Untersuchungen zum Einfluss von Vav-Proteinen auf die B-Zell-Entwicklung
←
→
Transkription von Seiteninhalten
Wenn Ihr Browser die Seite nicht korrekt rendert, bitte, lesen Sie den Inhalt der Seite unten
Universitätsklinikum Ulm
Universitätsklinik für Hals-Nasen-Ohrenheilkunde, Kopf- und Halschirurgie
Ärztlicher Direktor: Prof. Dr. med. T. Hoffmann
Forschungslabor
Leitung: Prof. Dr. rer. nat. C. Brunner
Untersuchungen zum Einfluss
von Vav-Proteinen auf die
B-Zell-Entwicklung
Dissertation zur Erlangung des Doktorgrades der
Medizin der Medizinischen Fakultät der Universität Ulm
Sebastian Karl Kieser
Stuttgart
2017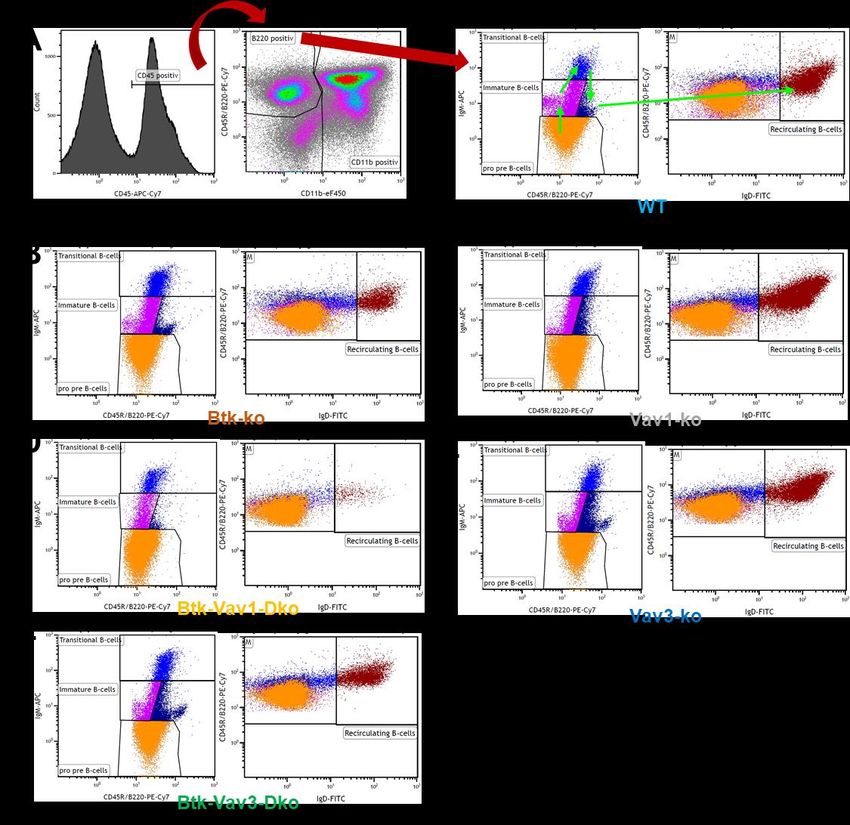
Amtierender Dekan: Prof. Dr. T. Wirth 1. Berichterstatter: apl. Prof. Dr. C. Brunner 2. Berichterstatter: apl. Prof. Dr. R. Schirmbeck Tag der Promotion: 13.12.2018
Für
meine Eltern
und
meinen Engel.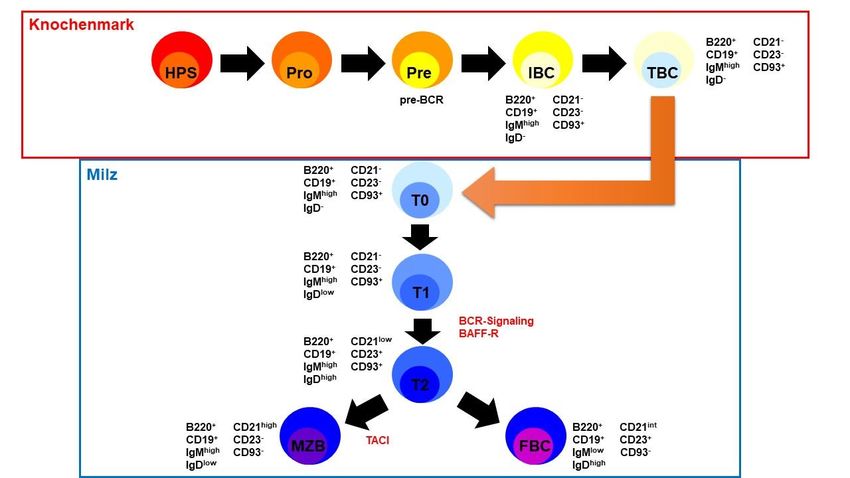
Ergebnisse dieser Doktorarbeit wurden in Form einer Posterpräsentation auf dem 47. jährlichen Treffen der deutschen Gesellschaft für Immunologie (47th Annual Meeting of the German Society for Immunology) vom 12.-15. September 2017 in Erlangen präsentiert: “Influence of Vav1 and Vav3 on B cell development and function”; Sebastian Kieser, Katja Fiedler, Simon Laban, Patrick J. Schuler, Klaus- Dieter Fischer, Thomas K. Hoffmann, Cornelia Brunner.
INHALTSVERZEICHNIS
ABKÜRZUNGSVERZEICHNIS ..................................................................................................... III
1 EINLEITUNG .......................................................................................................................... 1
1.1 B-ZELL-ENTWICKLUNG............................................................................................................. 1
1.1.1 ENTWICKLUNG IM KNOCHENMARK ................................................................................................... 1
1.1.2 PERIPHERE REIFUNG ....................................................................................................................... 2
1.2 B-ZELL-REZEPTOR (BCR)-NACHGESCHALTETE SIGNALWEGE .............................................................. 4
1.3 VAV-PROTEINE ...................................................................................................................... 8
1.3.1 VAV-ÜBERBLICK............................................................................................................................. 8
1.3.2 GEF-FUNKTION ............................................................................................................................. 9
1.3.3 INTERAKTIONSPARTNER DER VAV-PROTEINE ..................................................................................... 11
1.4 BRUTON’SCHE TYROSINKINASE (BTK) ........................................................................................ 12
1.5 ZIEL DER ARBEIT ................................................................................................................... 14
2 MATERIAL UND METHODEN ................................................................................................ 15
2.1 MATERIAL .......................................................................................................................... 15
2.1.1 MAUSLINIEN ............................................................................................................................... 15
2.1.2 CHEMIKALIEN UND REAGENZIEN ..................................................................................................... 16
2.1.3 PUFFER ...................................................................................................................................... 17
2.1.4 ANTIKÖRPER FÜR DIE DURCHFLUSSZYTOMETRIE ................................................................................ 18
2.1.5 PRIMER FÜR DIE POLYMERASE-KETTENREAKTION .............................................................................. 19
2.1.6 VERBRAUCHSMATERIALIEN ............................................................................................................ 19
2.1.7 GERÄTE ...................................................................................................................................... 20
2.1.8 DATENVERARBEITUNG .................................................................................................................. 21
2.2 METHODEN ........................................................................................................................ 22
2.2.1 ZELLZÄHLUNG.............................................................................................................................. 22
2.2.2 BLUTENTNAHME BEI MÄUSEN ........................................................................................................ 22
2.2.3 PRÄPARATION VON MÄUSEN ......................................................................................................... 22
2.2.4 ZELLAUFBEREITUNG AUS DEM BLUT................................................................................................. 23
2.2.5 ZELLAUFBEREITUNG AUS DER MILZ.................................................................................................. 24
2.2.6 ZELLAUFBEREITUNG AUS DEM KNOCHENMARK .................................................................................. 26
I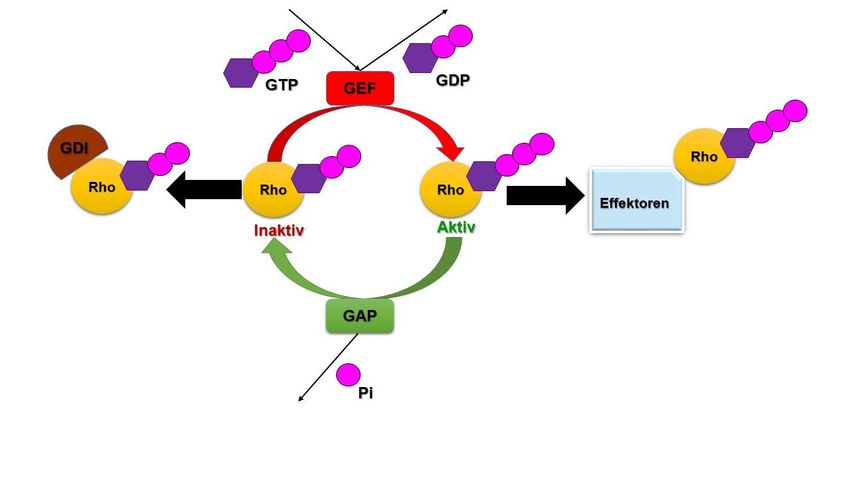
2.2.7 FUNKTIONSWEISE DER DURCHFLUSSZYTOMETRIE ............................................................................... 27
2.2.8 HISTOLOGISCHE FÄRBUNG DER MILZ ............................................................................................... 28
2.2.9 ANALYSE DER ANZAHL UND GRÖßE DER FOLLIKEL .............................................................................. 28
2.2.10 GENOTYPISIERUNG DER MÄUSE.................................................................................................... 29
2.2.11 STATISTISCHE AUSWERTUNG ........................................................................................................ 30
3 AUSWERTUNG .................................................................................................................... 31
