv ist (Duan et al., 1996) und wird in dieser Arbeit nicht weiter erläutert. 1.2.1 Saure Sphingomyelinase Die saure Sphingomyelinase (ASM) wurde als erstes genauer erforscht. Sie kommt hauptsächlich in Lysosomen vor, wo sie das Sphingomyelin der endosomalen Membranen verstoffwechselt. Die saure Sphingomyelinase (ASM) hydrolysiert Sphingomyelin zu Ceramid und Phosphorylcholin und ist somit ein wichtiger Regulator der Sphingolipidhomöostase (Jenkins et al., 2009). Die ASM arbeitet Zink- abhängig und wird außerdem als sogenannte sekretorische ASM nach extrazellulär ins Blutplasma sezerniert, wo sie das in Lipoproteinen (z.B. low density Lipoprotein, LDL) enthaltene Sphingomyelin verstoffwechselt (Schissel et al., 1998, Schissel et al., 1996). Es wurde gezeigt, dass es durch die Spaltung von Sphingomyelin in LDL zu einer erhöhten Aggregation und Einlagerung von LDL in die Gefäßwände kommt. Damit spielt die sekretorische ASM auch bei der Entstehung von Atherosklerose eine 7
Rolle (Smith and Schuchman, 2008). Des Weiteren erhöht sich die Aktivität der sekretorischen ASM bei zellulären Stressreaktionen mit erhöhten Entzündungs- mediatoren wie z.B. Tumornekrosefaktor (TNF)-alpha und Interferon (INF)-gamma. Das zeigt sich auch bei vielen Erkrankungen wie Alkoholismus (Reichel et al., 2011), Diabetes Mellitus Typ II oder einer Sepsis (Jenkins et al., 2010). Die Rolle der ASM bei Alkoholkonsum und Depression wird in anderen Abschnitten dieser Arbeit aus- reichend erläutert und deshalb hier nicht weiter beschrieben. 1.2.2 Neutrale Sphingomyelinase 2 Die Aktivität der neutralen Sphingomelinase wurde erstmals 1967 entdeckt (Schneider and Kennedy, 1967). Es sind unter den Säugetieren vier verschiedene NSM-Gene bekannt: SMPD2 (NSM-1), SMPD3 (NSM-2), SMPD4 (NSM-3) und die SMPD5 (mitochondrial assoziierte NSM) (Schneider and Kennedy, 1967). Die NSM-2 (SMPD3) ist die bisher am besten untersuchte neutrale Sphingomyelinase (Gault et al., 2010), wird im Folgenden weiter erläutert und in der hier vorliegenden Arbeit mit NSM betitelt. Sie ist ein membranassoziiertes Enzym von 71 kDa (Menck et al., 2017). Das Enzym selbst besteht aus zwei Domänen: Die membranassoziierte hydrophobe N-terminale Domäne, die an die Membran bindet und die C-terminale Domäne mit dem katalytischen Zentrum, welche in Richtung Zytosol zeigt. Die Hauptaktivität der NSM findet an der Innenseite der Zellmembran statt (Clarke et al., 2011). Das katalytische Zentrum kann erst durch einen neutralen pH-Wert und die Bindung von Mg2+- oder Mn2+- Ionen maximal arbeiten (Miura et al., 2004, Ago et al., 2006, Shamseddine et al., 2015) und wird durch ungesättigte Fettsäuren und anionische Phospholipide, insbesondere durch Cardiolipin und Phosphatidylserin in der Wirkung verstärkt (Marchesini et al., 2003, Wu et al., 2010). Die Neutrale Sphingomyelinase wird am häufigsten in Neuronen des Gehirns exprimiert (Mühle et al., 2013, Spence and Burgess, 1978, Stoffel et al., 2018). Sie katalysiert wie auch andere Sphingomyelinasen die Hydrolyse von Sphingomyelin unter Bildung von Ceramid und Phosphorylcholin. Die Abbauprodukte Ceramid und die weiteren Sphingolilpidmetaboliten sind, wie bereits oben beschrieben, gut erforschte Regulatoren vieler zellulärer Signalwege und sind an etlichen Prozessen zur Aufrechterhaltung der Gesundheit aber bei Störung dieser Regulation auch an der Entstehung von Krankheiten beteiligt (Wu et al., 2010). So spielt sie unter anderem bei der Knochenmineralisierung (Khavandgar et al., 2011), bei der Bildung von Exosomen (Chen et al., 2012, Khavandgar et al., 2011) und in der Entstehung von Krebs (Ito et al., 2009, Kim et al., 2008), wie zum Beispiel Leukämien (Kim et al., 2008) eine Rolle. Die NSM dient außerdem als Schlüsselmediator für die durch 8
