Taxonomische Beiträge zur Myrmekofauna Jordaniens (Hymenoptera: Formicidae)
←
→
Transkription von Seiteninhalten
Wenn Ihr Browser die Seite nicht korrekt rendert, bitte, lesen Sie den Inhalt der Seite unten

© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
Taxonomische Beiträge zur Myrmekofauna
Jordaniens (Hymenoptera: Formicidae)
CO. DIETRICH
Abstract: Contributions to the Myrmecofauna of Jordan (Hymenoptera: Formicidae). Three new
species of ants from Jordan are described: Tetramorium argentirubrum n. sp. (Tetramorium striativentre-
group), Camponotus husseini n. sp. (Camponotus sericeus-group) and Polyrhachis palaearctica n. sp.
(Polyrhachis dives-group). Some oriental ants require a new status: Camponotus opaciventris n. stat. (till
now Camponotus sericeus opaciventris), Polyrhachis grisescens n. stat. (till now Polyrhachis simplex gris-
escens), Polyrhachis spiniger n. stat. (till now synonym of Polyrhachis lacteipennis resp. Polyrhachis simplex).
The taxonomic status of Polyrhachis hauxiuelli and the taxonomical complex situation of the Campono-
tus sericeus-group are critically discussed. The Zoogeographie distribution of Polyrhachis (Myrmhopla) in
South Western Asia and of the three new species from Jordan is represented. The paper also deals with
the morphological differences between Lepisioto gracilicornis and hepisiota opaciventris and with func-
tional morphological aspects of the genus Lepisiota.
Key words: ants, Formicidae, Jordan, taxonomy, zoogeography, functional morphology, arid environ-
ments
Einleitung den verbreitete Ameisenart Jordaniens ist
Colobopsis truncata. Umgekehrt scheint Jor-
Der Raum entlang des Jordangrabens ist
danien eine umfangreiche, endemische
trotz seiner geringen Ausdehnung zoogeo-
Steppenfauna zu beherbergen, wie die Be-
graphisch bemerkenswert, weil hier Elemen-
schreibung dreier neuer Ameisenarten (zwei
te der Äthiopis und der Orientalis auf eine
Cardiocondyla-Arten SEIFERT 2003; Tetramo-
paläarktische Fauna treffen. Das gilt natür-
rium argentirubrum n. sp.) aus dem Shauma-
lich auch für Ameisen und wurde von BY-
ri Wildlife Reserve andeuten.
TINSKI-SALZ (1953) und KUGLER (1988) dar-
gestellt. Während für das Gebiet westlich Das ursprüngliche Vorhaben einer er-
des Grabens entlang der Mittelmeerküste sten Darstellung der jordanischen Myrme-
(Sinai, Israel, Libanon) Artenlisten vorlie- kofauna scheiterte an der schwierigen taxo-
gen (TOHME 1969, KUGLER 1988), ist Jorda- nomischen Grundsituation. Die Ameisenta-
nien, bis auf vereinzelte Notizen (z. B. xonomie dieses Raums bezieht sich noch
WHEELER &. MANN 1916), myrmekologisch hauptsächlich auf Arbeiten vor dem 2.
unerforscht. Die Ameisenfauna westlich des Weltkrieg. In jüngerer Zeit sind allerdings
Grabens mit jener östlich des Grabens an- aus dieser Region etliche neue Arten be-
nähernd gleich zu setzen, wäre falsch. Der kannt geworden (Aphaenogaster ledouxi, Te-
Gradient von Küste zur Wüste vollzieht sich tramorium arzi, Camponotus nadimi TOHME
in diesem Raum so rasch, dass sich die fau- 1969; Belonopelta loebii BARONI URBANI
nistische Situation Jordaniens anders dar- 1975; Amblyopone ophthalmica BARONI U R -
stellt. Beispielsweise fehlen in Jordanien die BANI 1978; Epixenus grassei, Epixenus syriaca,
für die Paläarktis qualitativ und quantitativ Epixenus Ubanicus TOHME & TOHME 1979;
so bedeutsamen Gattungen Lasius und for- Soknopsis fugax emesus, Solenopsis fugax kara-
mica, während sie im Libanon (Formica, La- ki TOHME & TOHME 1980; Messor nahaU
sius) und Israel (Lasius) zumindest ansatz- THOME & THOME 1981; Cataglyphis sabulosa Denisia 14, zugleich Kataloge
weise auftreten. Die einzige, weit nach Nor- der OÖ. Landesmuseen
KUGLER 1981; Yavneüa argamani, Leptaniüa Neue Serie 2 (2004), 319-344
319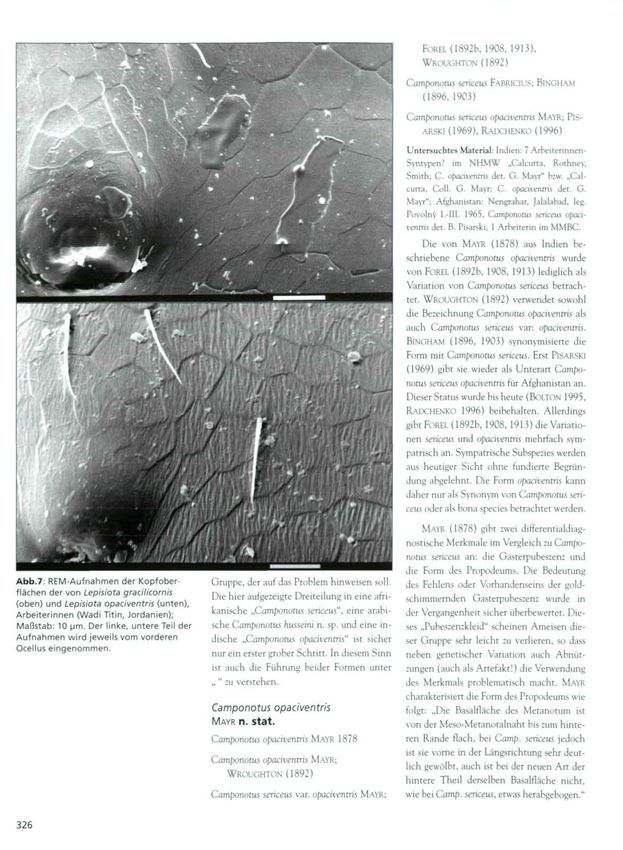
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
Schritt sind die folgenden Beiträge zur Te-
tramarium striativentre-, Lepisiota frauenfeldi-
und Camponotus sericeus-Gruppe, sowie zu
Polyrhachis (Myrmhopla) zu verstehen. Die
zoogeographische Sonderstellung entlang
des Grabenbruchs im Nahen Osten erfor-
dert die Berücksichtigung von Orientalis
und Äthiopis. Soweit es der derzeitige
Kenntnisstand zulässt, werden auch biologi-
sche Unterschiede oder Besonderheiten
dargestellt und die Zoogeographie der neuen
Arten erörtert.
Material und Methoden
Das verwendete Material stammt von
folgenden Institutionen und Privatsamm-
lungen (Abkürzungs-Codes folgen, soweit
vorhanden, ARNETT et al. 1993):
NHMW Naturhistorisches Museum
Wien (Österreich)
NHMB Naturhistorisches Museum
Basel (Schweiz)
TAU1 Zoologisches Museum der
Universität Tel Aviv (Israel)
A b b . 1: Verbreitung von Tetramorium MMBC Mährisches Museum in Brunn
argentirubrum n. sp. (voller Kreis), Camponotus (Tschechien)
husseini n. sp. (leerer Kreis) und Polyrhachis
palaearctica n. sp. (Kreuz) in Jordanien, sowie Über- MZLU Zoologisches Museum der
sicht besammelter Standorte (CCD) ohne diese Arten (-). Universität Lund (Schweden)
LMO Oberösterreichisches Landes-
museum (Linz, Osterreich)
bifurcata, Leptanitia Israelis, Leptaniüa judaica
KUGLER 1986; Anochetus n. sp., Stenamma n. CCC Coll. Cedric A. Collingwood
(Leeds, Großbritannien)
sp., Aphaenogaster n. sp., Goniomma n. sp.,
Acamholepis n. sp., Cataglyphis n. spp. KuG- CCD Coll. Christian O. Dietrich
LER 1988; Solenopsis dentata COLLINGWOOD (St. Polten, Österreich)
& KUGLER 1994; Cataglyphis sp., Cataglyphis CKW Coll. der Gesellschaft zur
holgerseni COLLINGWOOD & AGOSTI 1996; Kartierung der Wirbellosen
Cataglyphis cf. mauretanica COLLINGWOOD in Vorarlbergs (Bürs, Österreich)
litt. 1999, oder es wurden nomenklatorische CWW Coll. Wolfgang Wranik
Änderungen vorgenommen. Gleichzeitig (Rostock, Deutschland)
fehlen weitgehend neuere, für das betreffen- Das Material aus CCD stammt von 2-4
de Gebiet anwendbare Gattungsrevisionen. wöchigen Jordanienexkursionen im Früh-
Diese Umstände zeigen, dass die Determina- jahr 1996, Herbst 1996 und Frühjahr 1998
tion der jordanischen Ameisen teilweise ei- durch direkte Suche nach Ameisen und de-
ne taxonomische Grundlagenarbeit erfor- ren Nestern (Abb. 1). Die rasterelektronen-
dert. Das Beispiel von Cataglyphis holgerseni mikroskopischen Untersuchungen (REM-
COLLINGWOOD & AGOSTI 1996 (locus typi- Aufnahmen) erfolgten an einem JEOL
cus nahe Eilat), der bis dahin mit Catagly- Scanning Microscop JSM-35CF, durchge-
phis niger verwechselt wurde, zeigt, dass führt an der Abteilung für Ultrastrukturfor-
selbst große, auffallende Formen sehr kri- schung des Institutes für Zoologie (Univer-
tisch zu betrachten sind. Die Abklärung der sität Wien). Die Proben wurden luftgetrock-
Myrmekofaunistik Jordaniens kann also nur net, mit Gold besputtert (Sputtercoater
schrittweise erfolgen und als ein solcher HUMMER JR) und bei 15 kV untersucht.