3.1 UNTERSUCHTE MAUSMODELLE ................................................................................................ 31
3.2 VAV-PROTEINE BEEINFLUSSEN DIE GRÖßE UND DIE ENTWICKLUNG DER B-ZELLPOPULATION ................... 31
3.2.1 BTK-VAV1-DOPPELDEFIZIENZ FÜHRT ZU EINER ENTWICKLUNGSSTÖRUNG IM KNOCHENMARK ................... 31
3.2.2 BTK-VAV1-DOPPELDEFIZIENZ FÜHRT ZU EINER ENTWICKLUNGSSTÖRUNG IM VENÖSEN BLUT .................... 37
3.2.3 BTK-VAV1-DOPPELDEFIZIENZ UND VAV3-DEFIZIENZ FÜHREN ZU EINER VERÄNDERTEN POPULATION IN DER
MILZ................................................................................................................................................... 42
3.2.4 BTK-VAV1-DOPPELDEFIZIENZ FÜHRT TENDENZIELL ZU EINER ENTWICKLUNGSSTÖRUNG IN DER MILZ .......... 45
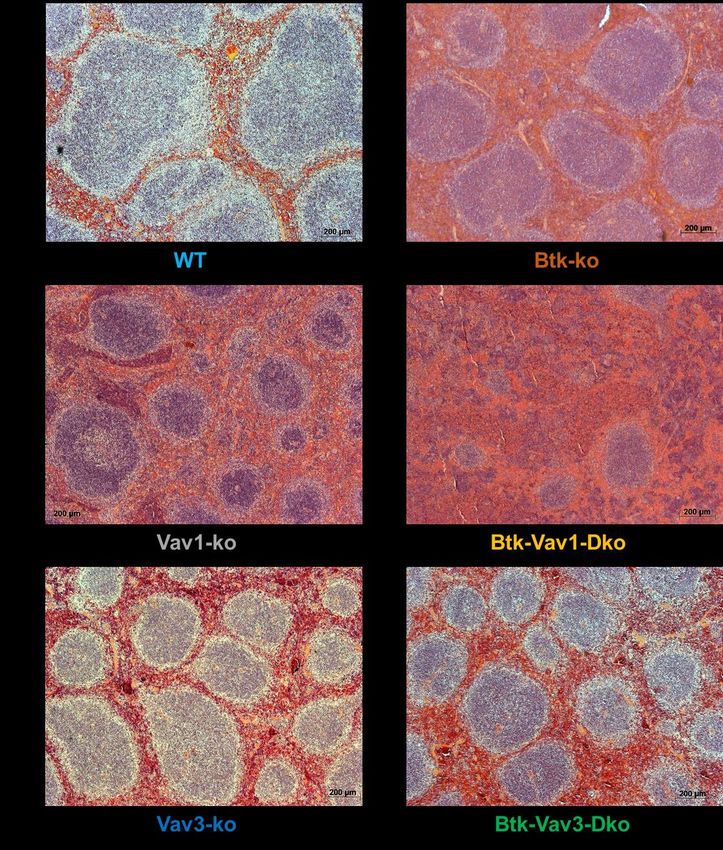
3.3 VAV-DEFIZIENZ FÜHRT ZU EINER GESTÖRTEN MIKROARCHITEKTUR DER MILZ ...................................... 49
4 DISKUSSION ........................................................................................................................ 54
5.1 EINE VAV3-DEFIZIENZ WIRKT SICH AUF DIE B-ZELLENTWICKLUNG IN DER MILZ AUS .............................. 56
5.2 EINE BTK-VAV1-DOPPELDEFIZIENZ WIRKT SICH TENDENZIELL AUF DIE B-ZELLENTWICKLUNG AUS, WÄHREND
EINE EINFACHE VAV1-MUTATION DIESE WEITGEHEND UNBEEINTRÄCHTIGT LÄSST....................................... 58
5.3 VAV-DEFIZIENZEN HABEN NICHT NUR EINFLUSS AUF DIE ENTWICKLUNG VON B-ZELLEN, SONDERN
BEEINFLUSSEN AUCH DIE MIKROARCHITEKTUR/FOLLIKELSTRUKTUR DER MILZ ........................................... 60
5.4 SCHLUSSFOLGERUNG UND AUSBLICK ......................................................................................... 62
5. ZUSAMMENFASSUNG ........................................................................................................ 65
6 LITERATURVERZEICHNIS ...................................................................................................... 67
DANKSAGUNG ....................................................................................................................... 75
LEBENSLAUF .......................................................................................................................... 77
II
Abkürzungsverzeichnis
ABC Animal Blood Counter (englisch)
AC-Domäne Acidic motif (englisch)
AKT Proteinkinasen B
APC Allophycocyanin
APC-Cy7 Allophycocyanin+Cyanin 7
BAFF B-cell activator of the TNF-α family (englisch)
BAFF-R BAFF-Rezeptor
BCR B-Zell-Rezeptor
BLNK, SLP-65, BASH B-cell linker proteins (englisch)
BSA Bovines Serumalbumin
BTK Bruton‘sche-Tyrosinkinase
[Ca2+]i Intrazelluläres Kalzium
CD Cluster of Differentiation (englisch)
CH-Domäne Calponin-homology Domäne
DAG Diacylglycerin
DBL Diffuses B-Zell Lymphom
DNA Desoxyribonukleinsäure
DH-Domäne DBL-homology Domäne
DKo Doppel-knock-out
eANOVA Einfaktorielle Varianzanalyse
III
EDTA Ethylendiamintetraessigsäure
eF450 eFlour 450
ER Endoplasmatisches Retikulum
F4/80 EGF-like module-containing mucin-like hormone
receptor-like 1 (englisch)
FACS Durchflusszytometrie
FCS Fetales bovines Serum
FITC Fluorescein isothiocyanate
Follicular b cell, FBC Follikuläre B-Zelle
GAP GTPase-aktivierendes Protein
GDI, Rho-GDI Guanin-Nukleotid-Dissoziations-Inhibitor
GEF Guanin-Nukleotid-Austauschfaktor
GDP Guanosindiphosphat
Grb2, GRB2 Growth-factor receptor-bound protein-2 (englisch)
GTP Guanosintriphosphat
H-chain Schwere Kette
HPS Hämatopoetische Stammzelle
Ig Immunglobulin
Immature b cell, IBC Unreife B-Zelle
I(1,4,5)P3 Inositoltrisphosphat
ITAM Immunrezeptor-Tyrosin-basiertes Aktivierungsmotiv,
Immunoreceptor tyrosine-based activation motif
(englisch)
Ko Knock-out
L-chain Leichte Kette
IVLy6C Lymphozytenantigen 6C
Ly6G Lymphozytenantigen 6 complex locus G6D
MAPK Mitogen-aktivierte Proteinkinase-Kaskade
Marginal zone b cell; Marginal-Zonen-B-Zelle
MZB
MHC Haupthistokompatibilitätskomplex
mIg membrangebundenes Oberflächen-Immunglobulin,
membranständiges Immunglobulin, B-Zell-Rezeptor
n Anzahl der analysierten Tiere (bei Abbildungen)
NaCl Natriumchlorid
PBS Phosphate Buffer Solution (englisch)
PCR Polymerase-Kettenreaktion
PE Phycoerythrin
PE-Cy5.5 Phycoerythrin-Cyanin 5.5
PE-Cy7 Phycoerythrin+Cyanin 7
PerCP Peridinin chlorophyll
PerCP/Cy 5.5 Peridinin chlorophyll/Cyanin 5.5
PH-Domäne Pleckstrin-homology Domäne
Pi Phosphat
PI-3K Phosphoinositid-3-Kinase
PI(4,5)P2 Phosphatidylinositol-(4,5)-bisphosphat
PI(3,4,5)P3 Phosphatidylinositol-(3,4,5)-trisphosphat
PI(4)P Phosphatidylinositol-(4)-Phosphat
PIP-5K Phosphatidylinositol 4-Phosphate-5 Kinase
VPKC Proteinkinase C
PKCβ Proteinkinase C Beta
PLCγ2 (1-)Phosphatidylinositol-(4,5)-bisphosphat
Phosphodiesterase Gamma-2
PR-Domäne Prolinreiche Domäne
Pre-BCR Prä-B-Zell-Rezeptor
PTK Protein-Tyrosin-Kinase
Ras Rat Sarcoma (englisch)
Rho Ras homologous (englisch)
Rpm Runden pro Minute
RPMI Zellkulturmedium (Roswell Park Memorial Institute)
sec Sekunde
SDS Natriumdodecylsulfat
SH2-Domäne SRC-homology 2 Domäne
SH3-Domäne SRC-homology 3 Domäne
sIg sezerniertes Immunglobulin/Antikörper
SLC Ersatz-Leichte Kette
SYK Spleen Tyrosine Kinase (englisch)
TACI Transmembrane activator and CAML interactor
(englisch)
Transitional b cell, TBC Transitionelle B-Zelle
TRIS Tris(hydroxymethyl)-aminomethan
WBC White blood count (englisch)/Leukozytenanzahl
WT, Wt Wildtyp
VI1 Einleitung
1.1 B-Zell-Entwicklung
1.1.1 Entwicklung im Knochenmark
Unser Körper ist täglich mit einer Vielzahl von Angriffen auf ihn konfrontiert. Um
diesen variantenreichen Angriffen standzuhalten, benötigen wir unter anderem ein
komplexes zelluläres Immunsystem, das sich grob in zwei Lager unterteilen lässt:
das angeborene und das erworbene Immunsystem. In dieser Arbeit beschäftige ich
mich mit einem Bestandteil des erworbenen Immunsystems, genauer gesagt, der
B-Zelle (aus dem Englischen: B= bursa-, oder bone marrow-derived). [33] Um deren
Funktion und Besonderheiten besser verstehen zu können, befasse ich mich zuerst
mit den Entwicklungsschritten dieser speziellen Immunzelle, die für meine Arbeit
von entscheidender Bedeutung sind.
