Die neutrale Sphingomyelinase und Alkohol kontrollieren das regionale Hirnvolumen: eine MRI-Studie in der weiblichen Maus - OPUS 4
←
→
Transkription von Seiteninhalten
Wenn Ihr Browser die Seite nicht korrekt rendert, bitte, lesen Sie den Inhalt der Seite unten
Aus der Psychiatrischen und Psychotherapeutischen Klinik der
Friedrich-Alexander-Universität Erlangen-Nürnberg
Direktor: Prof. Dr. med. Johannes Kornhuber
Die neutrale Sphingomyelinase und Alkohol kontrollieren das
regionale Hirnvolumen: eine MRI-Studie in der weiblichen
Maus
Der Medizinischen Fakultät
der Friedrich-Alexander-Universität
Erlangen-Nürnberg
zur Erlangung des
Doktorgrades Dr. med.
vorgelegt von
Christina Anna Carina Sauer
aus Bamberg
Als Dissertation genehmigt von der
Medizinischen Fakultät der Friedrich-Alexander-Universität
Erlangen-Nürnberg
Vorsitzender des Promotionsorgans: Prof. Dr. Markus F. Neurath
Gutachter: Prof. Dr. Christian P. Müller
Gutachter: Prof. Dr. Johannes Kornhuber
Tag der mündlichen Prüfung: 13. Juli 2021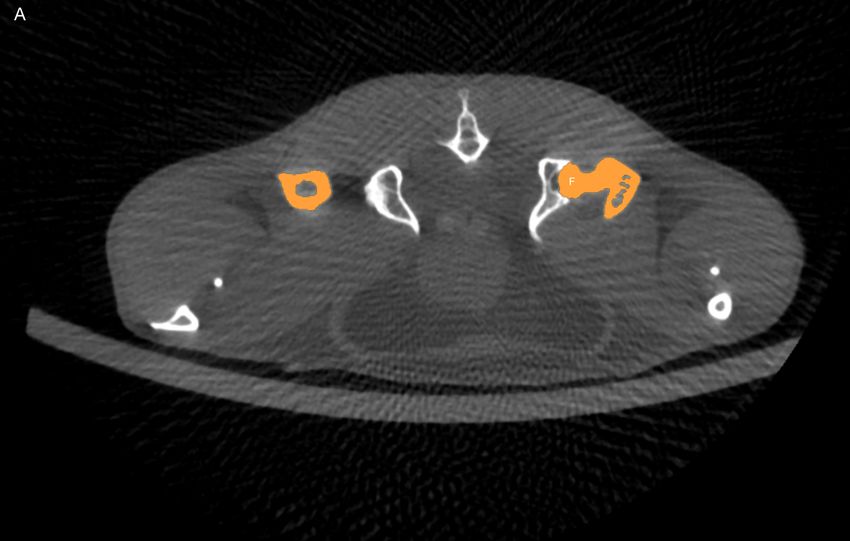
Für meinen Papa und meine ganze Familie
Inhaltsverzeichnis
Inhaltsverzeichnis i
1 Zusammenfassung 1
2 Einleitung 6
2.1 Alkoholismus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6
2.2 Rolle der Sphingolipide bei Alkoholsucht . . . . . . . . . . . . . . . . . . . . 13
2.3 Sphingolipide . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15
2.3.1 System der Sphingolipide . . . . . . . . . . . . . . . . . . . . . . . . 15
2.3.2 Entstehung von Ceramiden . . . . . . . . . . . . . . . . . . . . . . . . 17
2.3.3 Rolle der sauren Sphingomyelinase . . . . . . . . . . . . . . . . . . . 21