320
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
Die Berechnung des Fisher-Yates-Tests zur
exakten Lösung von Vierfelderproblemen A b b . 2: Dornenmorphometrie bei
erfolgt nach BORTZ et al. (1990). Po/y/7)ac/j;s-Arbeiterinnen (dorsolaterale
Ansicht des Mesosomas).
Für determinatorische Zwecke wurden
wurden folgende morphometrischen Merk-
male verwendet:
Lepisioux
K B . . . maximale Kopfbreite inklusive Augen
SL . . . maximale Scapuslänge
FL . . . maximale Hinterfemurlänge
Polyrhachis
K B . . . maximale Kopfbreite inklusive Augen
KL . . . mediane Kopflänge zwischen Occiput
und Clypeusvorderrand
OD. . . maximaler Durchmesser der Ocellen DSi = X/Y
A D . . . maximaler Durchmesser der Facettenau-
gen
PDW . Pronotalwinkel in Grad
(Abb. 2). Winkel zwischen den Symme- Camponotus
trieachsen der Pronotaldornen bei dorsa-
KB . . . maximale Kopfbreite inklusive Augen
ler Ansicht. Dieser Winkel darf nur in
einer bestimmten Ebene gemessen wer- P I . . . . Petiolarindex. Verhältnis zwischen Brei-
den. Als solche wurde jene Ebene defi- te (transversal) und Länge (longitudinal)
niert, bei der die Spitzen der Pronotal- der nodiformen Schuppe bei dorsaler
dornen und die Spitzen der Propodeal- Ansicht. Die Längenmessung bezieht
dornen gleichzeitig scharf zu sehen sind. sich nur auf die Schuppe und nicht auf
Die Winkelmessung erfolgte bei einer die Gesamtlänge des Petiolus. Das Tier
64-fachen Vergrößerung. muss dabei so gedreht werden, dass die
abschüssigen Flächen gleichermaßen ge-
DS1 . . Dornstigma-lndex (Abb. 2). 1st ein Maß
sehen bzw. nicht gesehen werden (vari-
um die Vertikalität der Propodealdornen
iert bei den einzelnen Individuen).
zu quantifizieren. Dabei wird das Tier la-
teral in eine Ebene gebracht, bei der die
Spitzen des Pronotal- und Propodealdor- Taxonomie
nes sowie der oberste Punkt des propode-
alen Stigmas scharf zu sehen sind. Die Tetramorium striativentre-
Strecke zwischen den Dornenspitzen Gruppe
wird durch eine Normale geteilt, die
Diese Gruppe setzt sich von den übrigen
durch den obersten Punkt des Stigmas
Tetramorium-Arten durch eine dichte,
führt. Der DSI drückt das Verhältnis die-
ser Teilstrecken aus. Bei positivem DSI längsgestrichelte Skuptur am ersten Gastert-
überragen die Propodealdornen die Pro- ergiten der weiblichen Kasten ab (RAD-
podealstigmen, bei negativen DSI nicht. CHENKO 1993).
DSI Bei der gemeinsamen Verwendung von
PDW und DSI in einer Trennfunktion Tetramorium striativentre MAYR
zur Unterscheidung zweier Gruppen Tetramorium caesphum var. striativentre
muss der DSI mit einem Faktor gewich- MAYR 1877; MAYR 1880
tet werden. Ohne Gewichtung würde die
Trennung praktisch nur auf dem PDW Tetramorium striativentre striativentre MAYR;
wegen seiner hohen Zahlen im Vergleich RUZSKY 1906, PlSARSKl 1967
zum DSI beruhen. Unter der Annahme
Tetramorium striativentre MAYR;
PDW und DSI seien gleich wichtige
RADCHENKO (1992,1993)
Merkmale, wurde der DSI so gewichtet,
dass die Steigung der gemeinsamen Re- Untersuchtes Material: Turkestan: 1 Syntypus-
gressionsgeraden der beiden zu trennen- Arbeiterin im NHMW „18980 Turkest., T. caes-
den Gruppen 1 wird. pitum v. striativentre G. Mayr, Type."
321
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
Untersuchtes Material: Afghanistan: 3 Parary-
pen-Arbeiterinnen (Alkoholmaterial) im MZLU
„Types 1496: 25-27"
MAYR (1880) vermerkte in der Fußnote:
„In der Fedtschenko'schen „Reise nach Tur-
kestan" erschien die russische Uebersetzung
dieser im J. 1874 abgeschlossenen Abhand-
lung." Es ist daher MAYR (1877) die russi-
sche Übersetzung vom ursprünglich deut-
schen Text (MAYR 1880) und nicht umge-
kehrt.
Im NHMW befinden sich zwei Exem-
plare der Tetramorium striativentre-Gruppe,
die beide von G. Mayr als „Type" von „T.
caespitum v. striativentre" etikettiert wurden.
Nach den von RUZSKY (1906) und RAD-
CHENKO (1993) angegebenen Merkmalen
lässt sich aber das Exemplar „12419 Tur-
kest." eindeutig als das langdornige Terramo-
rium schneiden EMERY 1898, das Exemplar
„18980 Turkest." ebenso eindeutig als das
kurzdornige Tetramorium striativentre MAYR
1877 identifizieren. Mayr sind also bei der
Beschreibung seines stnativentre zwei Arten
vorgelegen, und er war sich dieser beiden
Formen bewusst, ohne dem Umstand taxo-
nomische Bedeutung zu geben: „Erwähnt
möge werden, dass von dieser Varietät ein
Stück zwei sehr kurze unscheinbare Zähn-
chen am Metanotum hat, während ein an-
deres Dornen hat, welche deutlich länger als
an der Basis dick sind." (MAYR 1880). Es
wird daher unbedingt die Festlegung eines
Lectotypus notwendig. Seit RUZSKY (1906)
wird die kurzdornige Form als striativenrre,
die langdornige als schneiden bezeichnet, so
dass im Sinne der Stabilität bei der Festle-
Abb.3: REM-Aufnahmen von Tetramorium Tetramorium schneiden EMERY
argentirubrum n. sp., Arbeiterin (Shaumari
gung des Lectotypus darauf Rücksicht ge-
Reserve, Ost-Jordanien); Maßstab: 1000 um. Tetramorium caespitum Var. stnativentre nommen werden muss. Den Lectotypus aus
MAYR 1877; MAYR 1880 dem Material des NHMW zu wählen ist
Tetramorium schneiden EMERY 1898
nicht sinnvoll, da nur ein Syntypusexemplar
in einem schlechten Zustand vorhanden ist.
Tetramorium striadventre schneiden EMERY
RUZSKY (1906), PISARSKI (1967)
Tetramorium argentirubrum n. sp.
Tetramorium schneiden EMERY; RADCHENKO
(Abb. 3)
(1992, 1993)
Locus typicus: Shaumarie Wildlife Re-
Untersuchtes Material: Turkestan: 1 Syntypus-
Arbeiterin im NHMW „12419 Turkest., T. caes-
serve (Abb. 1, 4), Jordanien
pitum v. stnativentre G. Mayr, Type" Typus-Material: Holotypus-Arbeiterin,
Jordanien: Shaumari Wildlife Reserve,
Tetramorium kabulistanicum PISARSKI
21.3.1996 C O . Dietrich; - Paratypen: Jor-
Tetramorium kabulistanicum PlSARSKI 1967 danien: 15 Arbeiterinnen Shaumari Wildli-
Tetramorium kabulisumicum PlSARSKI; fe Reserve, 21.3.1996 C O . Dietrich. Der
RADCHENKO(1992, 1993) Holotypus und 6 Paratypen befinden sich im
322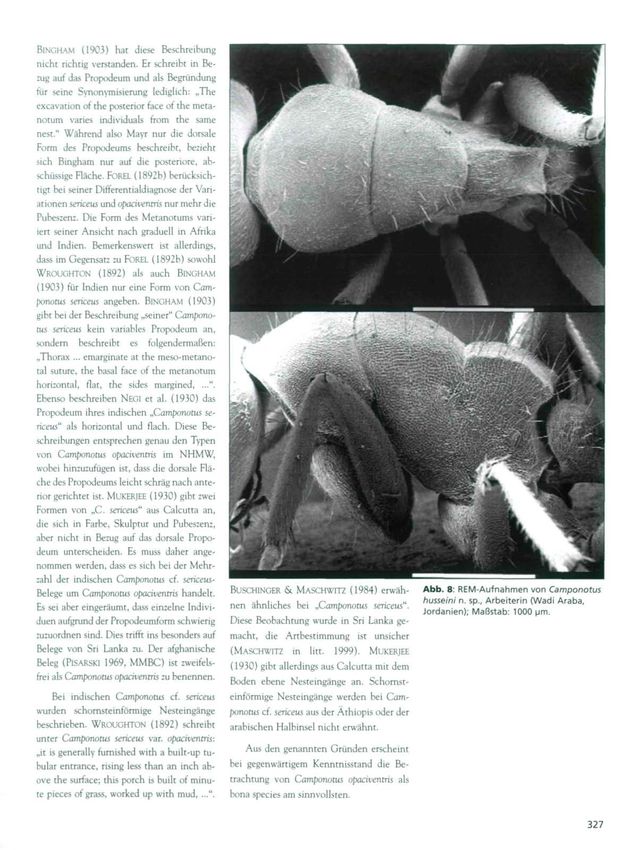
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
NHMW, je 3 Paratypen in TAUI, MMBC
und MZLU.