Die Entwicklung der B-Zelle kann grob in einen ersten, Antigen-unabhängigen, und
einen zweiten, Antigen-abhängigen Abschnitt unterteilt werden, der für diese Arbeit
aber eine untergeordnete Rolle spielt.
Im Knochenmark beginnt die Entwicklungsreise, ausgehend von einer
hämatopoetischen Stammzelle, die drei Haupt-Entwicklungsschritte benötigt, bevor
sie das Knochenmark verlassen kann. Die Entwicklungsschritte sind dabei durch
eine charakteristische Expression von Oberflächenmolekülen und der funktionellen
Genumlagerung der Immunglobuline definiert.
Im ersten Entwicklungsstadium, dem sogenannten Pro-B-Zellstadium, beginnt die
DJ-Umlagerung der schweren Kette (H-chain, aus dem Englischen „heavy“), der
sich die V-DJ-Umlagerung anschließt. [40] Anschließend bildet diese erfolgreich
gebildete µ-H-chain zusammen mit einer sogenannten Ersatz-Leichten Kette (SLC)
ein Heterodimer, den sogenannten Prä-B-Zell-Rezeptor (pre-BCR), welcher mit den
signalleitenden Untereinheiten Ig-α und Ig-β (CD79a und CD79b) assoziiert ist.
[35,40]
Dieser pre-BCR führt die B-Zelle nun in das Prä-B-Zell-Entwicklungsstadium und
übernimmt dabei zwei wesentliche Aufgaben. Auf der einen Seite verhindert der
pre-BCR, dass die B-Zelle zwei schwere Ketten mit zwei verschiedenen Spezifitäten
1bildet – dies wird auch als „Allelic exclusion“ bezeichnet. [50] Auf der anderen Seite
initiiert der pre-BCR die V-J-Genumlagerung in der leichten Kette (L-chain, aus dem
Englischen light). Dies führt schlussendlich dazu, dass die Zelle ein erstes
funktionierendes IgM-Molekül bzw. BCR auf ihrer Oberfläche exprimiert und sich ab
sofort in ihrem letzten Entwicklungsstadium im Knochenmark befindet, welches als
unreifes B-Zellstadium (immature B cell) bezeichnet wird. [40]
1.1.2 Periphere Reifung
Nun wandern diese, jetzt als Transitionelle B-Zellen (transitional b cell)
bezeichneten B-Zellen aus dem Knochenmark in das Blut aus, um auf ihrer weiteren
Reifungsreise als nächste Station periphere sekundäre lymphoide Organe, wie die
Milz, zu erreichen.
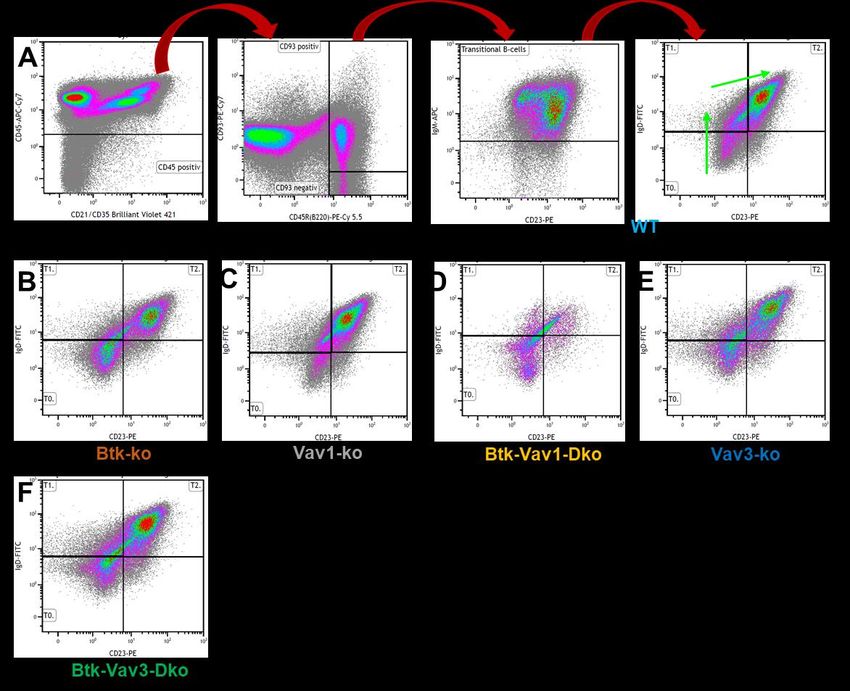
Hier habe ich in dieser Arbeit die verschiedenen Stadien der Population der
Transitionellen B-Zellen in T0-T2 eingeteilt, wobei die Einteilung anhand der
Expression verschiedener Oberflächenmoleküle erfolgte (vergleiche Abbildung 1).
Entscheidend für diese Arbeit und unsere Beobachtungen ist der Wechsel vom T1-
in das T2-Stadium, der durch zwei Signalwege eingeleitet wird. Zum einen muss die
B-Zelle in diesem Stadium einen ab sofort funktionierenden B-Zell-Rezeptor (BCR)
aufweisen. [11] Zum anderen kommt es zur Aktivierung durch B-cell activator of the
TNF-α family (BAFF) und dem auf der B-Zelle exprimierten BAFF-Rezeptor
(BAFF-R). Dieser BAFF-R wird erstmals im Stadium der immature B cell im
Knochenmark auf der Oberfläche exprimiert und ist ein wichtiger Signalweg für das
Überleben und die Entwicklung der B-Zelle. [4,36] Für die daran anschließende
Aufteilung der B-Zellen in den letzten Antigen-unabhängigen Entwicklungsschritt
sind nun neben einem funktionierenden BCR und BAFF-R ebenfalls ein sog.
Transmembrane activator and CAML interactor (TACI) vonnöten. Dieser scheint
besonders entscheidend für die Weiterentwicklung der T2-B-Zelle zu der
Marginal-Zonen-B-Zelle (marginal zone b cell; MZB) zu sein, anders als bei der
Follikulären B-Zelle (follicular b cell, FBC). [16,22]
2Abbildung 1: Schematische Darstellung der Antigen-unabhängigen B-Zellentwicklung in Knochenmark und Milz. Die
Differenzierung der verschiedenen Entwicklungsstadien erfolgt anhand der Expression unterschiedlicher
Oberflächenmoleküle, die jeweils neben dem Entwicklungsstadium aufgelistet sind. Die Grafik wurde nach dem Wissen der
aktuellen Literatur erstellt (von Chung et al., Trends Immunol., 2003 [11] sowie Bemark, JBR, 2015 [5]).
FBC = Follikuläre B-Zelle, HPS = Hämatopoetische Stammzelle, IBC = unreife B-Zelle, MZB = Marginal-Zonen-B-Zelle,
Pro = Pro-B-Zellstadium, Pre = Prä-B-Zellstadium, TBC = Transitionelle B-Zelle,
T0-2 = verschiedene Stadien der Transitionellen B-Zelle in der Milz.
Die Abbildung wurde selbst erstellt.
Da ich in meiner Arbeit nur diesen Teil der B-Zellentwicklung untersucht habe und
sich der nun T-Zell-abhängige Teil der B-Zellentwicklung anschließt, gehe ich auf
diesen Entwicklungsschritt nur kurz ein, ehe ich mich näher mit dem bereits
angesprochenen BCR und seinen nachgeschalteten Signalwegen beschäftige.
Die FBC der Milz und auch der Lymphknoten werden in ihrem nächsten
Reifungsschritt mit Hilfe von T-Zellen aktiviert, indem diese ihnen Antigene
präsentieren. Durch diese Aktivierung werden zunächst solche B-Zellen positiv
selektioniert, die einen für das vorliegende Antigen „passenden“ B-Zell-Rezeptor
(=membranständiges Immunglobulin (mIg)) aufweisen. Zusätzlich wird eine
Enzymmaschinerie in Gang gesetzt, die als Folge zur somatischen Hypermutation
der variablen Region des membranständigen Immunglobulins
(mIg = B-Zell-Rezeptor) bzw. sezernierten Immunglobulins (sIg = Antikörper) führt.
Diese somatische Hypermutation, kombiniert mit der Fähigkeit zum Klassenwechsel
der Immunglobuline, führt zu einer veränderten und einzigartigen Affinität dieser auf
das jeweilige Antigen. Schließlich können diese Zellen dann ihre Funktion als
3Antikörper-produzierende Plasmazellen oder als langlebige Gedächtniszellen (v.a.
B-Zellen aus den Keimzentren) übernehmen. [33,40]
1.2 B-Zell-Rezeptor (BCR)-nachgeschaltete Signalwege
Wie unter 1.1 bereits dargelegt, ist ein funktionierender BCR entscheidend für die
Differenzierung und das Überleben der B-Zelle. In diesem Abschnitt wird nun
genauer auf den Aufbau und die intrazelluläre Wirkungsweise dieses
Transmembranproteinkomplexes eingegangen. Da nicht nur die B-Zelle selbst,
sondern auch ihr BCR weitreichende und differenzierte Aufgaben übernimmt, wäre
eine komplette Darstellung der BCR-nachgeschalteten Signalwege an dieser Stelle
nicht zielführend. Deshalb werden in diesem Abschnitt die für diese Arbeit und ihre
Ergebnisse wichtigsten, bisher bekannten Signalwege dargestellt und erläutert.