2.3.4 Rolle der neutralen Sphingomyelinase . . . . . . . . . . . . . . . . . . 25
2.3.5 Unterscheidung von ASM und NSM . . . . . . . . . . . . . . . . . . . 28
2.4 Ziele . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 30
3 Material und Methoden 31
3.1 Material . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31
3.1.1 Mäuse . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31
3.1.2 Alkohol . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32
3.2 Methoden . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32
3.2.1 Tiermodell . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32
3.2.2 Alkoholkonsum . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32
i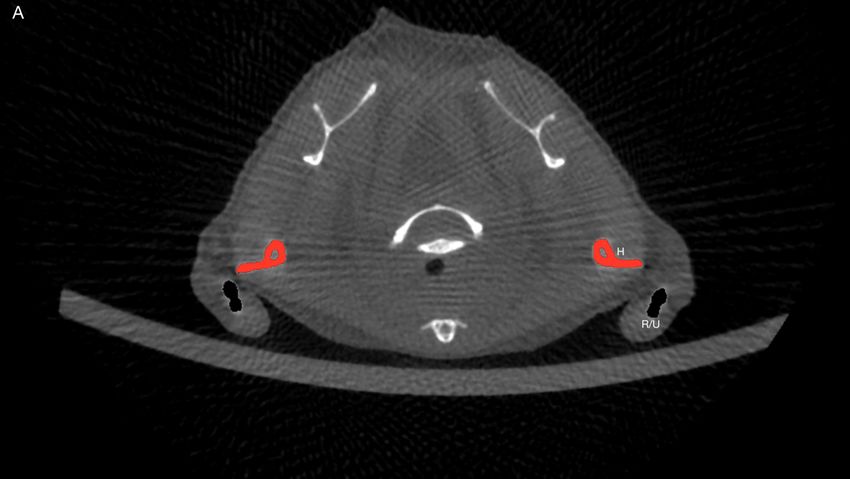
3.2.3 Magnetresonanztomographie und Computertomographie . . . . . . . . 33
3.3 Statistische Auswertung . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 35
4 Ergebnisse und Beobachtungen 42
4.1 Trinkverhalten . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 42
4.2 Hirnmorphologie . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 43
4.2.1 Dorsaler Hippocampus . . . . . . . . . . . . . . . . . . . . . . . . . . 43
4.2.2 Inselrinde . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 44
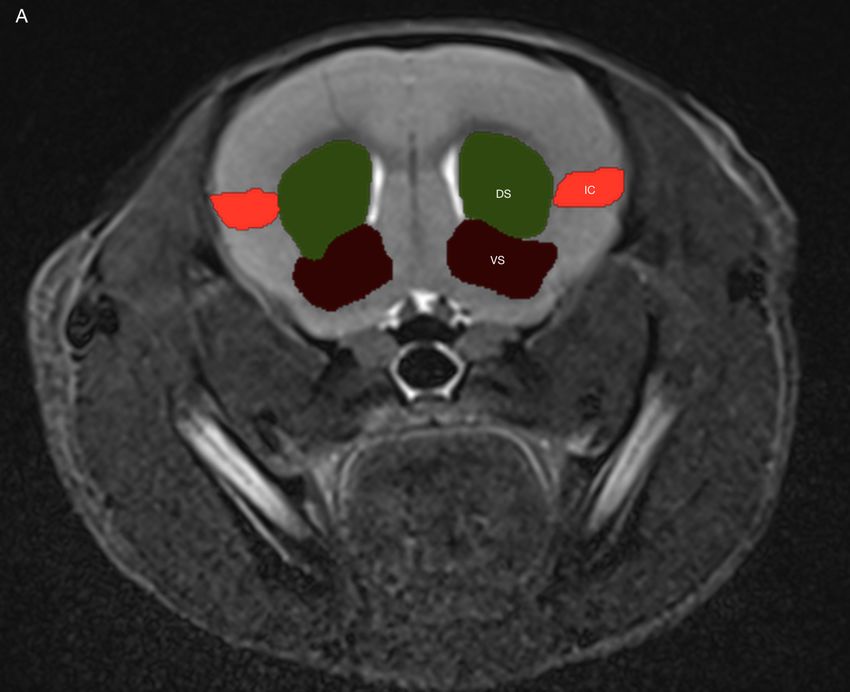
4.2.3 Dorsales Striatum . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 45
4.2.4 Ventrales Striatum . . . . . . . . . . . . . . . . . . . . . . . . . . . . 46
4.2.5 Amygdala . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 47
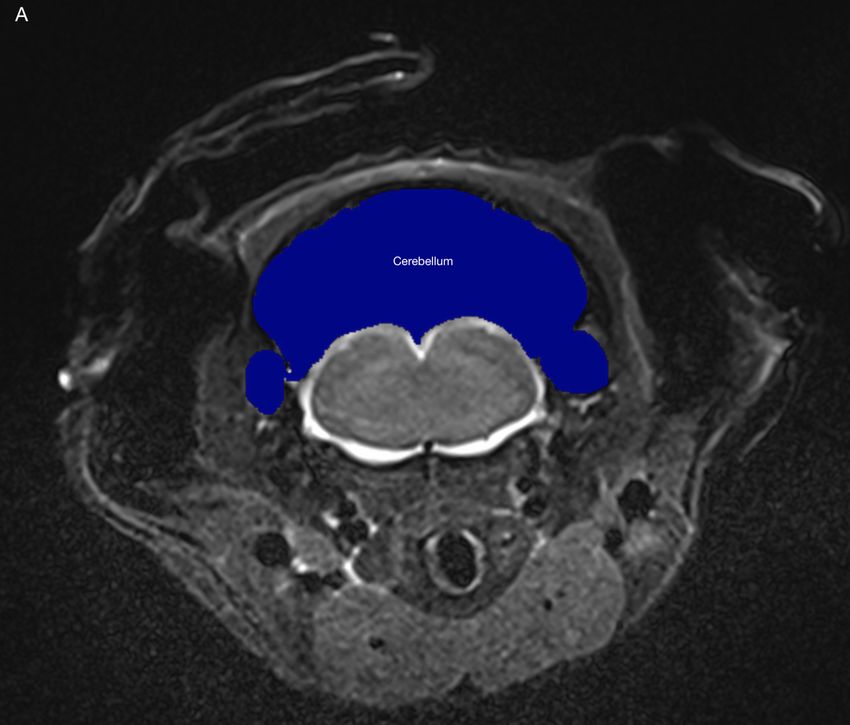
4.2.6 Cerebellum . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 47