Untersuchtes Material: Jordanien, Shauman
Wildlife Reserve, 21.3.1996 CO. Dietrich; Wadi
Butm bei Qasr Amra, 5.11.1996 CO. Dietrich.
Etymologie: Aufgrund des silbrigen
Glanzes der Gaster infolge der Längsskulp-
tur und des roten Vorderkörpers.
Diagnose: Arbeiterinnen: Länge
3,0-3,2 mm. Kopf bis einschließlich Stiel-
chen rot, einen deutlichen Kontrast zur dun-
kelbraun bis schwarzen Gaster bildend. Kopf
median dunkler. Extremitäten basal dunkler
als Mesosoma, nach distal heller als Mesoso-
ma werdend. Gaster infolge Skulptur silbrig
glänzend. Körper ± regelmäßig abstehend
behaart. Extremitäten hauptsächlich nieder-
liegend behaart, nur Beinunterseiten mit ab-
stehenden Haaren. Kopfkapselunterseite dfri-Exemplars deutlich länger. Die Kopf- A b b . 4: Locus typicus von Tetramorium
über den ganzen Vorderrand mit ca. 5 J-för- unterseite des Syntypus von Tetramorium argentirubrum n. sp. (Shaumari Reserve,
Ost-Jordanien).
migen Haaren (Psammophore). Neben der srriarii'enrre ist nicht vollständig sichtbar,
Psammophore weiter hinten noch 2-4 subce- scheint aber in Bezug auf die Behaarung
phale abstehende Haare. Körper mit Aus- ähnlich Tetramorium schneiden sein. Tetra-
nahme der Extremitäten und hinteren Gas- morium argentirubrum ist vom Kopf bis ein-
tersegmente längsskulpturiert. Skulpturie- schließlich Stielchen rot und bildet einen
rung 1. Gastertergiten wesentlich feiner und deutlichen Kontrast zur dunkelbraunen bis
regelmäßiger als am restlichen Körper und schwarzen Gaster. Kopf median dunkler. Die
erstreckt sich über den ganzen Tergiten und 3 Paratypen von Tetramorium kabulistanicum
die lateralen Teile des Sterniten. Kopfskulp-sind ± rotbraun, der Gaster zeigt kaum einen
tur feiner als die des Mesosomas. Anzahl der farblichen Kontrast zum Vorderkörper. Dies
Längsrunzeln zwischen den verlängerten steht im leichten Widerspruch zu PlSARSK]
Stimleisten ca. 15, Petiolus und Postpetiolus(1967) und RADCHENKO (1993). Die beiden
deutlich unterschiedlich skulpturiert - Pe- anderen Arten sind einheitlich rotbraun
tiolus dorsal netzförmig, Postpetiolus längs bzw. dunkelbraun. Tetramorium kabulistani-
skulpturiert. Allgemeine Körperform siehe cum besitzt im Gegensatz zu den anderen 3
Arten deutliche Schulterecken, und die
Abb. 3. Stirn mit leichtem Eindruck. Schul-
Längsrillen des ersten Gastertergiten rei-
terecken fehlen. Metanotaler Quereindruck
chen nicht bis zum posterioren Rand. Tetra-
erkennbar. Petiolus gestielt mit ventralem
morium schneideri unterscheidet sich von
Zähnchen und schmäler als Postpetiolus.
den übrigen 3 Arten durch sehr feine, zahl-
Differentialdiagnose: Beruht auf dem reiche Längsrunzeln am Kopf und dornen-
Vergleich mit den oben genannten Typen förmig verlängerte metastemale Zähne (Ter-
von Tetramorium striativentre, Tetramorium minologie nach RUZSKY 1906). Alle 4 Arten
schneiden und Tetramorium kabulistanicum. weisen Unterschiede in der Länge der Pro-
podealdomen auf: Tetramorium striativentre
Tetramorium argentirubrum besitzt 2-4,
(kleine Höcker) < Tetramorium argentiru-
im Mittel 2,8 (n = 12 Paratypen) subcepha-
brum < Tetramorium kabulistanicum < Tetra-
le abstehende Haare (die anterioren J-förmi-
morium schneideri. Tetramorium argentiru-
gen Haare der Psammophore bleiben unbe-
brum ist größer (Länge 3,0-3,2 mm) und ro-
rücksichtigt). Tetramorium kabulistanicum
buster als die anderen Arten (Länge Terra-
weist 4-5, im Mittel 4,7 (n = 3 Paratypen)
morium kabulistanicum 2,8-3,0 mm).
subcephale Haare auf. Im Gegensatz zu die-
sen beiden Arten sind die 6 subcephalen WHEELER & MANN (1916) geben Tetra-
Haare des untersuchten Tetramorium schnei- morium striativentre für Jordanien und das
323
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
dis, die sich durch Steppencharakter aus-
zeichnen (Abb. 4). Beide Standorte gehören
der irano-turanischen Region an.
Lepisiota (Baroniurbania)
Lepisiota (Baroniurbania) PAGLIANO &
SCARAMOZZINO 1989 n. Stat.
Acant/iolepis (Acantholepis) MAYR 1861;
SANTSCHI(1926)
PAGLIANO & SCARAMOZZINO (1989) er-
kannten, dass Acant/iolepis MAYR 1861 ein
jüngeres Homonym von Acantholepis KROE-
YER 1846 ist und führten für die Ameisen-
gattung den Ersatznamen Baroniurbania ein.
Dies war aber nicht notwendig, da ein jün-
geres, subgenerisches Synonym von Acan-
tholepis MAYR existierte. BOLTON (1994) er-
setzte daher Acant/iolepis MAYR durch Lepi-
siota SANTSCHI 1926. SANTSCHI (1926)
Abb. 5: REM-Aufnahme von Lepisiota südliche Syrien, KUGLER (1988) für Israel unterteilte die Gattung in den Subgenus
gracilicornis, Arbeiterin (Wadi Titin, a n D i e s e sü dlevantinische Verbreitung der Acantholepis s. str. MAYR (Typus-Art Hypo-
nordöstlich Aqaba, Jordanien); Maßstab: _ clinea frauenfeldi MAYR 1855) und den Sub-
100 um letramonum smanventre-Oruppe ist deutlich
genus Lepisiota (Typus-Art Plagiolepis roth-
vom vermutlichen Verbreitungszentrum
neyi FOREL 1884). Infolge der Homonymie
westlich Zentralasiens (Transkaspien, Tur-
ist aber Acant/iolepis s. str. MAYR als subge-
kestan, Afghanistan: EMERY 1909, KARAWA-
nerischer Name nicht mehr anwendbar. Da
JEW 1910, 1912, COLLINGWOOD 1960,1961,
sich sowohl Acantholepis s. str. MAYR, als
PlSARSKl 1967, 1969), aus dem 4 Formen be-
auch Baroniurbania PAGLIANO & SCARA-
schrieben wurden (Tetramorium striativentre,
MOZZINO auf den selben Typus beziehen,
Tetramorium schneiden, Tetramorium striati-
muss Baroniurbania als neuer Name für die
verirre schneiden var. longispina, Tetramorium
Untergattung gelten.
kabulistanicum), getrennt. Aufgrund dieser
Isolierung der nahöstlichen Tiere vom Ver- Die Differenzierung der Untergattungen
breitungszentrum und dem Fehlen von Te- Lepisiota s. str. und Baroniurbania erscheint
tramorium striativentre im jordanischen Ma- aufgrund des Formenreichtums und der
terial ist anzunehmen, dass es sich bei den deutlichen Morphologie gerechtfertigt. Ba-
Angaben von WHEELER & MANN (1916) roniurbania zeichnet sich durch einen grazi-
und KUGLER (1988) ebenfalls um Tetramo- len Körperbau mit langen Extremitäten aus.
rium argentirubrum handelt. Der Mesothorax ist verlängert und in der
Ökologie: Die Nester von Terramonum Mitte halsförmig verengt, die propodealen
argentirubrum befinden sich an beiden jorda- Stigmen sind durch spitze Auswüchse über-
nischen Standorten (Shaumari, Wadi Butm dacht (Abb. 5). Die eigenwillige Morpholo-
bei Qasr Amra, Abb. 1) im Bereich von Wa- gie muss im Zusammenhang mit ihrer Le-
bensweise als rasch laufende, tagaktive Be-
Tab. 1 : Mittelwerte, Standardabweichungen und Extremwerte von Kopf breite (KB),
wohner semiarider bis arider Gebiete be-
Scapuslänge (SL) und Hinterfemurlänge (FL) in um von Lepisiota gracilicornis (Wadi Titin,
8 km südlich Aqaba, ßaqir, Wadi Araba 36 km nördlich Aqaba) und Lepisiota opaciventris trachtet werden (siehe Diskussion).