Auch wenn es in dieser Arbeit hauptsächlich um den Antigen-unabhängigen
Entwicklungsschritt der B-Zellentwicklung geht, ist doch die spezifische und hoch
vielfältige Antigen-Bindungsfähigkeit eines der Auszeichnungsmerkmale der
B-Zelle. Diese hohe Heterogenität entsteht durch die in 1.1 beschriebene
VDJ-Umlagerung der schweren Kette und der VJ-Umlagerung der leichten Kette
und führt schließlich zu einer einzigartigen Bindungsstelle jeder B-Zelle, dem
membrangebundenen Oberflächen-Immunglobulin (mIg). Dieses Immunglobulin
allein besitzt allerdings keine Kinase-Aktivität, sondern nutzt hierzu die bereits
angesprochenen nicht-kovalenten Heterodimerproteine Ig-α und Ig-β. Jedes dieser
Proteine besitzt ein sog. Immunrezeptor-Tyrosin-basiertes Aktivierungsmotiv
(Immunoreceptor tyrosine-based activation motif; ITAM), welches nach
BCR-Bindung und -Konformationsänderung schließlich die BCR-nachgeschalteten
Signalwege in Gang setzt. [17,47]
Hierzu wird mittels der Protein-Tyrosin-Kinase (PTK) Lyn eine Phosphorylierung der
ITAM in Gang gesetzt, die daraufhin eine Bindungsstellung für Proteine mit
SH-2-Domäne schaffen. Hierzu zählt neben einer weiter verstärkten Rekrutierung
von Lyn auch SYK (spleen tyrosine kinase). [13,47] Um weitergehende Signalwege
zu aktivieren, werden weitere Proteine benötigt, die u.a. durch eine Aktivierung der
Phosphoinositid-3-Kinase (PI-3K) nach BCR-Stimulation und der damit
4verbundenen Phosphorylierung des Zellmembranbestandteils Phosphatidylinositol-
(4,5)-bisphosphat (PI(4,5)P2) zu Phosphatidylinositol-(3,4,5)-trisphosphat
(PI(3,4,5)P3) rekrutiert werden können. [13] Ein bekannter Mechanismus zur
Aktivierung der gerade angesprochenen PI-3K ist das integrale
Transmembranglykoprotein CD19. Dieses wird nach BCR-Aktivierung an
bestimmten Sequenzen (Tyr-X-X-Met) seines zytoplasmatischen Anteils durch Lyn
phosphoryliert und bietet somit eine Bindungsstelle für Proteine mit SH2-Domäne
wie z.B. für die p85-Bindungsuntereinheit der PI-3K. [21,57] Des Weiteren führt
diese Bindung zum einen dazu, dass die PI-3K sich in unmittelbarer Nähe zu ihrem
Substrat an der Zellmembran befindet. Zum anderen wird durch die Bindung die
Aktivität der katalytischen p110-Untereinheit der PI-3K verstärkt. [21] Durch die
Aktivität der PI-3K und ihrem Produkt, dem PI(3,4,5)P3, können nun
Effektorproteine, wie die Bruton‘sche-Tyrosinkinase (BTK), die
Proteinkinasen B (AKT), die (1-)Phosphatidylinositol-(4,5)-bisphosphat
Phosphodiesterase Gamma-2 (PLCγ2) und die Proteinkinase C-Beta (PKCβ),
rekrutiert und gebunden werden. [46,47] PI(3,4,5)P3 bietet außerdem den weiteren
Vorteil, dass alle Effektorproteine und ihre Interaktionspartner in räumlicher Nähe
zueinander gebracht werden, wodurch die signaltransduzierenden Effekte
ausgelöst werden können. [18]
amit es jedoch zu dem gerade geschilderten Zustand kommen kann, werden an
dieser Stelle noch zwei wichtige Proteine benötigt, die u.a. SYK und PLCγ2
verbinden, diese wurden pp70 und pp68 genannt und erhielten zusammen
schließlich den Namen BLNK (B-cell linker proteins). [19] BLNK
(oder auch als SLP-65 oder BASH bezeichnet) wird kurz nach BCR-Aktivierung
durch SYK phosphoryliert. In der Folge ist es maßgeblich nicht nur für die
Rekrutierung der PLCγ2 zur Zellmembran zuständig, sondern auch für weitere
Aktoren des BCR-nachgeschalteten Signalweges, wie z.B. Vav. [19] Anschließend
kann dieser so gebildete Komplex aus u.a. BLNK, SYK, LYN, PI-3K, Vav und PLCγ2
als Signalsom bezeichnet werden, der in der Lage ist, das einzelne Signal der
BCR-Aktivierung in verschiedene Signalwege zu verzweigen. [55]
Ein Beispielsignalweg ist die folgende stattfindende doppelte Phosphorylierung von
PLCγ2 durch SYK und BTK, wobei die Phosphorylierung durch BTK die Effektivität
5der PLCγ2 sogar noch steigert. [10,18] Die PLCγ2 hat nun die wichtige Aufgabe,
aus PI(4,5)P2, dem Substrat, das auch von der PI-3K benötigt wird, die Second
Messenger Inositoltrisphosphat (I(1,4,5)P3) und Diacylglycerin (DAG) zu
generieren. Um das Substrat in größeren Mengen zur Verfügung gestellt zu
bekommen, nutzt die B-Zelle eine weitere Kinase, die
Phosphatidylinositol 4-Phosphate-5 Kinase (PIP-5K). Diese phosphoryliert
Phosphatidylinositol-(4)-Phosphat PI(4)P an der D-5-Position seines Inositolringes,
um PI(4,5)P2 zu generieren. [18] Wichtige Akteure in diesem Rahmen sind zum
einen die BTK – diese fungiert als eine Art Shuttle, um die PIP-5K zur Zellmembran
und damit auch in die direkte Umgebung des Signalosoms zu befördern. [43] Auf
der anderen Seite haben wir die Vav-Proteine, auf die in einem anderen Punkt
genauer eingegangen wird, und hier v.a. Vav1, welches eine Interaktion mit der
GTPase Rac eingeht. [55] Rac, welches als GTPase in der Lage ist,
Guanosintriphosphat zu binden und zu hydrolisieren, ist ein bekannter wichtiger
Aktivator der PIP-5K und damit unmittelbarer Akteur in dem eben beschriebenen
Signalweg, der zur Substratbereitstellung der PLCγ2 dient. [52]
Doch was folgt nun nach der Generierung der zuvor erwähnten Second
Messengern? I(1,4,5)P3 bindet u.a. an spezifische Rezeptoren, z.B. auf dem
endoplasmatischen Retikulum, und führt in der Folge zu einer intrazellulären
Kalzium-Freisetzung aus diesem intrazellulären Ca2+-Speicherkompartiment. [48]
Durch diese gesteigerte intrazelluläre Kalziumkonzentration wiederum ist es der
Zelle möglich, viele verschiedene nachgeschaltete Effekte, wie z.B. den
Transkriptionsfaktor NF-κB, zu aktivieren. [13] Der Second Messenger DAG
hingegen hat als Ziel, die Gruppe der Proteinkinasen C zu aktivieren. Hier wirkt DAG
nach Kalziumbindung an der Proteinkinase C (PKC) als Aktivator der selbigen, die
daraufhin u.a. den MAP-Kinase-Signaltransduktionsweg
(Mitogen-aktivierte Proteinkinase-Kaskade, MAPK) reguliert, der viele
regulatorische Aufgaben in der Zelle übernimmt. [13,32]
Des Überblickes wegen folgt abschließend zu diesem Kapitel eine schematische
Darstellung des hier beschriebenen BCR-nachgeschalteten Signalwegs
(s. Abbildung 2) in vereinfachter Form. Wie aus dem eben dargestellten ersichtlich
ist, schließen sich viele verschiedene weitere Signalwege und nachgeschaltete
6Effekte an, deren genaue Darstellung und Erläuterung den Umfang dieser
Einleitung sprengen würden. Vielmehr widmen sich die folgenden Abschnitte nun
wichtigen Proteinen, die in dieser Arbeit untersucht wurden: den Vav-Proteinen und
der BTK.
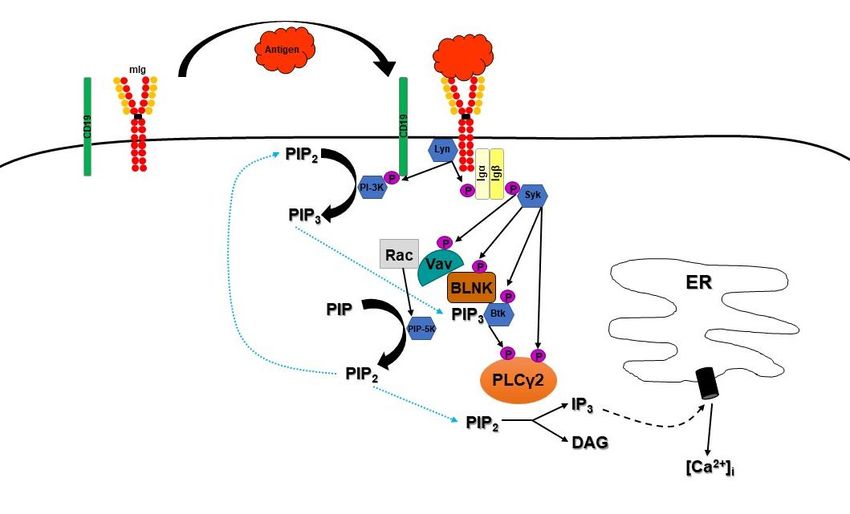
Abbildung 2: Schematisch vereinfachte Darstellung der im Text aufgeführten Signalwege nach Antigenbindung an den BCR
(eigene Darstellung nach dem aktuellen Wissensstand). Nach Antigenbindung an den Oberflächenrezeptor wird durch Lyn
das sog. ITAM (bestehend aus Igα und Igβ) phosphoryliert. Gleichzeitig phosphoryliert Lyn noch den Co-Rezeptor CD19, der
danach eine Bindungsstelle für PI-3K darstellt und damit das wichtige Substrat PIP3 in der Nähe des Signalosoms produzieren
kann. Über das phosphorylierte ITAM kann nun Syk rekrutiert werden, welches in der Folge verschiedene weitere Proteine
aktiviert, z.B. BLNK. Dieses Bindeprotein bildet auch gleichzeitig eine Art Plattform für die weiteren Effektoren des sich so
bildenden Signalosoms, bestehend u.a. aus Vav, Btk und PLCγ2. Das gebundene und aktivierte Vav interagiert nun u.a. mit
Rac, um die PIP-5K zu aktivieren, die in der Folge das Substrat für die PI-3K, aber auch die PLCγ2 liefert. Die PLCγ2 stellt
die entscheidende Weiche in dieser Darstellung dar, da sie IP3 für die intrazelluläre Kalziumfreisetzung und DAG für die
Aktivierung und Beeinflussung weiterer nachgeschalteter Effekte produziert.