4.2.7 Resultat . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 47
4.3 Knochenbau . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 48
4.3.1 Dichte Schädel . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 48
4.3.2 Dichte Wirbelsäule . . . . . . . . . . . . . . . . . . . . . . . . . . . . 49
4.3.3 Dichte Femur . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 50
4.3.4 Dichte Tibia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 50
4.3.5 Dichte Humerus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 51
4.3.6 Dichte Radius und Ulnar . . . . . . . . . . . . . . . . . . . . . . . . . 52
5 Diskussion 57
5.1 Zusammenfassung . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 57
5.2 Interpretation der einzelnen Ergebnisse . . . . . . . . . . . . . . . . . . . . . . 58
5.2.1 Gehirnmorphologie . . . . . . . . . . . . . . . . . . . . . . . . . . . . 58
5.2.2 Knochenbau . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 62
5.3 Limitationen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 62
5.4 Kontext der Arbeit . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 63
iiLiteraturverzeichnis 68
Abbildungsverzeichnis 79
Abkürzungsverzeichnis 80
iiiKapitel 1
Zusammenfassung
Hintergrund und Ziele
In der westlichen Welt wird Alkohol von der Mehrheit der Erwachsenen regelmäßig konsu-
miert, trotz eines nicht unerheblichen Risikos für gesundheitliche Folgen, wie psychische und
physische Schäden. Neben den medizinischen Schäden durch Alkohol, kommt es auch zu wirt-
schaftlichen Folgen und schwerwiegenden sozialen und familiären Problemen. Der volkswirt-
schaftliche Schaden durch Alkohol beläuft sich auf rund 40 Millionen Euro pro Jahr.