(Wadi Titin, Petra, Wadi Hisman) im südlichen Jordanien bei n Arbeiterinnen aus m
Proben, sowie Signifikanzniveaus bei zweiseitigen t-Tests mit gleichen Varianzen Differentialdiagnose von Lepisiota graci-
(F-Tests p > 0,3). Ucomis (FOREL 1892) und Lepisiota opacwen-
tns (FINZI 1936):
Lepisiota gracilicornis Lepisiota opaciventris
n = 28 aus rt1 = 8 n = 8 aus m = 3 P Lepisiota opariventris besitzt im Vergleich
KB 619 ±33 [ 545; 667] L 550 ± 36 [ 498; 592]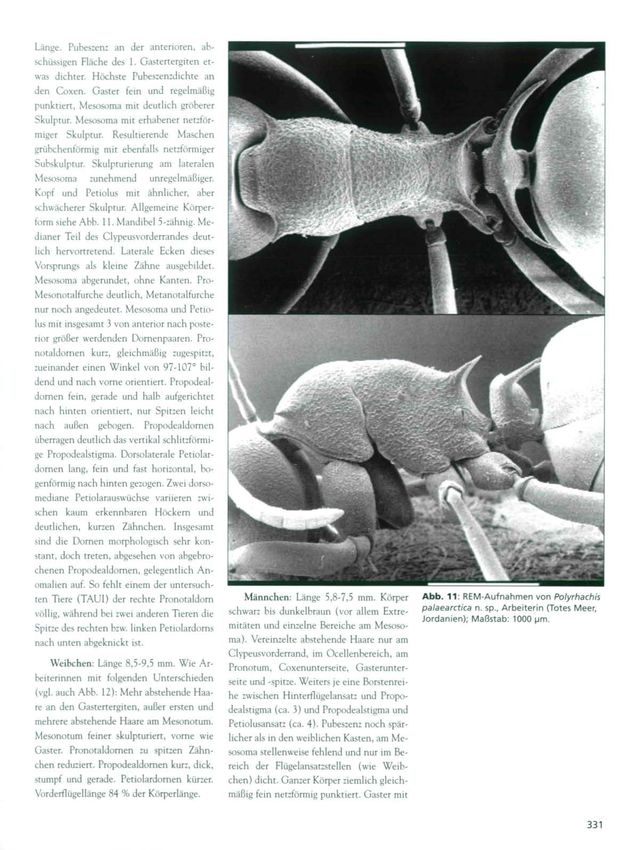
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
(Tab. 1, Abb. 6). Das steht im Widerspruch
0.60 T
zur Bestimmungstabelle von FlNZl (1936).
Er scheint aber Lepisiota graalicomis selbst
nicht gesehen zu haben, da er in Bezug auf
diese Art lediglich die Beschreibung von L graaTtcomis
FOREL (1892a) übersetzt und die Verbrei- 0.55-
tungsangaben von FOREL (1892a) und KA-
RAWAIEW (1911) übernommen hat. Lepisiota
opadventris weist einen sehr schmalen Kopf
auf(FlNZi 1936, Tab. 1). Im südjordanischen
Material lassen sich die beiden sympatri- O.SO-
schen, z. T. syntopen Arten morphome- m
trisch überlappungsfrei trennen. Zur Unter-
scheidung genügt ein Index (KB/SL oder
KB/FL), die Verwendung beider Indices er-
laubt aber eine sicherere Bestimmung. Als 0.45
Trennfunktion wurde eine Normale auf die
gemeinsame Regressionsgerade der beiden L opadventris
Cluster gewählt (Abb. 6). Daraus ergibt
sich:
0.40
KB > SLFL1,219/(SL+FL1,345) => 0.45 0,50 0,55 0.60 0,65
Lepisiota gracilicomis
KB/SL
KB < SLFL1,219/(SL+FL1,345) =>
Lepisiota opaciventris offen fouragierende Tiere, zum anderen sind Abb.6: Streudiagramm der Verhältnisse
sie optisch attraktiv (meist dichtes, goldenes Kopfbreite/Scapuslänge (KB/SL) und
Beide Arten haben ein sehr schwach Kopfbreite/Hinterfemurlänge (KB/FL) der
skulpuriertes, aber nicht völlig glattes Inte- Pubeszenzkleid an der Gaster; oft ein mit Arbeiterinnen von Lepisiota gracilianis und
gument. Lepisiota gracilicomis hat eine wa- dem dunklen Mesosoma kontrastierender Lepisiota graciaventris, einschließlich der
roter Kopf)- Diese Gruppe zeichnet sich Trennfunktion, aus dem südlichen
benförmige Oberflächenskulptur, dazwi-
Jordanien. Datengrundlage siehe Tab. 1.
schen ist das Integument fast glatt (Abb. 7). allerdings auch durch hohe morphologische
Die Waben werden am Gaster sehr flach, Variabilität, abgesehen von dem für Campo-
transversal gestaucht. Insgesamt erreicht Le- notus typischen Größenpolymorphismus,
pisiota gracilicomis bei schwarzen Tieren ei- aus. Gerade die differentialdiagnostisch ver-
nen leichten blaumetallischen Schimmer. wendeten Merkmale wie Färbung, Pube-
Lepisiota opaciventris besitzt ebenfalls solche szenz und Propodeumform weisen eine be-
Waben, hat aber dazwischen noch eine fei- achtliche Variationsbreite auf, deren taxo-
nere Subskulptur. Diese ist am Gaster quer, nomische Bedeutung noch nicht richtig ver-
am Kopf längs zur Körperachse orientiert. standen wird. Die leichte, mit freiem Auge
Durch diese Subskulptur erhält Lepisiota mögliche Ansprechbarkeit als „Camponotus
opaciventris einen seidigen Schimmer. Die sericeus" und die verwirrende Variabilität
Oberflächenskulpturen beider Arten sind mögen Gründe sein, dass bisher keine einge-
prinzipiell im Stereomikroskop erkennbar, henden Studien zu dieser Gruppe gemacht
im Zweifelsfall aber eine Präparation und
wurden. Das bisherige Bild, wonach eine
Begutachtung im Durchlichtmikroskop not-
einzige, nicht kosmopolitische, Ameisenart
wendig.
eine so weite Verbreitung hat, entspricht
wohl kaum der Realität. Vielmehr dürfte
Camponotus ser/ceus-Gruppe sich die Camponotus sericeus-Gruppe gerade
in Radiation befinden (siehe Diskussion).
Unter dem Sammelnamen „Camponotus
sericeus" verbirgt sich eine über weite Teile Eine momentane Radiation stellt die
Afrikas, der arabischen Halbinsel und In- überwiegend auf das Morphospezies-Kon-
diens verbreitete Gruppe, die sich durch be- zept basierende Taxonomie vor Probleme.
sondere Auffälligkeit auszeichnet. Zum ei- Vorliegende Arbeit versteht sich nur als
nen handelt es sich um große, tagaktive und kleiner Beitrag zur Camponotus sericeus-
325
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
FOREL (1892b, 1908, 1913),
WROUCHTON(1892)
Camponotus sericeus FABRICIUS; BlNGHAM
(1896,1903)
Camponotus sericeus opaciventris MAYR; PIS-
ARSKI (1969), RADCHENKO (1996)
Untersuchtes Material: Indien: 7 Arbeiterinnen-
Syntypen! im NHMW „Calcutta, Rothney,
Smith; C. opaciventris det. G. Mayr" bzw. „Cal-
cutta, Coll. G. Mayr; C. opaciventris det. G.
Mayr"; Afghanistan: Nengrahar, Jalalabad, leg.
Povolny l.-lll. 1965, Camponotus sericeus opaci-
ventris det. B. Pisarski, 1 Arbeiterin im MMBC.
Die von MAYR (1878) aus Indien be-
schriebene Camponotus opaciventris wurde
von FOREL (1892b, 1908, 1913) lediglich als
Variation von Camponotus sericeus betrach-
tet. WROUGHTON (1892) verwendet sowohl
die Bezeichnung Camponotus opaciventris als
auch Camponotus sericeus var: opaciventris.