BLNK = B-cell linker proteins, Btk = Bruton’sche Tyrosinkinase, [Ca2+]i = intrazelluläres Kalzium, DAG = Diacylglyzerin,
ER = Endoplasmatisches Retikulum, ITAM = Immunoreceptor tyrosine-based activation motif (bestehend aus Igα und Igβ),
IP3 = Inositoltrisphosphat, mIg = membranständiges Immunglobulin/B-Zell-Rezeptor, PI-3K = Phosphoinositid-3-Kinasen,
PI-5K = Phosphatidylinositol 4-Phosphate-5-Kinase, PIP = Phosphatidylinositol 4-phosphat,
PIP2 = Phosphatidylinositol-(4,5)-bisphosphat, PIP3 = Phosphatidylinositol-(3,4,5)-triphosphat,
PLCγ2 = (1-)Phosphatidylinositol-(4,5)-bisphosphat-phosphodiesterase Gamma-2.
Die Abbildung wurde selbst erstellt.
71.3 Vav-Proteine
1.3.1 Vav-Überblick
Beginnen werde ich mit den bereits im Titel dieser Arbeit angesprochenen Vav-
Proteinen und einer Anekdote zu ihrem Namensursprung. Manche bahnbrechende
Entdeckung der Menschheitsgeschichte beruht auf einem Missgeschick oder einem
Zufall – beispielsweise die Entdeckung des Penicillins durch Sir Alexander Fleming.
Und auch die Vav-Proteine wurden eher zufällig entdeckt, als eine Gruppe um den
spanischen Forscher Mariano Barbacid 1989 ihre Forschungsergebnisse
veröffentlichten. Auf der Suche nach sog. „transforming genes“ in menschlichen
Ösophaguskarzinom-Zellen, gelang ihnen die Isolation und Charakterisierung
dieser Proteine, die sie damals schon richtigerweise als Onkogene betitelten. [28]
Der Name „vav“ stellt eine Hommage des Erstautors an seine Muttersprache, dem
Hebräischen, dar: Da die Vav-Proteine das sechste in diesem Labor identifizierte
Onkogen darstellten, wurden sie nach dem sechsten Buchstaben aus dem
hebräischen Alphabet „vav“ benannt. [28] Nach der Entdeckung dieses Onkogens,
folgten einige Jahre später die Entdeckung zweier weiterer Vertreter dieser
Proteinfamilie, die Vav2 [23] und Vav3 [37] getauft wurden, so dass das ursprünglich
entdeckte und auch heute noch am besten erforschte Vav-Protein die Bezeichnung
Vav1 erhielt.
Während sich die Expression von Vav1 auf das hämatopoetische System allein
beschränkt, werden Vav2 und Vav3 auch in anderen Körperzellen exprimiert. [55]
Da sich Vav2 und Vav3 in größeren Teilen strukturell Vav1 gleichen und dieses
auch am bisher besten erforscht ist, bietet es sich an, anhand der Struktur von Vav1
einen groben Überblick über die verschiedenen bekannten Funktionen und
Interaktionen der Vav-Proteine zugeben. [49,55]
8Abbildung 3: Schematische Darstellung des Vav-Proteins am Beispiel von Vav1 (eigene Darstellung nach dem aktuellen
Wissensstand). Während die DH-Domäne für die GEF-Funktion des Proteins zuständig ist, wird sie von den umliegenden
Domänen in ihrer Aktivität reguliert (s. 1.3.2). Die PH-Domäne ist darüber hinaus noch zuständig für die Interaktion mit u.a.
von PI-3K generierten Phospholipiden und damit zur Rekrutierung des Proteins in die direkte Umgebung seiner
Interaktionspartner zum Signalosom. Die C-terminalen Abschnitte des Proteins sind für die Interaktion mit unterschiedlichen
Partnern zuständig, von denen einige in der Grafik aufgeführt sind.
AC = Acidic motif, Btk = Bruton’sche-Tyrosinkinase, CH = Calponin-homology Domäne, DBL = Diffuses B-Zell Lymphom,
DH = DBL-homology Domäne, GEF = Guanin-Nukleotid-Austauschfaktor, GRB2 = Growth-factor receptor-bound protein-2,
PH = Pleckstrin-homology Domäne, PR = Prolinreiche Domäne, SH2 = SRC-homology 2 Domäne,
SH3 = SRC-homology 3 Domäne, SYK = Spleen Tyrosine Kinase.
Die Abbildung wurde selbst erstellt.
1.3.2 GEF-Funktion
Die Vav-Proteine sind an vielen verschiedenen Aufgaben im Organismus beteiligt,
wobei eine ihrer wichtigsten und zugleich am besten untersuchten Funktion in 1.2
bereits angesprochen und hier kurz skizziert wird: Sie gehören zur sog.
DBL-Familie (Diffuses B-Zell Lymphom-Familie) der Rho-GEFs [55] und
übernehmen hier als Guanin-Nukleotid-Austauschfaktor
(„guanine nucleotide exchange factor“; GEF) die wichtige Aufgabe des
namensgebenden Austausches von GTP und GDP für die GTPasen aus der Ras
homologous (Rho)-Familie. [12] GTPasen sind eine große Proteingruppe, die man
sich als eine Art „molecular switch“ vorstellen kann. [6] Mit wichtigen Vertretern wie
RhoA, Rac1 and Cdc42 sind sie in der Lage, je nach gebundenem Zustand, mit
vielen verschiedenen zellulären Effektoren oder Zielproteinen zu interagieren. [6,12]
Hierbei ist ein aktiver, GTP-gebundener Zustand, von einem inaktiven, GDP-
gebundenen Zustand zu unterscheiden. In ihrem aktivierten Zustand können sie
viele verschiedene zelluläre nachgeschaltete Signalwege mitregulieren und so
9direkt Einfluss auf z.B. die Gentranskription, Zellproliferation, Migration,
Differenzierung und das Überleben nehmen. [55] Um diese Regelfunktion zu
erfüllen, greifen auf der einen Seite die Vav-Proteine als Rho-GEFs ein und
übernehmen die wichtige Austauschfunktion zur Aktivierung dieser Rho GTPasen.
[6] Auf der anderen Seite gibt es die sog. Guanin-Nukleotid-Dissoziations-Inhibitor
(„guanine nucleotide dissociation inhibitors“; Rho-GDIs) als eine Art Gegenspieler
der Rho-GEFs, denn sie binden zytoplasmatische
GDP-gebundene GTPasen und verhindern damit den Austausch von GDP mit GTP.
[6,38] (s. Abbildung 4)
Abbildung 4: Schematische Darstellung der Guaninnukleotidaustauschfunktion der VAV-Proteine (eigene Darstellung nach
dem aktuellen Wissensstand).
GDI = Guanin-Nukleotid-Dissoziations-Inhibitor, GDP = Guanosindiphosphat,
GEF = Guanin-Nukleotid-Austauschfaktor, GTP = Guanosintriphosphat, GAP = GTPase-aktivierendes Protein,
Pi = Phosphat, Rho = Ras homologous.
Die Abbildung wurde selbst erstellt
Diese Aufgabe der GEF-Aktivität wird durch die DH-Domäne der Vav-Proteine
ermöglicht. Doch jeder Regelkreis benötigt auch einen Regler, weswegen diese
Domäne auf der N-terminalen Proteinseite von der CH- und AC-Domäne und auf
der C-terminalen Proteinseite von der PH- und C1-Domäne flankiert wird. [58]
10Die Funktion der CH-Domäne wird kontrovers diskutiert und ist bis heute nicht
endgültig geklärt. Sicher ist, dass sie anders als CH-Domänen bei anderen
Proteinen, bei den Vav-Proteinen nicht für eine Interaktion mit F-Aktin zur Verfügung
steht. [27] Sie scheint allerdings noch nicht definitiv geklärte Einflüsse auf die
Autoinhibition der GEF-Aktivität der DH-Domäne und Auswirkungen auf die
intrazelluläre Kalziumfreisetzung zu besitzen. [27,55] Zum aktuellen Zeitpunkt wird
davon ausgegangen, dass die CH-Domäne eine Interaktion mit der C1-Domäne
eingeht, um damit die inhibitorische Eigenschaft der AC-Domäne zu verstärken.