zellulären Stress induzierte Erzeugung von Ceramid (Schneider and Kennedy, 1967). Auch die Funktion der NSM auf neuronaler Ebene ist bereits teilweise erforscht: Die NSM kommt ubiquitär im Körper vor, wird jedoch im ZNS am meisten exprimiert (Stoffel et al., 2018). Hier zeigte eine Studie bereits, dass sich nach systemischer Verabreichung des NSM-Inhibitors GW4869 der Ceramidgehalt im Gehirn verringerte und der Sphingomyelingehalt im Gehirn erhöhte, wohingegen sich der Ceramid- und Sphingomyleingehalt von Leber, Herz oder Skelettmuskel nicht verändert hat. Diese Ergebnisse deuteten darauf hin, dass die NSM im ZNS konstitutiv aktiv und im peripheren Gewebe ein induzierbares Enzym ist (Tabatadze et al., 2010). Weiterhin zeigte sich danach eine Beeinträchtigung der Bildung des räumlichen, nicht aber die Bildung des episodischen Gedächtnisses von Mäusen (Kalinichenko et al., 2019b, Stoffel et al., 2018, Tabatadze et al., 2010, Müller et al., 2015b). Andererseits konnte durch die NSM eine insgesamt erhöhte Erregbarkeit (stärkere Aktionspotential- frequenz und verkürzte Nachhyperpolarisationszeit) in Schnittpräparaten vom Hippo- campus sowie ein erhöhter Ceramid-Spiegel festgestellt werden. Gleichzeitig kam es zu einer erhöhten Informationsverarbeitung im Hippocampus (Norman et al., 2010). Man fand außerdem heraus, dass das bei der Aktivität von NSM entstehende Ceramid beim neuronalen Zelltod beteiligt zu sein scheint (Shamseddine et al., 2015). In Studien wurde gezeigt, dass Ethanol die NSM aktiviert, um über die ERK (extracellular-signal regulated kinase)-, JNK (c-Jun N-terminale Kinase)- und p38- MAP-Kinase-Wege in Astrozyten den Zelltod von Gliazellen zu erzeugen (Pascual et al., 2003). Außerdem konnte Wasserstoffperoxid (H2O2) durch Aktivierung von NSM die Apoptose von Oligodendrozyten erzeugen, was wiederum zur Verschlimmerung der Multiplen Sklerose beitrug, einer demyelinisierenden Autoimmmunerkrankung (Jana and Pahan, 2007). Auch an der Pathogenese von Morbus Alzheimer scheint die NSM beteiligt zu sein: Es konnte gezeigt werden, dass das Aβ25–35-Peptid die NSM-Aktivität in primären Astrozyten und Neuronen von Ratten hochregulierte und dort zur Apoptose führte (Ayasolla et al., 2004, Ju et al., 2005). Auch wird vermutet, dass die NSM einen Einfluss auf die Sekretion von Aβ-Peptid durch Exosomen hat. Wenn diese Exosomen aus Mikroglia freigesetzt werden, dann wird die Aβ- Oligomerisierung verhindert und gleichzeitig führt das Fehlen dieser durch NSM freigesetzten Exosomen zu einem löslichen Aβ-Peptid, welches wiederum den Zelltod von Neuronen verursacht (Dinkins et al., 2014, Wang et al., 2012, Yuyama et al., 2012). Auch kam es nach Ischämien im Gehirn durch eine erhöhte NSM Aktivität zu einer erhöhten Ausschüttung proinflammatorischer Zytokine wie TNF-α, Inter- leukin (IL)-1 und Interleukin (IL)-6 in Astrozyten (Gu et al., 2013). Insgesamt deuten diese Ergebnisse auf eine wichtige Rolle der NSM bei neuronalen Pathologien hin. 9