BlNGHAM (1896, 1903) synonymisierte die
Form mit Camponotus sericeus. Erst PlSARSKI
(1969) gibt sie wieder als Unterart Campo-
notus sericeus opaciventris für Afghanistan an.
Dieser Status wurde bis heute (BOLTON 1995,
RADCHENKO 1996) beibehalten. Allerdings
gibt FOREL (1892b, 1908, 1913) die Variatio-
nen sericeus und opaciventris mehrfach sym-
patrisch an. Sympatrische Subspezies werden
aus heutiger Sicht ohne fundierte Begrün-
dung abgelehnt. Die Form opaciventris kann
daher nur als Synonym von Camponotus seri-
ceus oder als bona species betrachtet werden.
MAYR (1878) gibt zwei differentialdiag-
nostische Merkmale im Vergleich zu Campo-
notus sericeus an: die Gasterpubeszenz und
die Form des Propodeums. Die Bedeutung
Abb.7 REM-Aufnahmen der Kopfober- Gruppe, der auf das Problem hinweisen soll. des Fehlens oder Vorhandenseins der gold-
flächen der von Lepisiota gracilicornis Die hier aufgezeigte Dreiteilung in eine afri- schimmernden Gasterpubeszenz wurde in
(oben) und Lepisiota opaciventris (unten),
Arbeiterinnen (Wadi Titin, Jordanien); kanische „Camponotus serkeus", eine arabi- der Vergangenheit sicher überbewertet. Die-
Maßstab: 10 um. Der linke, untere Teil der sche Camponotus husseini n. sp. und eine in- ses „Pubeszenzkleid" scheinen Ameisen die-
Aufnahmen wird jeweils vom vorderen dische „Camponotus opaciventris" ist sicher ser Gruppe sehr leicht zu verlieren, so dass
Ocellus eingenommen.
nur ein erster grober Schritt. In diesem Sinn neben genetischer Variation auch Abnüt-
ist auch die Führung beider Formen unter zungen (auch als Artefakt!) die Verwendung
„ " zu verstehen. des Merkmals problematisch macht. MAYR
charakterisiert die Form des Propodeums wie
Camponotus opaciventris folgt: „Die Basalfläche des Metanotum ist
MAYR n. stat. von der Meso-Metanotalnaht bis zum hinte-
Camponotus opaciventris MAYR 1878 ren Rande flach, bei Camp, sericeus jedoch
ist sie vome in der Längsrichtung sehr deut-
Camponotus opaciventris MAYR; lich gewölbt, auch ist bei der neuen Art der
WROUGHTON(1892) hintere Theil derselben Basalfläche nicht,
Camponotus sericeus var. opaciventris MAYR; wie bei Camp, sericeus, etwas herabgebogen."
326© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
BlNGHAM (1903) hat diese Beschreibung
nicht richtig verstanden. Er schreibt in Be-
zug auf das Propodeum und ab Begründung
für seine Synonymisierung lediglich: „The
excavation of the posterior face of the meta-
notum varies individuals from the same
nest." Während also Mayr nur die dorsale
Form des Propodeums beschreibt, bezieht
sich Bingham nur auf die posteriore, ab-
schüssige Fläche. FOREL (1892b) berücksich-
tigt bei setner Differentialdiagnose der Vari-
ationen sericeus und opaciventris nur mehr die
Pubeszenz. Die Form des Metanotums vari-
iert seiner Ansicht nach graduell in Afrika
und Indien. Bemerkenswert ist allerdings
dass im Gegensatz zu FOREL (1892b) sowohl
WROUGHTON (1892) als auch BlNGHAM
(1903) für Indien nur eine Form von Cam-
ponotus sericeus angeben. BINGHAM (1903)
gibt bei der Beschreibung „seiner" Campono-
tus sericeus kein variables Propodeum an,
sondern beschreibt es folgendermaßen:
„Thorax ... emarginate at the meso-metano-
tal suture, the basal face of the metanotum
horizontal, flat, the sides margined, ...".
Ebenso beschreiben NEGI et al. (1930) das
Propodeum ihres indischen „Camponotus se-
riceus" als horizontal und flach. Diese Be
Schreibungen entsprechen genau den Typen
von Camponotus opacit/entris im NHMW,
wobei hinzuzufügen ist, dass die dorsale Flä-
che des Propodeums leicht schräg nach ante-
rior gerichtet ist. MUKERJEE (1930) gibt zwe;
Formen von „C. sericeus" aus Calcutta an.
die sich in Farbe, Skulptur und Pubeszen:,
aber nicht in Bezug auf das dorsale Propo-
deum unterscheiden. Es muss daher ange-
nommen werden, dass es sich bei der Mehr-
zahl der indischen Camponotus cf. sericeus-
Belege um Camponotus opaciventris handelt. BUSCHINGER & MASCHWITZ (1984) erwäh- Abb. 8: REM-Aufnahmen von Camponotus
Es sei aber eingeräumt, dass einzelne Indivi- husseini n. sp., Arbeiterin (Wadi Araba,
nen ähnliches bei „Camponotus sericeus".
Jordanien); Maßstab: 1000 [im.
duen aufgrund der Propodeumform schwierig Diese Beobachtung wurde in Sri Lanka ge-
zuzuordnen sind. Dies trifft ins besonders auf
macht, die Artbestimmung ist unsicher
Belege von Sri Lanka zu. Der afghanische
(MASCHWITZ in litt. 1999). MUKERJEE
Beleg (PISARSKI 1969, MMBC) ist zweifels-
(1930) gibt allerdings aus Calcutta mit dem
frei als Camponotus opaciventris zu benennen.
Boden ebene Nesteingänge an. Schornst-
Bei indischen Camponotus cf. sericeus einförmige Nesteingänge werden bei Cam-
wurden schornsteinförmige Nesteingänge ponotus cf. sericeus aus der Äthiopis oder der
beschrieben. WROUGHTON (1892) schreibt arabischen Halbinsel nicht erwähnt.
unter Camponotus sericeus var. opaciventris:
„it is generally furnished with a built-up tu- Aus den genannten Gründen erscheint
bular entrance, rising less than an inch ab- bei gegenwärtigem Kenntnisstand die Be-
ove the surface; this porch is built of minu- trachtung von Camponotus opaciventris als
te pieces of grass, worked up with mud, ...". bona species am sinnvollsten.
327© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
nem weitgehend rotem Mesosoma auf. Füh-
ler bei dunkelköpfigen Tieren dunkelbraun,
bei rotköpfigen Tieren hellbraun. Distaler
Teil des Scapus angedunkelt. Beine wie Me-
sosoma gefärbt, distal, zumindest Tarsenspit-
ze, heller werdend. Gaster infolge Pubeszenz
golden. Körper ± regelmäßig abstehend be-
haart. Extremitäten hauptsächlich nieder-
liegend behaart, nur an Beinunterseite ab-
stehende Haare. Gesamter Clypeusvorder-
rand mit einer Reihe (ca. 8) auffälliger, lan-
ger distal leicht nach unten gekrümmten
und zugespitzten Borsten (ca. so lang wie
Clypeus). Diese werden offenbar leicht ab-
genutzt und sind dann z. T. wesentlich kür-
zer, stumpf und ohne distale Krümmung en-
dend. Am Hinterrand des Mentums 2 lange
J-förmige Borsten (Psammophore). Einen
weiteren Teil der Psammophore stellen zahl-
reiche J-förmige nach unten und innen ge-
richtete Borsten am Mandibelaußenrand
Abb. 9: Locus typicus von Camponotus Camponotus husseini n. sp. (Abb. 8)
husseini n. sp. (Sibkh et Täba, Wadi Araba, dar. Auf der Kopfunterseite in der Regel ei-
Jordanien). Locus typicus: Wadi Araba (Abb. 1, 9), nige abstehende Haare von unterschied-
Jordanien licher Länge (Tab. 2). Pubeszenz von ante-
Typus-Material: Holotypus-Arbeiterin, Jorda- rior nach posterior zunehmend. Abstand
nien: Wadi Araba 36 km nördlich Aqaba, zwischen den Pubeszenzhärchen am Kopf oft
26.10.1996 CO. Dietrich; - Paratypen: Jorda- geringer als deren Länge, dorsales Gasterin-
nien: 11 Arbeiterinnen Wadi Araba 36 km nörd- tegument nahezu vollständig von einem Pu-
lich Aqaba, 26.10.1996 CO. Dietrich. Der Holo- beszenzkleid bedeckt. Der Bereich des Cly-
typus und 5 Paratypen befinden sich im NHMW, peus bildet eine Ausnahme und liegt in Be-
je 3 in TAUI und MMBC
zug auf Länge und Dichte der Härchen zwi-
Untersuchtes Material: Jordanien: Wadi Araba schen Mesosoma und Gaster. Das gastrale
36 km nördlich Aqaba, 26.10.1996 CO. Die- Pubeszenzkleid unterliegt offenbar ebenfalls
trich; Südende Totes Meer, 19.41998 CO. Die-
Abnützungen und kann daher stellenweise
trich; Ägypten: Gebel Elba, 1.2.1933 H. Priesner;
aufgelichtet sein. Gesamtes Integument fein
Jemen: Hamdan, Wadi Dhar, 20.9.1996 H. Burt-
punktiert, am Mesosoma lateral in eine grö-
scher; Hadramaut, Tarim, 26.9.1996 H. Burt-
scher; El Kod, 10.12.1988 W. Wranik; Sana'a, bere netzförmige Skulptur übergehend. All-
3.1993 CA. Collingwood; Saudi Arabien: Riyad, gemeine Körperform siehe Abb. 8. Die Ar-
10.1.1980 A.S. Talhouk; Riyad, 3.1983 C.A.Col- beiterinnen zeigen den für Camponotus typi-
lingwood. schen Größenpolymorphismus. Mandibel 5-
Etymologie: Zu Ehren König Husseins zähnig. Clypeusvorderrand nach der oben
von Jordanien, der im Zeitraum der Bearbei- genannten Borstenreihe median rechtwin-
kelig abgestutzt. Auf dieser schmalen Fläche
tung verstarb.