So konsumieren circa 6,7 Millionen Menschen alleine in Deutschland Alkohol in einer ge-
sundheitlich riskanten Form und 1,6 Millionen Menschen im Alter von 18-64 Jahren gelten
in Deutschland als alkoholabhängig. In diesem Zusammenhang ist der sehr unkritische Um-
gang und die gesellschaftliche Akzeptanz des regelmäßigen Konsums von Alkohol als äußerst
problematisch anzusehen. Diese Situation birgt eine weitere Erschwerung in der Therapie der
Erkrankung. Es gibt keine einfache spezifische Therapie, vielmehr ist die Behandlung der Alko-
holsucht sehr komplex, erfordert die volle Bereitschaft des Betroffenen und seiner Angehörigen
und dauert im Grunde ein Leben lang. Aufgrund der mangelnden Behandlungsoptionen ist die
Suche nach weiteren therapeutischen Möglichkeiten Gegenstand der aktuellen Forschung. In
den Fokus der Forschung sind hier die Sphingolipide gerückt, die im Gehirnstoffwechsel ei-
ne wichtige, aber zum Teil noch unverstandene Rolle übernehmen. Als Enzym der Familie der
Sphingomyelinasen (SMasen) ist auch die neutrale Sphingomyelinase (NSM) in den Hirnstoff-
wechsel eingebunden. Das Ziel dieser Arbeit war herauszufinden, welche Auswirkungen der
heterozygote Knockout der NSM in Kombination mit dem Konsum von Alkohol einerseits auf
die Entwicklung von Sucht und anderseits auf Morphologie von Gehirn und Knochen hat.
1Material und Methoden
Über einen Zeitraum von 43 Tagen wurde das Trinkverhalten von weiblichen fragilitas ossium
(fro)-Mäusen mit einem heterozygoten Defekt der NSM sowie wildtyp (wt)-Mäusen als Kon-
trollgruppe beobachtet. In der Versuchsgruppe wurde sowohl den fro- als auch den wt-Mäusen
der freiwillige Konsum einer 16 vol.%-Ethanollösung angeboten. Der Kontrollgruppe hingegen
stand ausschließlich Wasser zur Verfügung. Das Trinkverhalten der Mäuse wurde analysiert,
um so eine Aussage zu Konsum und Präferenz zu treffen. Zur Darstellung von Effekten des
Alkoholkonsums, der Defizienz der NSM und des Zusammenspiels beider Faktoren auf Hirn-
und Knochengewebe, schloss sich an die Phase des Trinkens bei allen 31 Mäusen eine Messung
des Gehirns mittels Magnetresonanztomographie (MRT) und eine Computertomographie (CT)-
gestützte Messung des kompletten Skeletts an.
Im Rahmen der Auswertung der Messungen mit dem Programm „aycan OSiriX mit Segmenta-
tion“ wurden die Werte für Hirnvolumina und Knochendichte der fro-Mäuse mit den jeweiligen
Kontrollgruppen verglichen.
Ergebnisse und Beobachtungen
Mäuse mit einem heterozygoten Defekt der NSM zeigten eine erhöhte Präferenz für Alkohol
und einen erhöhten Konsum von Alkohol im Vergleich zu wt-Mäusen. Der totale Flüssigkeits-
konsum und der Konsum von Leitungswasser lagen unter dem Wert der wt-Mäuse. Bei den
Auswertungen der Knochendichte durch eine CT-Untersuchung zeigten sich keine signifikanten
Unterschiede in der Knochendichte zwischen wt-Mäusen und fro-Mäusen, unabhängig davon,
ob die Mäuse Alkohol oder Leitungswasser tranken. In den Messungen der Hirnvolumina zeig-
ten sich jedoch deutliche Unterschiede zwischen fro-Mäusen, die ausschließlich Wasser tranken
und derer, die freiwillig Ethanol konsumieren konnten. Bei Ethanol-konsumierenden heterozy-
goten fro-Mäusen kam es zu einer Reduktion der Volumina in gemessenen Hirnregionen. Diese
Veränderung war vor allem in den Bereichen des dorsalen Hippocampus signifikant und im Be-
reich der Inselrinde trendsignifikant. Bei wt-Mäusen konnte diese Reduktion trotz den Konsums
von Ethanol nicht aufgezeigt werden.