1.3 Alkoholkonsum und Alkoholsucht 1.3.1 Übersicht Der schädliche und unangemessene Konsum von Alkohol ist ein weltweites Problem von Menschen jeden Alters. Alkohol ist gerade in westlichen Industrieländern eine weit verbreitete psychoaktive Droge (Heath, 2000). Der regelmäßige Konsum von Alkohol kann zu einer Alkoholkonsumstörung und zu Alkoholsucht führen (European Monitoring Center for Drugs and Drug Addiction (EMCDDA), 2012). Durch den schädlichen Alkoholkonsum kommt es jährlich zu etwa 2,5 Millionen Todesfällen. Alkoholkonsum zählt als weltweit drittgrößter Risikofaktor für Krankheiten und Behinderungen, bei Ländern mit mittlerem Einkommen ist es sogar der größte Risikofaktor (World Health Organization (WHO), 2018). Mit nahezu 4 % aller Todesfälle weltweit liegt Alkoholkonsum als Todesursache vor HIV/AIDS, Gewalt oder Tuberkulose. Besonders bedrohlich einzuschätzen ist der Alkoholkonsum für Männer, weil dieser den Hauptrisikofaktor für den Tod zwischen 15 und 59 Jahren aufgrund von Herz-Kreislauf-Erkrankungen, Gewalt und Verletzungen darstellt. Alkoholismus bei Männern ist wesentlich häufiger als bei Frauen (7,4 % verglichen mit 1,4 %), was auch dadurch unterstützt wird, dass Männer weniger häufig abstinent werden als Frauen (Schuckit, 2009). Auch die auf Alkohol zurückzuführenden Todesfälle liegen mit 6,2 % weltweit bei Männern über denen der Frauen (1,1 %) (World Health Organization (WHO), 2018, Ezzati et al., 2004, Rehm et al., 2007, Rehm et al., 2003, Schuckit, 2009). Zu den häufigsten und frühesten Folgen von schädlichem Alkoholkonsum gehört die Entstehung einer alkoholischen Fettleber und die Hyperlipidämie (Reichel et al., 2015). Er kann außerdem zu somatischen und neuropsychiatrischen Erkrankungen führen (Room et al., 2005), zu denen Herz-Kreislauf-Erkrankungen wie Herzinfarkt, Schlaganfall und Kardiomyopathie, Magen-Darm-Erkrankungen wie Leberzirrhose, Hepatozelluläres Karzinom und neuropsychiatrische Erkrankungen wie Epilepsie, Depression und Angststörungen gehören. Außerdem kann es durch schädlichen Alkoholkonsum zu kognitiven Defiziten, Schlafproblemen und peripherer Neuropathie kommen (Room et al., 2005, Schuckit, 2009, Urbano-Marquez et al., 1989). 1.3.2 Alkohol und der Sphingolipidstoffwechsel Durch die regelmäßige Einnahme von psychoaktiven Drogen wie Alkohol kann sich eine Abhängigkeit entwickeln. Hat sich die Alkoholsucht erst einmal entwickelt, so stellt diese eine schwere psychiatrische Störung dar, die sich in vielerlei Hinsicht pathologisch auf den Organismus auswirkt. Das Problem der Suchterkrankung ist, dass es bis heute keine befriedigende Behandlungsstrategien und nur unzureichende 10
Pharmakotherapien gibt (Kalinichenko et al., 2018). Als ein möglicher Ansatz in der Pathogenese und schließlich auch als ein therapeutischer Ansatz in der Alkoholsucht gelten die Sphingolipide und deren Homöostase. Veränderungen des Sphingolipid-Metabolismus stehen im Zusammenhang mit dem Auftreten von Depressionen, Angstzuständen, kognitiven Beeinträchtigungen, kardiovaskulären Risiken, Adipositas, Gewichtsverlust und Sepsis (Sigruener et al., 2014, Heimerl et al., 2014, Drobnik et al., 2003, Demirkan et al., 2013, Gracia-Garcia et al., 2011, Mielke et al., 2010a, Mielke et al., 2010b). All diese Krankheiten können auch mit starkem Alkoholmissbrauch assoziiert sein. Gerade die psychischen Erkran- kungen Alkoholsucht, Depression und Angststörung treten häufig komorbide auf (Müller et al., 2017, Pettinati et al., 2013). Frühere Forschungen zeigten, dass Alkohol die saure und neutrale Sphingo- myelinase (ASM und NSM) in vitro und in vivo aktiviert (Deaciuc et al., 2000, Fernandez et al., 2013, Liangpunsakul et al., 2012, Liangpunsakul et al., 2010, Liu et al., 2000, Pascual et al., 2003, Reichel et al., 2011, Reichel et al., 2010, Saito et al., 2005, Kornhuber et al., 2015). Hierbei beobachtete man unter anderem in Studien mit alkoholabhängigen Patienten, dass akuter und chronischer Alkoholkonsum zu einer