[41,58]
Diese AC-Domäne übt ihre inhibitorische Eigenschaft durch drei regulatorische
Tyrosinregionen aus: Y142, Y160 und Y174. [27] Allen drei Regionen gemeinsam
ist, dass sie die Sequenz φYXXφ beinhalten (φ = Ile, Leu, oder Val und
X = hydrophil). [1] Hierdurch ist es ihnen möglich, in ihrem unphosphorylierten
Zustand die GEF-Aktivität der DH-Domäne zu blockieren, wobei Y174 die stärkste
Wirkung nachgesagt wird und Y142 und Y160 lediglich als Verstärker vermutet
werden. [1,62]
Auch um die genaue Funktion und Auswirkungen der PH- und C1-Domäne wird seit
Jahren kontrovers diskutiert. Die Autoren sind sich einig, dass diese beiden
Domänen essentielle Auswirkungen auf die Aktivität der DH-Domäne besitzen, doch
wie genau, bleibt weiterhin ungeklärt. Die Fähigkeit der PH-Domäne mit PI(4,5)P2
und PI(3,4,5)P3 zu interagieren, wird als Möglichkeit der Rekrutierung der Vav-
Proteine zum Signalosom und zur Beeinflussung auf die DH-Domäne zwar logisch
vermutet, konnte aber abschließend nicht sicher geklärt werden. [14,27,41,55,58]
So bleibt weiterhin definitiv ungeklärt, auf welchem genauen Weg diese Domänen
die Aktivitäten der Vav-Proteine beeinflussen und auch weitere, noch unbekannte
Interaktionspartner sind nicht ausgeschlossen.
1.3.3 Interaktionspartner der Vav-Proteine
Der C-terminale Abschnitt der Vav-Proteine steht ganz im Zeichen der Interaktion
mit weiteren Akteuren des Signalosoms in der B-Zelle und deren nachgeschalteten
Signalwegen. Die prolinreiche Region ist mit der ersten SH3-Domäne zusammen
im Stande, z.B. mit der für diese Arbeit wichtigen Bruton’schen Tyrosinkinase zu
11interagieren. [55] Die darauffolgende SH2-Domäne ist in der Lage, die Vav-Proteine
an thyrosinphosphorylierte Regionen zu binden, welches eine der postulierten
Möglichkeiten ist, die Vav-Proteine, wie unter 1.2 geschildert, z.B. an BLNK zu
binden und damit in der Reaktionskette des Signalosoms zu verankern. [27]
1.4 Bruton’sche Tyrosinkinase (Btk)
Die zweite Proteingruppe, die in dieser Arbeit untersucht wurde, ist die Bruton’sche
Tyrosinkinase (Btk). Diese zytoplasmatische „non-receptor“-Tyrosinkinase gehört
zur Familie der Tec-Kinasen, die in verschiedensten Zellen des hämatopoetischen
Systems eine Rolle spielen und deren wichtige Funktion für die B-Zellentwicklung
und -Funktion unter 1.2 bereits angesprochen wurde. [7] Für diese Arbeit wurde die
Btk als eine Art vergleichendes Protein hinzugezogen – da Mutationen dieser
Kinase zu distinkten und vielfach untersuchten Krankheitsbildern führen und damit
einige wichtige Signalwege der Btk gut untersucht sind.
Die wichtigste Erkrankung beim Menschen, die durch eine Mutation der Btk
verursacht wird, ist die X-chromosomal vererbte Agammaglobulinämie [54,60], nach
deren Erstbeschreiber Ogden C. Bruton, die Btk ihren heutigen Namen erhalten hat.
[8] Mutationen wurden in allen Exons der Btk beschrieben und führen in der Folge
zu einem Expressions- bzw. Funktionsverlust der Btk. [44] Mittlerweile wurden über
600 verschiedene Mutationen im Btk-Gen identifiziert. [63] Bei der Maus gibt es eine
ähnliche, mit einem milderen Phänotyp vergesellschaftete Erkrankung. Diese wird
durch eine spontane Btk-Mutation in einem bestimmten Mausstamm ausgelöst,
welche auch beim Menschen identifiziert wurde (R28C) –
die X-chromosomal vererbte Immundefizienz. [42,51] Beide Erkrankungen sind
bedingt durch einen Verlust der Btk-Expression oder durch eine Mutation, die zur
Funktionsänderung in einer der Btk-Subdomänen führt. [44] Dies führt bei den
betroffenen Patienten bzw. bei der Maus zu einem Defekt der B-Zellentwicklung mit
einem Block auf dem Prä-B-Zellstadium (vergleiche Abbildung 1) sowie zusätzlich
zu einer Reduktion von reifen B-Zellen im peripheren Blut und den sekundären
lymphoiden Organen. Dies hat eine massiv reduzierte Konzentration von
Serumimmunglobulinen zur Folge. [7,44]
12Durch die bisherige jahrelange Forschung an diesen Krankheitsbildern und der
Tatsache, dass Btk vom Pro-B-Zellstadium bis zur reifen B-Zelle exprimiert wird, ist
sie ein geeigneter Kandidat, um Auswirkungen der BCR-vermittelten Signalweiter-
leitung auf die B-Zellentwicklung und -Funktion zu erforschen. [7] Dies zeigt sich
auch an der Tatsache, dass sie ein beliebtes Ziel in der aktuellen Therapie von
solchen malignen Erkrankungen der B-Zelle darstellt, die durch eine konstitutiv-
aktive, BCR-vermittelte Signaltransduktionskaskade gekennzeichnet sind. [9]
Außerdem konnte in 1.2 bereits gezeigt werden, dass sie ein wichtiger Teil des
BCR-induzierten Signalosoms sowie des BCR-nachgeschalteten Signalwegs
darstellt, mit vielfältigen Auswirkungen auf die Zellphysiologie, wie z.B. Proliferation
und Apoptose. [34]
131.5 Ziel der Arbeit
Ziel dieser Arbeit ist die weiterführende Erforschung der Rolle der Vav-Proteine auf
die B-Zellentwicklung und die BCR-vermittelte Signalweiterleitung. Wie in den
oberen Punkten beschrieben, wird bisher in der Forschung die Funktion der Vav-
Proteine in diesen Punkten vor allem durch ihre Funktion im Singalosom gesehen,
in dem die Btk eine der wichtigen Endschritte darstellt. Wir schlussfolgerten
dadurch, dass die Vav-Proteine zusammen mit der Btk in einem der
BCR-Aktivierung nachgeschalteten Signalweg zusammen agieren und fungieren.
Um dem Vav-Signalweg weiter auf den Grund zu gehen, untersuchten wir neben
Btk und Vav1 auch Vav3, über dessen Funktion im Immunsystem und in den B-
Zellen noch viele Unklarheiten bestehen.
Dadurch ergaben sich die folgenden Fragestellungen:
1.) Welche Auswirkungen haben Vav1- bzw. Vav3-Mutationen auf die
Populationsverteilung in den unterschiedlichen B-Zellentwicklungsstadien
(vergleiche dazu Abbildung 1)?
2.) Welche Konsequenzen haben Vav1- bzw. Vav3-Mutationen auf die
Mikroarchitektur sekundärer lymphatischer Organe am Beispiel der Milz?
3.) Durch welche molekularen Mechanismen sind mögliche gefundene
Auffälligkeiten erklärbar?
142 Material und Methoden
2.1 Material
2.1.1 Mauslinien
Für die vorliegende Arbeit wurden ausschließlich männliche Tiere analysiert.
Analysiert wurden hierbei Btk-knock-out Mäuse (Btk Y/-), Vav1-knock-out Mäuse
(Vav1 -/-), Vav3-knock-out Mäuse (Vav3 -/-) sowie Btk-Vav1-Doppel-knock-
out Mäuse (Btk Y/-, Vav1 -/-) und Btk-Vav3-Doppel-knock-out Mäuse
(Btk Y/-, Vav3 -/-). Für die Analyse in der Durchflusszytometrie wurden Tiere im Alter
von 7-15 Wochen verwendet, bei der Analyse der Btk-knock-out und
Btk-Vav3-Doppel-knock-out Mäuse kamen Tiere bis zum Alter von 29 Wochen zum
Einsatz. In der histologischen Untersuchung wurden Tiere im Alter von
9-13 Wochen untersucht. Die Chimären wurden durch Kreuzung der einzelnen
Knock-out Mäuse gezüchtet. Als genetischer Hintergrund wurde für alle
untersuchten Mäusestämme ein C57BL/6J-Hintergrund verwendet.
Bei der Btk-knock-out Maus wurde mittels gene targeting die Exone 13 und 14 des
Btk-Gens mit einer Neomycin-Kassette modifiziert. Dies führt zu einer Deletion des
open reading frames sowie der Einführung eines zusätzlichen Stop-Codons und
damit zu einem vollständigen Fehlen der Btk-Proteinexpression. [29]
Für die Vav1-Knock-out Maus wurde ein 14,2 kb DNA-Abschnitt des Vav1-Gens
lokalisiert und mit dem 9,65 kbp Vektor pAZ10 transfiziert. Dieser Vektor enthält
eine Neomycin-Kassette, der zur Deletion des open reading frames mit nachfolgend
fehlender Genexpression von Vav1 führt. [65]
Bei der Vav3-Knock-out Maus wurde das 5‘-Ende des Vav3-Lokus mit einer
Neomycin-Kassette genetisch modifiziert, dies führt zu einer Deletion des open
reading frames mit nachfolgend fehlender Genexpression von Vav3. [45]
Zusätzlich wurden für die beschriebenen Experimente wildtypische
C57BL/6J-Mäuse mituntersucht.
Die Mäuse wurden im Tierstall an der Universität Ulm gezüchtet und gentypisiert.
Die Haltung erfolgte nach SFP-Bedingungen. Alle Untersuchungen an Tieren
wurden dem Regierungspräsidium Tübingen angezeigt (Mitteilung nach § 4 Abs. 3
Tierschutzgesetz; Lizenzen 0.110, 0.221).