Schlussfolgerung
Der alleinige Konsum von Alkohol zeigte keinen Effekt auf die Morphologie von Strukturen
im Gehirn. Obwohl Ethanol durch eine erhöhte Produktion von Ceramid eine neurotoxische
Zelldegeneration auslöst, war weder die Dauer noch die konsumierte Alkoholmenge ausrei-
chend, um in diesem Setting einen Effekt zu bewirken. In Interaktion mit einem heterozygoten
2Knockout der NSM kam es jedoch bei Konsum von Ethanol zu signifikanten Auswirkungen.
Interessanterweise war das Volumen des dorsalen Hippocampus zu Beginn des Experimentes
im Vergleich zur wt-Gruppe signifikant erhöht. Somit kann angenommen werden, dass die fro-
Mäuse vor Beginn des Alkoholkonsums über eine erhöhte Resilienz gegen Stressoren verfügten.
Trotz der eigentlich verminderten Produktion von Ceramid durch die Defizienz der NSM, traten
bei gleichzeitigem Konsum von Ethanol neurotoxische Effekte und eine Verminderung defi-
nierter Strukturen auf. Als signifikant ist diese Reduktion im dorsalen Hippocampus und als
trendsignifikant in der Inselrinde zu werten. Dabei handelt es sich um Strukturen, die immense
Wichtigkeit in der Pathogenese psychiatrischer Erkrankungen, wie Sucht und Depression, auf-
weisen. Ebenfalls konnte in NSM-Knockout-Mäusen eine erhöhte Präferenz und ein erhöhter
Konsum von Ethanol per se festgestellt werden. Ein Einfluss auf die Mineralisierung und Dich-
te der Knochen bei fro-Mäusen durch die Mutation selbst, den Konsum von Ethanol oder das
Zusammenwirken beider Faktoren war in diesem Experiment nicht festzustellen. Die Ergebnis-
se belegen spezifische Auswirkungen durch das Zusammenspiel der heterozygoten Defizienz
der NSM mit Ethanol auf die Entwicklung und Morphologie von Hirnstrukturen.
3Abstract
Background and objectives
The majority of adults in the Western world consume alcohol frequently despite the many known
negative consequences resulting therefrom, including public health, public safety and general
social welfare. Additionally, alcohol is a known factor in contributing to many negative econo-
mic issues, with an even larger overall societal impact. As an example, the cost to Germany’s
Gross Domestic Product resulting from alcohol abuse amounts to 40 million Euros per year.
In Germany alone, approximately 6.7 million people consume harmful amounts of alcohol on
a casual basis and around 1.6 Million people aged between 18 and 64 are alcohol dependent.
Wide spread social acceptance of alcohol consumption further hinders medical treatment of al-
cohol dependence. There is no easy, specific treatment for alcohol disorders; indeed, there are
a variety of treatment options, however, all require that patient’s social environment’s full ac-
ceptance and support; not to mention that alcohol treatment may be required for life. Due to the
lack of sufficient treatment options, researchers are focusing on new biochemical pathways for
therapeutic advances, including the biochemistry of sphingolipids. Sphingolipids are involved
in fundamental processes of brain metabolism, yet not all of them are fully understood. As a part
of the family of sphingomyelinases (SMases), the neutral sphingomyelinase (NSM) became the
focus of research. Although NSM has the highest concentration of SMases in the human brain,
only little about its functioning is known to research so far. In this paper we discuss NSM’s
impact on the development of alcohol dependence and its impingement on brain and bone mor-
phology in combination with the consumption of alcohol.