Aktivierung der ASM führt. Auch bei Patienten mit einer akuten Alkoholvergiftung kam es zu einem signifikanten Anstieg der ASM-Aktivität in peripheren Blutzellen. Dieser normalisierte sich während der Abstinenzzeit wieder (Reichel et al., 2011, Reichel et al., 2010). Umgekehrt führte auch eine vermehrt aktive ASM zu einem erhöhten Alkoholkonsum bei Mäusen und eine genetisch vermindert exprimierte ASM zu einem reduzierten Alkoholkonsum (Müller et al., 2017). Bei Mäusen, die mit Alkohol gefüttert wurden, zeigten diese außerdem einen signifikanten Anstieg der Ceramide und deren Vorläufern Sphingosin und Sphinganin in der Leber (Clugston et al., 2011) und in anderen Organen (Mühle et al., 2013). Wie bereits im Kapitel 1.1 „Sphingolipide und Sphingomyelinasen“ beschrieben, hydrolysieren Sphingomyelinasen das Sphingomyelin in Ceramid und Phosphoryl- cholin (Schneider and Kennedy, 1967). Erhöhte Ceramidspiegel wiederum führen zu oxidativem Stress in Zellen (Nikolova-Karakashian and Rozenova, 2010, Pascual et al., 2003, Hannun and Obeid, 2008) und wurden in diesem Zusammenhang in erhöhten Spiegeln in der Leber gemessen (Clugston et al., 2011, Deaciuc et al., 2000, Liangpunsakul et al., 2010, Zhao et al., 2011). Oxidativer Stress im Zusammenhang mit erhöhten Ceramidspiegeln wiederum kann zu Leberschäden und auch Schäden des zentralen Nervensystems führen (García-Ruiz et al., 2003, Pascual et al., 2003). In anderen experimentellen Studien mit Mausmodellen, bei denen Ceramide genetisch und pharmakologisch gehemmt wurden, konnte die Verminderung von 11
Entzündungen, der alkoholinduzierten Fettleber, des Leberzelluntergangs und der gestörten Glucosetoleranz festgestellt werden (Deaciuc et al., 2000, García-Ruiz et al., 2003, Liangpunsakul et al., 2012). Demnach liegt ein Zusammenhang von Sphingolipidstoffwechsel und Alkoholismus nahe. 1.3.3 NSM und Alkohol Auch die neutrale Sphingomyelinase spielt bei der Alkoholkonsumstörung eine Rolle. Dies wurde bereits in Zellkulturen, Mausmodellen und bei alkoholabhängigen Patienten beobachtet (Deaciuc et al., 2000, Fernandez et al., 2013, Liangpunsakul et al., 2012, Liangpunsakul et al., 2010, Liu et al., 2000, Pascual et al., 2003, Reichel et al., 2011, Reichel et al., 2010, Saito et al., 2005, Kornhuber et al., 2015). Beobachtet wurde unter anderem, dass es durch eine dosisabhängige Alkoholexposition zur Apoptose von Leberzellen und damit begleitend zu einer erhöhten Aktivität der NSM kam (Liu et al., 2000). Auch bei Astrozyten führte Ethanol zum Zelltod und dieser ging mit der Stimulation von ASM und NSM sowie mit erhöhten Ceramidspiegeln einher (Pascual et al., 2003). Dennoch ist die genaue Rolle, welche die NSM im Zusammen- hang mit Alkoholsucht spielt, bis heute nicht ausreichend verstanden. 1.4 Depression 1.4.1 Übersicht Die Depression zählt zu den affektiven Störungen mit einer Lebenszeitprävalenz von 16-20 %. Sie belegt damit den vierten Platz der globalen Krankheitslast, wird vermutlich bis 2030 weltweit auf dem zweiten und in Ländern mit hohem Einkommen sogar auf dem ersten Platz liegen (Mathers and Loncar, 2006). In Deutschland erkranken innerhalb eines Jahres ca. 6,2 Mio. Menschen an einer unipolaren Depression bei einer 1-Jahres-Inzidenz von 7,7 % (Jacobi et al., 2014). Sie kann in jedem Alter auftreten, die Erstmanifestation liegt jedoch im Mittel zwischen dem 18. und 25. Lebensjahr (Medizinische Fachredaktion Pschyrembel, 2020). Obwohl die Depression bereits jetzt einen hohen Stellenwert in der Gesundheitsversorgung einnimmt (Lopez et al., 2006, Ustün et al., 2004), wird