befindet sich noch eine Reihe sehr kleiner,
Diagnose: Arbeiterinnen: Länge abstehender Börstchen. Abgestutzte Fläche
5,4-9,8 mm. Kopf dunkelrot bis schwärzlich, bei sehr großen Tieren als breite Einkerbung
restlicher Körper mit dunkelbraunem bis am Clypeusvorderrand ausgebildet. Propo-
schwarzem Integument. Vorallem die südle- deum abgesetzt, dorsal parallelseitig, die Sei-
vantinischen Tiere zeigen einen farblich tenränder scharfkantig, posteriore Ecken als
deutlichen Kontrast zwischen Kopf und Me- stumpfe Zähne oder Lappen ausgezogen.
sosoma, während bei den jemenitischen Tie- Propodeum bei lateraler Ansicht deutlich
ren häufig kaum ein farblicher Unterschied konvex, posterior stärker abfallend als ante-
erkennbar ist. Exemplare vom ägyptischen rior. Petiolus dick, aber nicht knotenförmig.
Gebel Elba weisen einen roten Kopf mit ei-
328© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
Tab. 2: Mittel- und Extremwerte der Anzahl subcephaler Borsten und Probenherkunft der Arbeiterinnen von
Camponotus husseini n. sp. und „Camponotus sericeus".
Camponotus husseini n. sp. „Camponotus sericeus"
Jordanien, Totes Meer, CCD, n = 4 8,3 [5; 10] Gambia, CCC, n = 1 0,0
Jordanien, Wadi Araba, CCD, n = 21 3,3 [0; 9] Mali, Kita, NHMW, n = 8 0,0 [0; 0]
Jemen, Tarim, CKW, n = 6 4,5 (3; 5] Tschad, Mt.Kelinguen, NHMW, n = 11 0,0 [0; 0]
Jemen, Wadi Dhar, CKW, n = 6 5,0 [3; 8] Tschadgebiet, 3 Orte, NHMW, n = 4 0,0 [0; 0]
Jemen, El Kod, CWW, n = 3 3,0 [2; 5] Ägypten, Siwa, NHMW, n = 7 1,4 [0;2]
Jemen, Sana'a, CCC, n = 10 4,5 M;7] Burundi, Bujumbura, LMO, n = 3 0,0 [0; 0]
Saudi Arabien, Riyad, CCC, n = 2 5,0 [2;8] Uganda, Karamoja, LMO, n = 9 0,1 [0; 1]
Ägypten, Gebel Elba, NHMW, n = 3 6,0 [5; 7] Tanzania, Manyara, Meru, LMO, n = 4 0,8 [0; 1)
Gruppenwerte, m = 8 5,0 [3,0; 8,3] Gruppenwerte, m = 8 0,3 [0,0; 1,4]
Differentialdiagnose: Das herangezoge- Camponotus husseini und „Camponotus
ne Vergleichsmaterial von dem aus West- sericeus" sind durch diese Funktion nicht
afrika (Senegal) beschriebenen Camponotus überlappungsfrei, aber deutlich getrennt
sericeus stammt hauptsächlich aus östlicher (Fisher-Yates-Test p < 0,001; n = 90). Beide
gelegenen Populationen (Mali, Ägpyten, Merkmale - subcephale Beborstung und Pe-
Tschad, Burundi, ...), also zwischen Locus tiolusform - können bei Einzeltieren zu De-
typicus von Camponotus sericeus und der bis- terminationsschwierigkeiten führen, es sind
her bekannten Verbreitung von Camponotus daher Kleinserien anzuraten. Vor allem auch
husseini. Die Bezeichnung dieser Tiere als in Hinblick darauf, dass die Taxonomie der
„Camponotus sericeus" ist daher provisorisch Camponotus sericeus-Gruppe noch unklar
zu verstehen. Ein Exemplar stammt aus ist. Beispielsweise besitzen die Tiere der Pro-
Gambia, d. h. sehr nahe dem Typenort von be Tschad, Mt. Kelinguen (Kreuze in Abb.
Camponotus sericeus. 10) keine subcephale Borsten, fällt aber von
der Petiolusform eindeutig in den Campono-
Beide, Camponotus husseini und „Campo-
tus husseini-Cluster.
notus sericeus" unterscheiden sich durch die
Form des Propodeums von Camponotus opa- Ökologie: Alle jordanischen Standorte
civentris (siehe dort). Vorallem Camponotus von Camponotus husseini befinden sich in
husseini ist durch die Form des Propodeums der sudanischen Region des Wadi Araba
deutlich von indischen Populationen ge- und des südlichen Toten Meeres, wo er syn-
trennt. Camponotus husseini unterscheidet top mit Polyrhachis palaearctica auftreten
sich von den beiden anderen Formen durch kann (Abb. 1). An allen Standorten befin-
mehrere unterschiedlich lange mediane und den sich wenigstens vereinzelte Gehölze
laterale subcephale Borsten (Tab. 2, Abb. 8). (Abb. 9), die vermutlich für trophobionti-
„Camponotus sericeus" und „Camponotus sche Beziehungen benötigt werden. Derarti-
opaciventris" haben keine oder nur aus- ge Beziehungen wurden in der Camponotus
nahmsweise seitlich 1-2 kleine subcephale sericeus-Gruppe mehrfach angegeben
Borsten. Grundsätzlich verändert sich die (WROUGHTON 1892, MUKERJEE 1930, COL-
Petiolusform bei Arten der Camponotus LINGWOOD 1985).
sericeus-Gruppe mit zunehmender Indivi-
duengröße von nodiform zu squamiform
(Abb. 10). Der Petiolus von Camponotus
husseini ist aber signifikant schmäler als bei
„Camponotus sericeus" (Tab. 3). Als Trenn-
funktion wurde die Symmetrieachse zwi-
schen den Regressionsgeraden der Cluster Tab. 3: Mittelwerte, Standardabweichungen und Extremwerte von Kopfbreite (KB) in pm
und Petiolarindex (Pl) von Camponotus husseini n. sp. und „Camponotus sericeus"
Camponotus husseini und „Camponotus seri-
(Datengrundlage siehe Tab. 2) bei n Arbeiterinnen aus m Proben, sowie
ceus" gewählt (Abb. 10). Daraus ergibt sich: Signifikanzniveaus bei zweiseitigen t-Tests mit gleichen Varianzen (F-Test p > 0,6).
KB > (PI-O,7137)/0,000246 => „Cam- Camponotus husseini n. sp. „Camponotus sericeus"
ponotus sericeus" n = 57 aus m = 9 n = 36 aus m = 11 P
KB < (P1-O,7137)/O,OOO246 => Campo- KB 1991 ±485 [1491; 3321] 2021 ± 448 [1510,3057] n. s.
notus husseini Pl 1,307 ±0,136 [1,105; 1,667] 1,110 ±0,131 [0,962; 1,480] < 0,001
329© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
chis dives-Gruppe ist allerdings derzeit un-
1.7 klar. Gleichzeitig ist „Polyrhachis simplex" bis
in die jüngste Zeit ein gebräuchlicher Na-
1.6 C. husseini n. sp. me: BODENHEIMER (1929, 1935), MENOZZI
(1929, 1933), MUKERJEE (1930), MUKERJI
1.5 (1932), FINZI (1936), BYTINSKI-SALZ
(1953), WHEELER 6k WHEELER (1953,1974),
1.4 COLLINGWOOD (1960), HUNG (1967), Pl-
SARSKI (1967, 1969), OFER (1970), Aziz 6k
K 1.3 AL-ALI (1977), OFER et al. (1978), HEFETZ
& ORION (1982), COLLINGWOOD (1985),
1.2 DEGEN et al (1986), KUGLER (1988), DEGEN
6k GERSANI (1989), TIWARI (1999). Aus die-
1.1 sen Gründen sei hier der Einfachheit halber
"C. sericeus"
von „Polyrhachis simplex" die Rede und des-
1 sen Abgrenzung von einigen ähnlichen
PotyrJmchis-Arten im Folgenden behandelt.
0,9 •+• -+• -+• •+•
1400 1600 1800 2000 2200 2400 2600 2800 3000 3200 3400 Polyrhachis palaearctica n. sp.