152.1.2 Chemikalien und Reagenzien
Folgende Chemikalien und Reagienzen wurden für die Experimente verwendet:
❖ 10 bp DNA ladder, Fa. PeqLab, Deutschland
❖ Ammoniumchlorid, Fa. Sigma, Deutschland
❖ Ampuwa, Fa. Fresenius Kabi, Deutschland
❖ Aqua dest., Fa. Fresenius Kabi, Deutschland
❖ SeaKem® LE Agarose, Fa. Lonza, Schweiz
❖ BSA, Fa. PAA, Österreich
❖ Dinatriumhydrogenphosphat, VWR International GmbH, Deutschland
❖ EDTA-Dinatriumsalz-Dihydrat, Fa. AppliChem Panreac, USA
❖ Eosin G-Lösung 0,5% wässrig, Fa. Carl Roth, Deutschland
❖ Eisessig, VWR International GmbH, Deutschland
❖ Ethanol, Fa. Sigma-Aldrich, Deutschland
❖ FACS lysing solution, Fa. BD, Deutschland
❖ FCS, Fa. Biochrom, Deutschland
❖ Formaldehyd-Lösung, Fa. Otto Fischar GmbH&Co. KG, Deutschland
❖ Glutamin, Fa. Biochrom, Deutschland
❖ Hämalaunlösung sauer nach Mayer, Fa. Carl Roth, Deutschland
❖ Kaliumchlorid, VWR International GmbH, Deutschland
❖ KAPA Taq PCR Kits, Fa. PeqLab, Deutschland
❖ Kohlenstoffdioxid, Linde AG - Gases Division Germany, Deutschland
❖ Midori Green, Fa. Biozym, Deutschland
❖ Murines IgG, Fa. Jackson ImmunoResearch, USA
❖ Natriumchlorid, Fa. Sigma-Aldrich, Deutschland
❖ Natriumazid, Fa. AppliChem, Deutschland
❖ Paraffin, VWR International GmbH, Deutschland
❖ Penicillin, Fa. Biochrom, Deutschland
❖ PBS (1x), Fa. Gibco by life technologies, USA
❖ RNAse freies Wasser, Fa. QIAGEN, Deutschland
❖ Roti®-Histokitt II, Fa. Carl Roth, Deutschland
❖ RPMI Medium 1640, Fa. ThermoFisher Scientific, USA
❖ Streptomycin, Fa. Biochrom, Deutschland
❖ TRIS, Fa. Carl Roth, Deutschland
162.1.3 Puffer
Tabelle 1: Verwendete Puffer und ihre Zusammensetzung
Nachfolgend gelistet sind die verwendeten Puffer für die Experimente und ihre Zusammensetzung.
BSA = Bovines Serumalbumin, EDTA = Ethylendiamintetraessigsäure, FACS = Durchflusszytometrie,
FCS = Fetales bovines Serum, NaCl = Natriumchlorid, PBS = Phosphate Buffer Solution,
RPMI = Zellkulturmedium (Roswell Park Memorial Institute), SDS = Natriumdodecylsulfat,
TRIS = Tris(hydroxymethyl)-aminomethan
FACS-Puffer 500 ml 1x PBS
(500 ml) 0,5 g BSA (0,1%)
0,5 ml Natriumazid-Lösung (10%)
PBS/ 1mM EDTA 500 ml 1x PBS
1 mM EDTA
RPMI/2% FCS 500 ml 1x RPMI Medium 1640
(500 ml) 10 ml FCS
5 ml Penicillin/Streptomycin
Tail buffer 50 mM TRIS pH=8
(100 ml) 100 mM EDTA
100 mM NaCl
1% SDS
63 ml Ampuwa
172.1.4 Antikörper für die Durchflusszytometrie
Tabelle 2: Verwendete Antikörper für die Durchflusszytometrie
Gelistet sind die verwendeten Antikörper für die Durchflusszytometrie mit ihren konjugierten Fluoreszenzmarkern.
APC = Allophycocyanin, APC-Cy7 = Allophycocyanin+Cyanin 7, CD = Cluster of Differentiation,
F4/80 = EGF-like module-containing mucin-like hormone receptor-like 1, FITC = Fluorescein isothiocyanate,
Ig = Immunglobulin, Ly6C = Lymphozytenantigen 6C, Ly6G = Lymphozytenantigen 6 complex locus G6D,
MHC II = Haupthistokompatibilitätskomplex-II, PE = Phycoerythrin, PE-Cy5.5 = Phycoerythrin-Cyanin 5.5,
PE-Cy7 = Phycoerythrin+Cyanin 7, PerCP = Peridinin chlorophyll, PerCP/Cy 5.5 = Peridinin chlorophyll/Cyanin 5.5.
Antigen Klon Konjugation Firma
CD3e 145-2C11 eFlour450 eBioscience, USA
CD4 RM4-5 PerCP BD Pharmingen,
USA
CD8a 53-6.7 PE BioLegend, USA
CD11b M1/70 eFlour450 eBioscience, USA
CD11c HL3 APC BD Pharmingen,
USA
CD19 6D5 PE-Cy7 BioLegend, USA
CD21/CD35 7E9 Brilliant Violet 421 BioLegend, USA
(CR2/CR1)
CD23 B3B4 PE BD Pharmingen,
USA
CD45R (B220) RA3-6B2 PE-Cy5.5 eBioscience, USA
CD45R/B220 RA3-6B2 PE-Cy7 BioLegend, USA
CD45 30-F11 APC-Cy7 BD Pharmingen,
USA
CD93 (AA4.1, AA4.1 PE-Cy7 BioLegend, USA
early B lineage)
F4/80 BM8 PE-Cy7 BioLegend, USA
IgD 11-26c FITC eBioscience, USA
IgM II/41 APC eBioscience, USA
Ly6C PE Miltenyi Biotech,
Deutschland
Ly6G 1A8 PE-Cy5.5 BioLegend, USA
Ly6G (Gr-1) RB6-8C5 PE-Cy5.5 eBioscience, USA
MHC II (I-A/I-E) M5/114.15.2 FITC eBioscience, USA
Ter119 PE Miltenyi Biotech,
Deutschland
182.1.5 Primer für die Polymerase-Kettenreaktion
Tabelle 3: Primer für die Polymerase-Kettenreaktion
Gelistet sind die verwendeten Primer für die Polymerase-Kettenreaktion(PCR)-Untersuchung sowie ihre Sequenzen.
ko = Knock-out, wt = Wildtyp
Primer Sequenz
oIMR0367 cag aga agg ttc cat gtc gg
oIMR0368 gcc att agc cat gta ctc gg
oIMR0204 cac ggg tag cca acg cta tgt c
AZ 107 att agg acc tga tgg gtg cag ctt
AZ 138 gat gga gcc cag tgt gtc tgt ata
AZ 140 gtc ctc gtc ttc ctc tgc gg
AZ 120 aag cgc ctc ccc tac ccg gt
Vav3 wt N2 atg gag ccg tgg aag cag tgc gct
Vav3 wt N4 gag aaa ctg gga cat ctg ggg cct cag
Vav3 ko 341 cag gta gcc gga tca agc gta tgc
Vav3 ko 347 gca ctc gct gct gca gcg gc
2.1.6 Verbrauchsmaterialien
Folgende Verbrauchsmaterialien wurden für die Experimente verwendet:
❖ 1,5 ml Reagenzgefäße, Fa. Sarstedt, Deutschland
❖ 1,5 ml Microtubes steril, Fa. Brand, Deutschland
❖ 2 ml Reagenzgefäße, Fa. Sarstedt, Deutschland
❖ 5 ml Rundboden-Röhrchen mit Zellsieb, Fa. BD, USA
❖ 15 ml Conical Tube, Fa. Corning, USA
❖ 50 ml Conical Tube, Fa. Corning, USA
❖ Cell Strainer Nylon Filter (40 µm Porengröße),
Fa. ThermoFisher Scientific, USA
❖ Combitips advanced ®, Fa. Eppendorf, Deutschland
❖ Deckgläser, Fa. ThermoFisher Scientific, USA
❖ EDTA Reagenzgefäße, Fa. Kabe Labortechnik, Deutschland
❖ Einbettkassetten, Fa. Simport Ltd., Kanada
❖ Einmalhandschuhe, Fa. Ansell, Deutschland
❖ FACS-Tubes, Fa. Beckman Coulter, Deutschland
❖ Gewebekulturplatte (6-Well), Fa. ThermoFisher Scientific, USA
19❖ LightCycler Multiwell Platte, Fa. Roche, Deutschland
❖ Objektträger, Fa. ThermoFisher Scientific, USA
❖ Pipettenspitzen (10 µl steril), Fa. Eppendorf, Deutschland
❖ Pipettenspitzen (20, 200, 1000 µl), Fa. Starlab, Deutschland
❖ Plastikpipetten (5, 10, 25, 50 ml), Fa. Corning Incorporated, USA
❖ Spritze (10 ml), Fa. Braun, Deutschland
❖ SubQ-Einwegspritze (∅ 0,45 mm, 26G), Fa. BD, USA
❖ Urinbecher, Fa. Sarstedt, Deutschland
2.1.7 Geräte
Folgende Geräte wurden für die Experimente verwendet:
❖ Animal Blood Counter, Fa. Scil, Deutschland
❖ Automated CellCounter TC20®, Fa. Biorad, Deutschland
❖ Biofuge pico, Fa. Heraeus, Deutschland
❖ ChemiDoc MP, Fa. Biorad, Deutschland
❖ Eismaschine, Fa. Scotsman, Deutschland
❖ FACS Galios, Fa. Bechman Coulter, Deutschland
❖ Feinwaage, Fa. Sartorius, Deutschland
❖ Kühlschrank, Fa. Liebherr, Deutschland
❖ Maus-Restrainer, Eigenanfertigung, Universität Ulm, Deutschland
❖ Megafuge 1.0R, Fa. Heraeus, Deutschland
❖ Multipipette, Fa. Eppendorf, Deutschland
❖ Paraffinausgießstation Modell TPS-1 EVO II, Fa. supplink, Deutschland
❖ PCR Maschine Labcycler, Fa. Sensoquest, Deutschland
❖ Rotary 3003, Fa. pfm medical, Deutschland