Material and methods
For a period of 43 days we observed the drinking behavior of two groups of female mice. Group
1. fragilitas ossium (fro)-mice suffering from a heterozygous defect of neutral sphingomyelinase
gene (fro/wt); and group 2. wildtyp (wt)-mice as a control group. In the experimental group
16 vol.%- ethanol solution was offered to fro- and wt-mice in a free-choice drinking manner.
4Whereas, the control group was offered only water. The mice drinking behavior was analyzed
to determine consumption and preference. Magnetic resonance imaging (MRI) and computed
tomography (CT) scans were performed on all 31 mice following the drinking phase to show
the effects of alcohol consumption, the impact of NSM, and the interaction of both on the mice
brains and bones. The differences in volume of mice brains in MRI-scans and bone density in
CT-scans could be determined and compared using of „aycan OSiriX Segmentation“ software.
Results and observations
Compared to wt-mice, NSM knockout mice showed a higher preference and a higher consumpti-
on of alcohol but smaller numbers in total intake and water intake. When analyzing bone density
with CT-scans, no significant difference was found in numbers between fro- and wt-mice. This
does not depend on whether mice consumed only water or had the chance to choose ethanol.
In contrast, the evaluation of measured brain structures showed significant differences between
fro-mice drinking water or consuming ethanol. A reduction of volume in specific brain struc-
tures could be observed especially in the dorsal hippocampus and individually in insular cortex.
This change could not be found in wt-mice consuming ethanol.
Conclusion
Alcohol had no effect on brain morphology. While, ethanol leads to neurotoxic cell degenerati-
on resulting from higher ceramide production in the brain. Factors such as time of exposure or
alcohol quantity was not sufficiently significant to alter this experiment’s outcome. Only the in-
teraction of ethanol consumption and the heterozygous deficiency of NSM were of significance.
The most noteworthy observation made was that the volume of the brain’s dorsal hippocampus
was higher at the experiment’s outset when compared to the wt-group leading to the conclusion
that fro-mice were more resilient towards stressors before they had access to ethanol. Also, neu-
rotoxic cell degeneration resulting from higher ceramide production and a reduction of specific
brain structures has a causal connection to consuming ethanol. Finally, the resulting volume re-
duction of dorsal hippocampus and individually in insular cortex should be rated as significant
as these structures are immensely important in the pathogenesis of psychiatric diseases, inclu-
ding, dependence issues and depression. NSM-knockout mice showed a higher preference and
consumption of ethanol per se. No impact could be observed on bone mineralization or density
in fro-mice caused by the mutation. No observations were made with respect to the consumption
of ethanol or the interaction of both. These results substantiate specific effects on the develo-
pment of brain morphology through the interaction of a heterozygous deficiency of NSM and
ethanol.
5Kapitel 2
Einleitung
2.1 Alkoholismus
Alkohol gilt als eine massive psychoaktive Droge in der westlichen Welt. (55) Die Mehrheit der
erwachsenen Bevölkerung konsumiert regelmäßig Alkohol trotz der erheblichen gesundheitli-
chen Risiken durch Konsum und Abusus. (154) Diese Tatsache ist auch auf die Entwicklung
unserer Kulturen und Umwelt zurückzuführen. (101) In nahezu jeder Familie der westlichen
Gesellschaft gibt es eine Person, die, direkt oder indirekt, unter den Folgen von Alkohol lei-
det. Schätzungsweise ist Alkohol weltweit für 3.2% aller Todesfälle verantwortlich. (154) In
Deutschland konsumieren circa. 6,7 Millionen Menschen zwischen 18 und 64 Alkohol in einer
gesundheitlich riskanten Form und circa 1,6 Millionen Menschen können als alkoholabhängig
bezeichnet werden. (15) Charakterisiert wird die Alkoholabhängigkeit durch den zwanghaften
Konsum großer Mengen Alkohol und den Verlust der Kontrolle über das Trinkverhalten. Erkran-
kungen durch Störungen des Alkoholkonsums gelten als eine der häufigsten psychiatrischen Er-
krankungen weltweit. Vor allem in Ländern mit hohem und mittelhohem Einkommen ist dieses
Problem vorhanden. (17)
Neben den physischen und psychischen Schäden resultieren auch schwerwiegende soziale und
wirtschaftliche Folgen, die sich durch Kriminalität, verminderte wirtschaftliche Produktions-
kraft und vermehrte Kosten im Gesundheitssystem äußern. Dies bedeutet im Gegenzug, dass
Aufwendungen in Höhe von mehr als 10% des Bruttoinlandsproduktes einer Industrienation
im Zusammenhang mit Alkoholkonsum und -abusus ausgegeben werden. (128) Konkret heißt
das Ausgaben in Höhe von rund 20 Milliarden (Mrd.) Euro pro Jahr alleine in Deutschland.