KB [Mm] (Abb. 11, 12)
Locus typicus: Totes Meer (Abb. 1,13),
A b b . 10: Streudiagramm der Maße Kopfbreite (KB) und Petiolarindex (PI) der
Arbeiterinnen von Camponotus husseini n. sp. (voller Kreis) und „Camponotus sericeus" Jordanien
(leere Symbole), einschließlich der Trennfunktion. Datengrundlage siehe Tab. 2, 3. Die
Typus-Material: Holotypus-Arbeiterin, Jorda-
Kreuze markieren die Probe „Tschad, Mt. Kelinguen", welche die subcephale Beborstung
nien: Totes Meer 5,4 km nördlich Mündung Wa-
von „Camponotus sericeus" und den Petiolus von Camponotus husseini aufweist. Die
Exemplare mit größter Nähe zum Locus typicus von Camponotus sericeus (Senegal) sind di Main, 29.10.1996 CO. Dietrich; - Paratypen:
mit einem Quadrat markiert (Gambia, Mali). Jordanien: 11 Arbeiterinnen Totes Meer 5,4 km
nördlich Mündung Wadi Main, 29.10.1996 CO.
Polyrhachis (Myrmhopla) Dietrich. Der Holotypus und 5 Paratypen befin-
den sich im NHMW, je 3 in TAU1 und MMBC.
Eine Revision der orientalischen Polyr-
/lac/iis-Arten fehlt bislang, obwohl es seit Untersuchtes Material: Jordanien: Totes Meer
MAYR (1862) nicht an Ordnungsversuchen 5,4 km nördlich Mündung Wadi Main,
mangelt. Die aus jüngerer Zeit stammende 29.10.1996 CO. Dietrich; Hammamat Main,
1.11.1996 CO. Dietrich; Südende Totes Meer,
Revision auf subgenerischem Niveau von
19.4.1998 CO. Dietrich; 1 Arbeiterin 30 km
HUNG (1967) brachte kaum brauchbare Er-
nördlich Tafila, 2.5.1996 M. Haiada, LMO; Is-
kenntnisse. Erst DOROW (1996) schaffte rael: 2 Weibchen En Gedi, 20.5.1970 J. Kugler; 3
durch seine Revision auf Artgruppenniveau Arbeiterinnen, 1 Männchen W. Avugot,
eine fundierte Basis. Die für den westpalä- 17.8.1966 J. Kugler, TAUI; Jemen: 1 Männchen
arktischen Raum hauptsächlich relevante Aden, 29.10.1988 W. Wranik.
Polyrhachis dives-Gruppe muss aber, wie die Etymologie: Die paläotropische Gat-
Untersuchung des Materials aus dem Nahen tung Polyrhachis dringt nur ausnahmsweise
Osten und des NHMW zeigte, einige Neue- in die Paläarktis vor. Soweit derzeit bekannt
rungen erfahren. hat die neue Art als einzige ihren Verbrei-
BOLTON (1974) synonymisiert „this well tungsschwerpunkt in der Paläarktis.
known name" Polyrhachis simplex MAYR Diagnose: Arbeiterinnen: Länge
1862 (ohne Typenmaterial gesehen zu ha- 5,7-8,0 mm. Ganzer Körper schwarz. Abste-
ben) mit den in Vergessenheit geratenen hende Haare nur an Gaster- und Beinunter-
Polyrhachis lacteipennis SMITH 1858. COL- seiten, sowie je ein medianes Borstenpaar
LlNGWOOD & AGOSTI (1996) geben für die am Hinterkopf, zwischen den Stirnleisten
Arabische Halbinsel Polyrhachis lacteipennis und am Clypeusvorderrand. Auf den ersten
an („P. simplex MAYR 1862 which is probably 3 Gastertergiten meist 0-1, am 3. Tergiten
a synonym"), ohne Boltons Synonymisie- bis zu 3 abstehende Haare. Ganzer Körper
rung zu berücksichtigen. Die taxonomische mit schütterer Pubeszenz. Abstand zwischen
und zoogeographische Situation der Polyrha- Härchen im allgemeinen größer als ihre
330© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
Länge. Pubeszenz an der anterioren, ab-
schüssigen Räche des 1. Gastertergiten et-
was dichter. Höchste Pubeszenzdichte an
den Coxen. Gaster fein und regelmäßig
punktiert, Mesosoma mit deutlich gröberer
Skulptur. Mesosoma mit erhabener netzför-
miger Skulptur. Resultierende Maschen
grübchenförmig mit ebenfalls netzförmiger
Subskulptur. Skulpturierung am lateralen
Mesosoma zunehmend unregelmäßiger.
Kopf und Petiolus mit ähnlicher, aber
schwächerer Skulptur. Allgemeine Körper-
form siehe Abb. 11. Mandibel 5-zähnig. Me-
dianer Teil des Clypeusvorderrandes deut-
lich hervortretend. Laterale Ecken diese-
Vorsprungs als kleine Zähne ausgebildet.
Mesosoma abgerundet, ohne Kanten. Pro-
Mesonotalfurche deutlich, Metanotalfurche
nur noch angedeutet. Mesosoma und Petio-
lus mit insgesamt 3 von anterior nach poste-
rior größer werdenden Dornenpaaren. Pro-
notaldornen kurz, gleichmäßig zugespitzt,
zueinander einen Winkel von 97-107° bil-
dend und nach vorne orientiert. Propodeal-
dornen fein, gerade und halb aufgerichtet
nach hinten orientiert, nur Spitzen leicht
nach außen gebogen. Propodealdornen
überragen deutlich das vertikal schlitzförmi-
ge Propodealstigma. Dorsolaterale Petiolar-
dornen lang, fein und fast horizontal, bo-
genförmig nach hinten gezogen. Zwei dorso-
mediane Petiolarauswüchse variieren zwi-
schen kaum erkennbaren Höckern und
deutlichen, kurzen Zähnchen. Insgesamt
sind die Dornen morphologisch sehr kon-
stant, doch treten, abgesehen von abgebro-
chenen Propodealdornen, gelegentlich An-
omalien auf. So fehlt einem der untersuch-
ten Tiere (TAUI) der rechte Pronotaldorn Mannchen: Länge 5,8-7,5 mm. Körper A b b . 11: REM-Aufnahmen von Polyrhachis
völlig, während bei zwei anderen Tieren die schwarz bis dunkelbraun (vor allem Extre- palaearctica n. sp., Arbeiterin (Totes Meer,
Spitze des rechten bzw. linken Petiolardorns Jordanien); Maßstab: 1000 |jm.
mitäten und einzelne Bereiche am Mesoso-
nach unten abgeknickt ist. ma). Vereinzelte abstehende Haare nur am
Clypeusvorderrand, im Ocellenbereich, am
Weibchen: Länge 8,5-9,5 mm. Wie Ar- Pronotum, Coxenunterseite, Gasterunter-
beiterinnen mit folgenden Unterschieden seite und -spitze. Weiters je eine Borstenrei-
(vgl. auch Abb. 12): Mehr abstehende Haa- he zwischen Hinterflügelansatz und Propo-
re an den Gastertergiten, außer ersten und dealstigma (ca. 3) und Propodealstigma und
mehrere abstehende Haare am Mesonotum. Petiolusansatz (ca. 4). Pubeszenz noch spär-
Mesonotum feiner skulpturiert, vorne wie licher als in den weiblichen Kasten, am Me-
Gaster. Pronotaldornen zu spitzen Zähn- sosoma stellenweise fehlend und nur im Be-
chen reduziert. Propodealdornen kurz, dick, reich der Flügelansatzstellen (wie Weib-
stumpf und gerade. Petiolardornen kürzer. chen) dicht. Ganzer Körper ziemlich gleich-
Vorderflügellänge 84 % der Körperlänge. mäßig fein netzförmig punktiert. Gaster mit
331© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
chis palaearctica) und indisch-pakistanischen
(„Polyrhachis simplex") Populationen.
Arbeiterinnen: Besonders deut-
lich tritt der Unterschied zwischen Polyrha-
chis palaearctica und JPolyrhachis simplex" in
der Form der Propodealdornen hervor. Bei
ersterer sind diese nur halb aufgerichtet und
überragen deutlich die Propodealstigmen.
Bei „Polyrhachis simplex" sind die Dornen
steil aufgerichtet bzw. aufgebogen, so dass
diese die Propodealstigmen nicht oder kaum
überragen. Diese stark aufgerichteten Pro-
podealdornen der indischen „Polyrhachis
simplex" („metanotal spines erect") sind in
der Abbildung von BlNGHAM (1903) vor al-
lem im Vergleich zur Polyrhachis hauxuieüi
(„metanotal spines suberect") erkennbar.
Ebenso schreibt HlNGSTON (1923) über die
Propodealdornen seiner hindustanischen
Abb. 12: REM-Aufnahme von Polyrhachis seichterer Skulptur und daher glänzender. Polyrhachis simplex: „are large and prominent
palaearctica n. sp., Weichen (En Gedi, Gröbste Netzskulptur am Propodeum und and their points are turned upward and er-
Israel); Maßstab: 1000 um. ect". Um diese Vertikalität der Propodeal-
Mesosomaseiten. Mandibel einfach gekerbt.