❖ Power Pack, Fa. Biometra, Deutschland
❖ Pipetten (0,1-2,5 µl, 1-10 µl, 2-20 µl, 10-100 µl, 20-200 µl, 100-1000 µl),
Fa. Eppendorf, Deutschland
❖ Tiefkühltruhe (-20 °C), Fa. Liebherr, Deutschland
❖ Trockenschrank, Fa. Heraeus, Deutschland
❖ Wasserbad, Fa. GFL, Deutschland
❖ Waterbath 1000, Fa. pfm medical, Deutschland
❖ Zeiss Observer.D1, Fa. Zeiss, Deutschland
❖ Zeiss A-Plan 5x/0,12 Ph0 M27 Objektiv, Fa. Zeiss, Deutschland
202.1.8 Datenverarbeitung
Folgende Computerprogramme wurden für die Erstellung dieser Dissertation
verwendet:
❖ Adobe Acrobat Reader DC
❖ AxioVision Rel. 4.8.1.0
❖ GraphPad Prism 7
❖ Image Lab V5.0 Build 18
❖ Kaluza Aquisation and Analysis
❖ Microsoft Excel 2013 und 2016
❖ Microsoft PowerPoint 2013 und 2016
❖ Microsoft Word 2013 und 2016
❖ RefWorks
❖ Zotero
212.2 Methoden
2.2.1 Zellzählung
Die Bestimmung der Zellzahl pro µl Blut erfolgte direkt nach der Blutentnahme
automatisch mithilfe des „Animal Blood Counters“ (ABC, Fa. Scil). Die Zellzählung
der lebenden Zellen des Knochenmarks erfolgte nach dem ersten Waschschritt
nach Gewinnung des Knochenmarks aus den Röhrenknochen (Femur) der Mäuse,
und die Zellzählung der lebenden Zellen der Milz erfolgte nach der Erythrozytenlyse,
durchgeführt durch Inkubation der Gesamtzellen der Milz in einer
0,16 M Ammoniumchloridlösung. Für die Zellzählung wurde in jedem Fall eine
1:1-Verdünnung mit Tryptanblau hergestellt und mit dem automatischen
Zellzählgerät „Automated CellCounter“ TC20® (Fa. Biorad) die Anzahl der lebenden
Zellen in einem Milliliter Zellsuspension bestimmt. Daraufhin konnte dann jeweils
die Gesamtzellzahl des entsprechenden Organs durch die Gesamtmenge der
Zellsuspension errechnet werden.
2.2.2 Blutentnahme bei Mäusen
Für die Blutentnahme wurden die Tiere zuvor für einige Minuten mithilfe einer
Wärmelampe hyperämisiert. Anschließend wurden sie mithilfe eines Restrainers in
ihrer Bewegungsfreiheit eingeschränkt. Nach dem Anritzen der Schwanzvene
mittels Skalpell wurden die Blutstropfen in einem EDTA-Röhrchen aufgefangen.
Anschließend wurde das Blut wie unter 2.2.1 beschrieben analysiert.
2.2.3 Präparation von Mäusen
Vor der Präparation wurden die MäuseTierschutzgesetz-konform in einem dafür
geeigneten Gefäß mittels Kohlenstoffdioxid getötet. Anschließend wurden sie
überstreckt, um einen Genickbruch zu erzielen. Der eingetretene Tod wurde durch
die Abwesenheit des Zwischenzehenreflexes überprüft.
Anschließend wurden die Tiere in Rückenlage fixiert und die Bauchhöhle durch
einen longitudinalen Schnitt der Sagittalebene eröffnet. Danach wurde die Milz aus
der Bauchhöhle luxiert und von den restlichen Organen getrennt. Dann erfolgte eine
22Überführung der Milz in ein 15 ml kanonisches Tube auf Eis, welches mit
5 ml RPMI/2% FCS befüllt war.
Für die Entnahme des Knochenmarks wurde ein Femurknochen vom Beckenring
abgesetzt und anschließend die Verbindung mit dem Unterschenkel durch die
Durchtrennung der genusalen Muskulatur und Bänder erreicht. Nach erfolgtem
Absetzen der Oberschenkelmuskulatur wurde der Knochen mehrfach mittels einer
SubQ-Spritze mit RPMI/2 % FCS ausgespült. Als Auffanggefäß diente ebenfalls ein
15 ml kanonisches Tube auf Eis.
2.2.4 Zellaufbereitung aus dem Blut
Nach der unter 2.2.1 beschriebenen Zählung der Zellen per µl Blut werden, je
Färbungsansatz für die Durchflusszytometrie, in ein 1,5 ml-Reagenzgefäß
75 µl PBS/1mM EDTA vorgelegt und anschließend 25 µl zu untersuchendes Blut
hinzugegeben. Zur Unterbindung unspezifischer Antikörperbindung wurde die
Zellsuspension mit muriner IgG-Lösung (0,2 µl murines IgG und 9,8 µl FACS-Puffer
je Probe → entspricht einer Konzentration von 0,1 µg murinem IgG/Probe) für 5 min
auf Eis inkubiert. Anschließend wird die Probe mit 20 µl des entsprechenden
Antikörpermixes für 20 min im Dunkeln bei 4 °C gefärbt. Das entsprechende
Färbeschema kann der Tabelle 1 entnommen werden. Danach werden die roten
Blutzellen der Probe durch Zugabe von 500 µl „FACS lysing solution“ (BD) für
10 min bei Raumtemperatur im Dunklen lysiert. Nach Zentrifugation der Probe für
20 sec bei 13.000 rpm (Runden pro Minute) wird diese noch zweimal mit jeweils
500 µl PBS/1mM EDTA gewaschen und in insgesamt 300 µl PBS/1mM EDTA
resuspendiert und in das entsprechende FACS-Tube zur Messung überführt.
23Tabelle 4: Farbschemata der Antikörpermixe zur Analyse der Blutzellen mittels Durchflusszytometrie
Der angegebene Kanal bezieht sich auf die Messeinstellungen des Gerätes FACS-Galios der Firma Beckman Coulter.
APC = Allophycocyanin, APC-Cy7 = Allophycocyanin+Cyanin 7, CD = Cluster of Differentiation, eF450 = eFlour 450,
F4/80 = EGF-like module-containing mucin-like hormone receptor-like 1, FITC = Fluorescein isothiocyanate,
Ly6C = Lymphozytenantigen 6C, Ly6G = Lymphozytenantigen 6 complex locus G6D,
MHC II = Haupthistokompatibilitätskomplex-II, PE = Phycoerythrin, PE-Cy7 = Phycoerythrin+Cyanin 7,
PerCP = Peridinin chlorophyll, PerCP/Cy 5.5 = Peridinin chlorophyll/Cyanin 5.5.
Population Antikörper Kanal µl/Ansatz
Lymphoide IgD-FITC FL1 0,5
Zellreihe CD8a-PE FL2 0,5
CD4-PerCP FL4 0,5
CD19-PE-Cy7 FL5 0,2
IgM-APC FL6 0,5
CD45-APC-Cy7 FL8 0,3
CD3e-eF450 FL9 1
FACS-Puffer 16,5
Myeloide MHCII-FITC FL1 0,1
Zellreihe Ly6C-PE FL2 1
Ly6G- FL4 0,2
PerCP/Cy5.5
F4/80-PE-Cy7 FL5 0,5
CD11c-APC FL6 0,5
CD45-APC-Cy7 FL8 0,3
CD11b-eF450 FL9 0,5
FACS-Puffer 16,9
2.2.5 Zellaufbereitung aus der Milz
Nach der in 2.2.3 beschriebenen Entnahme der Milz wird diese über ein 40 µm
Nylonmesh vereinzelt und mit 10 ml RPMI/ 2% FCS gespült. Die so entstandene
Zellsuspension wird danach im 15 ml kanonischen Tube für 5 min bei 1200 rpm und
Raumtemperatur zentrifugiert. Anschließend wird der Überstand abgesaugt und die
Erythrozyten durch Zugabe von 5 ml 0,16M Ammoniumchloridlösung für 5 min bei
Raumtemperatur lysiert. Die Reaktion wird durch Zugabe von 10 ml RPMI/ 2% FCS
gestoppt und die Zellsuspension wie oben beschrieben zweimal gewaschen. Nach
Resuspension des Zellpellets in 5 ml RPMI/2 % FCS wird die Zellzahl der
Zellsuspension wie unter 2.2.1 beschrieben ermittelt. Jeweils 1x10 6 Zellen werden
in 1,5 ml-Reagenzgefäße je Analyse per Durchflusszytometrie gegeben,
anschließend mittels 1 ml-FACS-Puffer gewaschen und die Reagenzgefäße bei
13.000 rpm für 25 sec zentrifugiert. Nachdem der Überstand verworfen wurde, wird
die Zellsuspension mit 10 µl einer murinen IgG-Lösung (vergleiche 2.2.4) für 5 min
bei 4 °C geblockt. Anschließend werden jeweils 20 µl des entsprechenden
24Sie können auch lesen

















