ihre Bedeutung für die Gesellschaft und für die individuelle Person weiterhin noch sehr unterschätzt (Ärztliches Zentrum für Qualität in der Medizin (ÄZQ), 2019, Lopez and Murray, 1998). Zu den Hauptsymptomen einer Depression zählen ein Interessensverlust, ein verminderter Antrieb und eine gedrückte Stimmungslage. Zur Diagnosestellung nach ICD-10 müssen zwei der Hauptsymptome für mindestens zwei Wochen andauern. Weitere Symptome, die Zusatzsymptome genannt werden, sind unter anderem Konzentrationsstörungen, vermindertes Selbstwertgefühl, Appetit- und damit einher- 12
gehender Gewichtsverlust, Schlafstörungen, Morgentief und Libidoverlust (DGPPN (Deutsche Gesellschaft für Psychiatrie und Psychotherapie, 2017, DIMDI Deutsches Institut für Medizinische Dokumentation und Information, 2017, World Health Organization (WHO), 1993). Die Depression wird je nach Anzahl der Zusatz- symptome in eine leichte, mittelgradige und schwere Depression unterteilt (DIMDI Deutsches Institut für Medizinische Dokumentation und Information, 2017). Der Krankheitsverlauf ist typischerweise episodisch und weist nicht selten Komorbiditäten mit anderen psychiatrischen Erkrankungen auf (World Health Organization (WHO), 2008). Dazu gehören unter anderem eine erhöhte Suizidalität, die generalisierte Angststörung und die Alkoholsucht (Lenz et al., 2019). Obwohl die Erkrankung so häufig ist und deshalb auch bereits als Volkskrankheit betitelt wird, ist die genaue Pathogenese bis heute nicht vollständig geklärt (World Health Organization (WHO), 2008). Die Ätiologie wird durch mehrere Theorien erklärt. Dabei spielen genetische Faktoren sowie einschneidende Lebensabschnitte und -ereignisse als multifaktorielle Auslöser eine Rolle. Ein weit verbreiteter Erklärungsansatz ist die Monoaminhypothese, nach der eine Depression neurobiologisch durch eine Verringerung der Neurotransmitter erklärt wird, insbesondere durch verringerte Serotonin-, Noradrenalin- und Dopaminlevel (Duval et al., 2000, Kornhuber et al., 2014). Gestützt wird diese Hypothese durch die Funktionsweise und den Erfolg der bisher bekannten Antidepressiva. Dazu gehören unter anderem die selektiven Serotonin Rückaufnahme Hemmer (SSRI’s) Sertralin und Citalopram. Sie hemmen die Wiederaufnahme von Serotonin aus dem synaptischen Spalt zurück in die Präsynapse, sodass die biologische Verfügbarkeit von Serotonin in diesem erhöht wird und infolgedessen die depressiven Symptome abnehmen (Ustün et al., 2004). Die Monoaminhypothese ist jedoch als alleinige Erklärung für die Entstehung der Depression nicht ausreichend. So zeigen SSRI’s zwar eine antidepressive Wirkung, diese tritt allerdings erst zeitverzögert ein, obwohl der Serotoninspiegel schon nach kurzer Zeit messbar erhöht ist (Kornhuber et al., 2014, Kornhuber et al., 2010, Kornhuber et al., 2008). Aufgrund dessen sind die SSRI's nicht bei akut und schnell behandlungsbedürftigen psychiatrischen Anlässen wie beispielsweise bei einer akuten Panikattacke oder einer akuten Psychose indiziert. Zusätzlich wurde festgestellt, dass man bei gesunden Patienten eine Depression nicht durch einen erzeugten Monoamin-Mangel (Serotonin und Noradrenalin) induzieren kann. Lediglich bei bereits mit SSRI‘s behandelten Patienten konnte durch erzeugten Monoamin-Mangel ein Rückfall in die Depression induziert werden (Ruhé et al., 2007). Aus diesen Gründen muss es noch andere Mechanismen geben, die bei der Pathogenese der Depression eine Rolle spielen. 13