Clypeus ohne auffälligen Vorsprung. Kopf- dornen morphometrisch verwerten zu kön-
morphometrie siehe Differentialdiagnose. nen, wurde der Dornstigma-Index (DSI)
Pronotal- und Propodealdornen fehlen völ- verwendet. „Polyrhachis simplex" zeigt einen
lig. Petiolus nach oben deutlich divergie- DSI nahe Null, d. h. Dornenspitze überragt
rend, trapezförmig. Die oberen, lateralen wie oben erwähnt kaum das Stigma, wäh-
Ecken dieses Trapez (seitliche Petiolardor- rend Polyrhachis palaearctica einen 5-fach
nen) als deutliche, spitze Dömchen ausgezo- größeren DSI aufweist (Tab. 4). Weiters sind
gen. Vorderflügellänge 96 % der Körperlän- die Propodealdornen bei Polyrhachis palae-
ge. arctica deutlich weniger spreizend als bei
A b b . 13: Locus typicus von Polyrhachis „Polyrhachis simplex" (PDW, Tab. 4). Zur si-
palaearctica n.sp. (Kleinoase am Toten Differentialdiagnose; beruht auf dem cheren Trennung empfiehlt es sich, beide
Meer, nördlich AI Mara, Jordanien). Vergleich zwischen nahöstlichen (Polyrha- Merkmale in Kombination anzuwenden
(Abb. 14). Als Trennfunktion wurde eine
Normale auf die gemeinsame Regressionsge-
rade ( D S I ^ = DSI-218) der Cluster Poly-
rhachis palaearctica und „Polyrhachis simplex"
gewählt.
PDW < DSI-218 + 89,22 =>
Polyrhachis palaearctica
P D W > DSI-218+ 89,22 =>
polyrhachis simplex"
Polyrhachis spiniger und Polyrhachis grises-
cens unterscheiden sich von Polyrhachis pa-
laearctica durch apical stark auswärts gebo-
gene Propodealdornen, der Dornenmorpho-
metrie (Abb. 14), gröberer Skulptur am Me-
sosoma und, ebenso wie polyrhachis sim-
plex", mehrere abstehende Haare an den er-
sten 3 Gastertergiten (vor allem am 3. Ter-
giten mehr als 5 Haare).
332© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
Weibchen: Alle drei Dornenpaare sind Tab. 4: Mittelwerte, Standardabweichungen und Extremwerte von Pronotalwinkel (PDW)
in Grad und Dornstigma-Index (DSI) von Polyrhachis palaearctica n. sp. (Jordanien, Totes
bei Polyrhachis palaearctica kürzer als bei
Meer CCD, LMO; Israel, Avdat TAUI) und „Polyrhachis simplex" (Afghanistan, Dar-i-Nur
„Polyrhachis simplex". Der auffälligste Unter- MMBC; Pakistan, Balakot NHMW; Indien, Calcutta NHMW; Himalaya NHMB) bei n
schied besteht in den Pronotal- und Propo- Arbeiterinnen aus m Proben, sowie Signifikanzniveaus bei zweiseitigen t-Tests mit
dealdornen. Die Pronotaldornen sind bei gleichen Varianzen (F-Test p > 0,6; PDW) und ungleichen Varianzen (F-Test p< 0,1; DSI)
Polyrhachis palaearctica zu kleinen Zähnen Polyrhachis palaearctica n. sp. „Polyrhachis simplex"
reduziert (Abb. 11, OFER 1970), während sie n = 9 aus m = 4 n = 8 aus m = 4 P
bei „Polyrhachis simplex" deutlich ausgebildet PDW 102 ±4,1 i [97; 107] 121 ±4,8 [115; 128] < 0,001
und jenen der Arbeiterinnen wesentlich DSI 0,119 ± 0,0230 ! [0,089; 0,155] 0,024 ± 0,0428 [-0,020; 0,085]© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
denen Nesttypen, wobei immer vorgegebe- und führen dies unabhängig voneinander
ne Hohlräume in Bodennähe ausgebaut auf unterschiedliche jahreszeitliche Feuch-
werden. Arboricole Nester sind selten und tigkeitsbedingungen zurück. Aufgrund der
dann am Stamm in Bodennähe. MAXWELL- verschiedenen klimatischen Situation in Is-
LEFROY & HOWLETT (1909), DUTT (1912) rael und Hindustan erfolgen diese Wande-
und HlNGSTON (1923) berichten aus Indien rungen auch zu unterschiedlichen Zeiten. In
neben den bodennahen Nestern auch von, Israel dauern sie jeweils einen Monat von
für Weberameisen typischen, arboricolen April-Mai und zurück von November-De-
Nestern an Asten. Sie unterscheiden diese zember. In Hindustan verlassen die Ameisen
größeren und festeren arboricolen Nester ihre Bodennester zu Sommerbeginn und
deutlich von den arboricolen Trophobion- verbringen den Monsun in ihren arborico-
ten-"Ställen", wie sie auch aus Israel (OFER len Nestern. Im September verliert der
1970, DEGEN & GERSANI 1989) bekannt Monsun seine Kraft, und die Tiere kehren
sind. MUKERJI (1932) berichtet ebenfalls zurück.
von arboricolen Nestern bei Calcutta. Die
Nester der israelischen Weberameisen sind Schwärmverhalten: Das Auftreten der
ohne Kammern (OFER 1970). HlNGSTON Geschlechtstiere ist erstaunlich schlecht
(1923) beschreibt sowohl für die Bodennes- dokumentiert. Aus dem Nahen Osten (Si-
ter, als auch für die arboricolen Bauten zahl- nai) gibt es nur eine Meldung von Mitte Ju-
li (BODENHEIMER & THEODOR 1929, MENOZ-
reiche Nestkammern. Eine solche Kompar-
timentierung des Nestes ist bei Polyrhachis ZI 1929). Das alate Weibchen von En Gedi
selten (MUKERJI 1932). (Israel) wurde am 20.6., das Männchen von
W. Avugot (Israel) am 17.8. und das jeme-
Sehr unterschiedlich sind auch die Be- nitische Polyrhachis Jwlaearctica-Männchen
schreibungen des Nesteinganges, die auf (Aden) am 29.10. gesammelt. Für Indien
unterschiedliche Defensivstrategien schlie- wird August, September (DUTT 1912) und
ßen lassen. OFER (1970) beschreibt den Oktober (WROUGHTON 1892) angegeben.
Nesteingang der israelischen Weberameise Aus den nordindischen (WROUGHTON
lediglich als enge, runde Öffnung mit einem 1892) und afghanischen (COLLINGWOOD
Durchmesser von 2-3 mm, welche die 1960) Bergen gibt es Einzelhinweise auf En-
gleichzeitige Passage zweier Ameisen ermög- de Mai. Bemerkenswert ist das Verhältnis
licht. Befindet sich das Nest in einer Höhle, zwischen Vorderflügel- und Körperlänge.
wird der Eingang mit einem Vorhang aus Während bei „Polyrhachis simp!ex"-Königin-
Seidengespinst verschlossen. HlNGSTON nen die Flügel länger als der Körper sind, ist
es bei Polyrhachis palaearctica und den
(1923) beschreibt den Eingang bodennaher
Männchen von „Polyrhachis simplex" (89-92
Nester der indischen „Polyrhachis simplex"
%) umgekehrt. Dies lässt schließen, dass
als ein „few inches" hohes, vertikales Türm-
„Polyrhachis simplex" nach dem Hochzeits-
chen aus Seidengespinst. An der Spitze des
flug zu größeren Dispersionsleistungen fähig
Türmchens beobachtete er oft den Kopf ei-
ist.
nes Wächters: „Sometimes, like a Stopper in
the mouth of a bottle, its head is thrust firm-
Ökologie: Aus Jordanien wurde Polyrha-
ly into the gate, and in this efficient manner
chis palaearctica bisher nur in der sudani-
it closely bars the way." Vielleicht steht die-
schen Region des Toten Meeres gefunden
ses Verhalten im Zusammenhang mit der
(WHEELER & MANN 1916, Abb. 1). Polyrha-
Morphologie der Propodealdornen. Im
chis palaearctica benötigt einen wesentlich
Gegensatz zur Polyrhachis palaearctica besitzt
dichteren Vegetationsbestand als Campono-
die „Polyrhachis simplex" stark aufgerichtete
tus husseini, kann aber auch salzreiche Klei-
Dornen. Damit könnte sie sich im Gespinst
noasen nutzen (Abb. 13). Detailiertere An-
des engen Türmchens verankern, um nicht
gaben zur Ökologie befinden sich in OFER
von einem Gegner herausgezogen zu wer-
(1970), DEGEN & GERSANI (1989) und DE-
den.
GEN et al. (1989).
Sowohl HlNGSTON (1923) als auch ÖFER
(1970) berichten von jahreszeitlich beding- Polyrhachis spiniger MAYR n. stat.
ten Wanderungen (und somit Nestwechsel) Polyrhachis spiniger MAYR 1878
334Sie können auch lesen

















