1.4.2 Depression und der Sphingolipidstoffwechsel Eine weitere mögliche Erklärung der Pathogenese der Depression beruht auf einem veränderten Sphingolipidstoffwechsel. Verschiedene Studien zeigen, dass es einen Zusammenhang zwischen einem veränderten Sphingomyelin- und Ceramidstoff- wechsel und dem Auftreten einer Depression gibt. Gerade über die ASM ist in diesem Zusammenhang schon viel bekannt. Hierzu wurde die ASM/Ceramid-Hypothese der Depression aufgestellt. Grundlage dieser ist die Erkenntnis, dass die ASM und die Ceramidspiegel im Plasma von Patienten mit einer schweren depressiven Episode erhöht sind (Kornhuber et al., 2005), verglichen mit einer Kontrollgruppe (Gracia- Garcia et al., 2011). Es zeigte sich zudem bei Patienten mit einer schweren depressiven Episode eine erhöhte ASM-Aktivität in mononukleären Zellen des peripheren Bluts (Kornhuber et al., 2005), diese korrelierte mit dem Ausmaß des klinischen Bildes der Depression und führte außerdem zu einem Anstieg der proinflammatorischen Zytokine IL-1, IL-6 und TNF-α (Kornhuber et al., 2005, Lang and Borgwardt, 2013). Auch umgekehrt konnte gezeigt werden, dass bei Mäusen mit einer überexprimierenden ASM (tgASM) ein depressives Verhalten vorlag (Müller et al., 2017). Ein depressiver und anxiogener Phänotyp wurde insbesondere bei weiblichen tgASM-Mäusen beobachtet (Zoicas et al., 2016). Der Einfluss von Sphingolipiden auf das Krankheitsbild der Depression konnte eine weitere Studie verdeutlichen, in der es durch eine direkte Injektion von C16-Ceramid in den dorsalen Hippocampus von naiven Mäusen zu einer erhöhten ASM-Aktivität im Serum und im Hippocampus kam. Gleichzeitig zeigte sich eine Zunahme der Ceramidkonzentration und des depressiven Verhaltens sowie eine Abnahme der neuronalen Proliferation (Gulbins et al., 2013, Müller et al., 2017, Zoicas et al., 2016, Zoicas et al., 2020a). Auch in medikamentöser Hinsicht wurde ein Mitwirken des Sphingolipidhaushalts bei antidepressiver Therapie festgestellt. Es handelt sich dabei um die sogenannten FIASMAs. Dies ist ein Akronym und leitet sich von der englischen Bezeichnung Functional Inhibitor of Acid SphingoMyelinAse ab (Kornhuber et al., 2010). Viele Anti- depressiva wie Amitriptylin, Desipramin, Fluoxetin, Flupentixol, Fluphenazin, Imipramin, Nortriptylin, Paroxetin und Sertralin sind aktuell bekannte FIASMAs (Kornhuber et al., 2011, Kornhuber et al., 2010, Kornhuber et al., 2008). Diese Pharmaka führen zu einer funktionellen Hemmung der ASM durch Induktion eines partiellen Abbaus dieser. Es kommt dabei zu einer Ablösung der ASM von der inneren Membran des Lysosomens, in welcher die ASM vorwiegend zu finden ist. Dadurch kommt es schließlich im Lumen des Lysosomens zum proteolytischen Abbau und damit zum Wirkungs- und Funktionsverlust der ASM. Die vorher erhöhten ASM- Spiegel normalisieren sich und die depressiven Symptome werden besser 14
(Kornhuber et al., 2011, Kornhuber et al., 2010, Kornhuber et al., 2008). Die Hemmung selbst geschieht somit indirekt (Kölzer et al., 2004). Studien bestätigten die obige Erkenntnis und zeigten nach Gabe der Antidepressiva Amitriptylin und Fluoxetin eine erniedrigte ASM-Aktivität und Ceramidkonzentration im Hippocampus sowie eine erhöhte neuronale Plastizität (Gulbins et al., 2013). Außerdem kam es bei genetisch herunterregulierter ASM zu gleichen Effekten wie nach der Gabe von Antidepressiva und man konnte prognostisch je nach Ausmaß der ASM-Aktivität auf eine Besserung der depressiven Symptome durch eine medikamentöse Therapie schließen (Mühle et al., 2019). All dies bestärkt die ASM/Ceramid-Hypothese. Eine gewisse Restaktivität der ASM bleibt trotz funktioneller Hemmung der ASM durch FIASMAs erhalten. Es kam durch Gabe von FIASMAs nicht zum klinischen Bild des Morbus Niemann-Pick, einer autosomal-rezessiv vererbten Lipidspeicherkrankheit mit fehlerhafter ASM (Kornhuber et al., 2010, Sakuragawa et al., 1977). Ungeachtet dieser eben beschriebenen Erkenntnisse über die ASM und die Ceramide hinsichtlich emotionalen Verhaltens ist weiterhin wenig über die Beteiligung der anderen Sphingomyelinasen in der Pathophysiologie der Depression und Angst bekannt (Kalinichenko et al., 2019c). 1.4.3 NSM und Depression Die im Gehirn stark exprimierte NSM (Mühle et al., 2013, Spence and Burgess, 1978) und die dadurch entstehenden Ceramide spielen eine entscheidende Rolle beim emotionalen Verhalten (Gulbins et al. 2013, 2018; Zoicas et al. 2019). So zeigte sich erst kürzlich eine erhöhte NSM-Aktivität mit ängstlichem Verhalten im dorsalen Mesencephalon von weiblichen Ratten, die von Geburt an ein ängstliches Verhalten aufwiesen. Diese Hirnregion ist mit ängstlichem und depressivem Verhalten assoziiert (Zoicas et al., 2020b). Eine In vitro-Studie zeigte außerdem eine gehemmte Interferon-α-induzierte Serotonin-Aufnahme in T-Zellen nach pharmakologischer Hemmung der NSM durch Sphingolacton-24 (Su et al., 2011). Durch die damit indirekte Erhöhung des Serotonins könnte es zu einem antidepressiven Effekt durch NSM-Hemmung kommen (Müller et al., 2015a). 1.5 Der NSM-Inhibitor GW4869 GW4869 ist ein nichtkompetitiver Inhibitor der neutralen Sphingomyelinase 2. Die NSM ist damit bisher die einzige Sphingomyelinase, bei der ein spezifischer Inhibitor existiert (Canals et al., 2011, Luberto et al., 2002). Appliziert wird der NSM-Inhibitor intraperitoneal (i.p.) oder subkutan (s.c.), da eine direkte Injektion ins Gehirn sehr 15
wahrscheinlich zu Entzündungen und Schädigungen des Gewebes führen würde (Tabatadze et al., 2010, Takahashi et al., 2006). Die mittlere inhibitorische Konzentration IC(50) liegt bei 1 μM (Luberto et al., 2002) und gibt die nötige GW4869- Konzentration an, mit welcher die NSM in vitro zu 50 % gehemmt wird (Lumitos AG, 1997-2021). GW4869 wurde bereits in vielen in vitro- und in vivo-Studien (Luberto et al., 2002) zur Untersuchung von NSM-spezifischen Funktionen verwendet (Wu et al., 2010). Man konnte bei einer täglich i.p. injizierten GW4869-Dosis von 1,25 mg/kg keine unge- wünschten Nebenwirkungen feststellen. GW4869 überwand bei dieser Dosis und Applikationsform in hinreichendem Ausmaß die Blut-Hirn-Schranke und konnte so die NSM-Aktivität im Gehirn ausreichend stark hemmen. Diese medikamentenindu- zierten Veränderungen im Sphingolipidgehalt des Gehirns waren denen der hetKO- Mäuse (fro), mit genetisch bedingt verminderter NSM-Aktivität, sehr ähnlich. Das deutete darauf hin, dass GW4869 effektiv und gezielt auf die NSM abzielt (Tabatadze et al., 2010). Es wurde außerdem festgestellt, dass die Auswirkungen von GW4869 auf Sphingomyelin und Ceramidgehalt am auffälligsten im Hippocampus waren, wo die NSM auch ein sehr starkes Expressionsmuster aufweist (Hofmann et al., 2000). Auch diese Ergebnisse deuten darauf hin, dass GW4869 ein wirksamer Inhibitor der NSM ist und dass die NSM an der konstitutiven Regulation des Ceramidstoffwechsels im Gehirn beteiligt sein könnte (Tabatadze et al., 2010). Bei menschlichen Brustkrebszellen (MCF7) konnte in vitro gezeigt werden, dass nach einer GW4869-Behandlung mit einer Konzentration von 10-20 μM und Hinzugabe von TNF eine erheblich und bei 20 μM GW4869 sogar eine vollständig gehemmte TNF-abhängige Sphingomyelin-Hydrolyse stattfand. GW4869 schützte außerdem dosisabhängig vor dem Zelltod (Luberto et al., 2002) und hemmte die Ceramid- vermittelte Entstehung von multivesikulären, späten Endosomen (multivesicular body, MVB) und die Freisetzung von reifen Exosomen aus späten Endosomen (MedChemExpress, 2013-2020, Luberto et al., 2002, Chen and Brigstock, 2016, Essandoh et al., 2015, Nakamura et al., 2015). Auch bei RAW264.7 Makrophagen, die mit 10 bzw. 20 μM GW4869 vorbehandelt und dann mit Lipopolysacchariden (LPS) inkubiert wurden, zeigte sich eine starke Abschwächung der LPS-induzierten Exosomen-Feisetzung und der proinflamma- torischen Zytokinproduktion (Essandoh et al., 2015). Es konnte auch gezeigt werden, dass nach GW4869-Behandlung die Induktion der Apoptose in Krebszellen blockiert wird (Menck et al., 2017). In vivo konnten ähnliche Effekte wie bei den in vitro behandelten Makrophagen gezeigt werden. Bei 10-12 Wochen alten männlichen Wildtyp-C57BL/6-Mäusen, 16
welche 2,5 μg/g GW4869 i.p. injiziert bekamen, gefolgt von einer i.p. LPS-Injektion (zur Stimulation der LPS-induzierten Exosomenfreisetzung) zeigte sich, dass GW4869 die LPS-induzierte Exosomenfreisetzung sowie die LPS-stimulierte pro- inflammatorische Zytokinproduktion und Herzentzündung bei diesen Mäusen hemmte. Auch die LPS-bedingten Myokardfunktionsstörungen wurden gemildert und das Überleben der Mäuse verbessert (Essandoh et al., 2015). 17
Zielsetzung und Hypothesen der vorliegenden Arbeit Ziel der vorliegenden Arbeit lag in der Erforschung des Zusammenhangs von Alkoholsucht, Depression und Angststörungen sowie dessen Geschlechtsspezifität im Hinblick auf die Regulation durch die NSM. Wie bereits beschrieben, sind Sphingolipide als Bestandteil der Zellmembran ubiquitär in unserem Körper vorhanden. Das absolute und relative Verhältnis der Sphingolipide und Ceramide in der Zellmembran wird über verschiedene Pathways reguliert und kann über Signalmoleküle und Signalkaskaden zu sichtbaren physio- logischen Veränderungen führen (Hannun and Obeid, 2008). Hierdurch formte sich der Grundgedanke, dass der Sphingolipidstoffwechsel eine wesentliche Rolle in der Pathogenese von psychiatrischen Erkrankungen wie der Alkoholsucht, der schweren Depression und der Angststörung spielt. Die NSM wird fast ausschließlich im Gehirn exprimiert (Mühle et al., 2013, Spence and Burgess, 1978) und wurde bereits im Zusammenhang mit den psychiatrischen Erkrankungen Alkoholsucht (Deaciuc et al., 2000, Fernandez et al., 2013, Liangpunsakul et al., 2012, Liangpunsakul et al., 2010, Liu et al., 2000, Pascual et al., 2003, Reichel et al., 2011, Reichel et al., 2010, Saito et al., 2005, Kornhuber et al., 2015), Depression und Angststörung beschrieben (Zoicas et al., 2020b). Ausgehend von diesen Überlegungen sollten hier im pharmakologischen Modell der NSM-Inhibition durch GW4869 einerseits das freiwillige Alkoholtrinkverhalten und andererseits die Emotionalität in Form von Angst- und Depressionsverhalten im Mausmodell untersucht werden. Beide Experimente erfolgten gegenüber einer mit Saline (0,9 % NaCl) behandelten Kontrollgruppe sowie geschlechtergetrennt mit je einer Gruppe männlicher und einer Gruppe weiblicher pharmakologisch mit GW4869 NSM-gehemmter Mäuse. Die in diesem Zusammenhang aufgestellten Hypothesen lauten: 1. Der NSM-Inhibitor GW4869 verändert das Alkoholtrinkverhalten bei Mäusen. 2. Der NSM-Inhibitor GW4869 verändert das Angst- bzw. Depressionsverhalten bei Mäusen in entsprechenden Verhaltensparadigmen. 3. Beide oben genannten GW4869-Effekte sind geschlechtsabhängig. 18
Sie können auch lesen

















