Die Höhe dieser Summe setzt sich einerseits aus den direkten Kosten für die Behandlung der
Alkoholerkrankung und derer Folgeerkrankungen, Entziehungskuren und stationären Aufent-
halten zusammen. Andererseits auch aus den indirekten Kosten durch Arbeitsausfälle und einer
6verminderten wirtschaftlichen Produktionskraft. (11)
Bei 10-20% der Konsumenten kommt es zu einer Vielzahl von medizinischen Komplikationen
wie Organschäden und Einschränkungen des Immunsystems. Durch die Aufnahme von Alkohol
werden zwar alle Organe betroffen, die größten Schäden entstehen jedoch an Leber, Pankreas
und im Gehirn. Neben dem eigenen aktiven Konsum gilt die passive Schädigung während der
Schwangerschaft als ebenso schwerwiegendes Problem. Acht von 1000 Kindern werden mit
dem „Fetalen Alkohol Syndrom“ geboren, welches durch den mütterlichen Konsum von Al-
kohol in der Schwangerschaft in massiven erworbenen geistigen Einschränkungen resultiert.
(128) Das entspricht rund 4000 Kindern jährlich, die durch Alkohol geschädigt werden und
bei denen sich kognitive Einschränkungen, Lern- und Entwicklungsstörungen und psychische
Auffälligkeiten zeigen. (78) Bei jungen Erwachsenen sind die Folgen von Alkoholkonsum, vor
allem vom sogenannten exzessiven „Binge Drinking“, noch weitreichend ungeklärt. (128) Ver-
mutet wird jedoch, dass das jugendliche Gehirn deutlich sensitiver auf Alkohol reagiert und es
zu schwereren Schäden und höheren kognitiven Einbußen kommt. (25) (127) (44) (150) Auch
gilt zu beachten, dass der frühe Beginn des Alkoholkonsums im Jugendalter später häufiger zu
Stress-induziertem Alkoholkonsum führt (123) (43) und die Wahrscheinlichkeit eines krankhaf-
ten Umgangs mit Alkohol erhöht. (34)
Laut der „Global Burden of Disease“ Studie der Weltgesundheitsorganisation stellt Alkohol
neben Tabak, Bluthochdruck und hohen Cholesterinwerten, eine der häufigsten Ursachen für
eine Verkürzung der Lebensdauer dar. (40) Hinzu kommen auch soziale Probleme wie Gewalt
oder familiäre Probleme durch den Konsum von Alkohol. (12)
Durch den Missbrauch von Alkohol wird die Entwicklung psychiatrischer Erkrankungen ge-
fördert. Alkoholabhängigkeit oder -missbrauch fördert die Entstehung von Angststörungen und
Depressionen mit einem Risiko, das 2- bis 3-fach höher als das Risiko der Normalbevölkerung
ist. (137) Zur Linderung der Symptomatik behandeln sich Patienten mit diesen psychiatrischen
Erkrankungen häufig selbst durch den Konsum von Alkohol, was die Abhängigkeit fördert.
Häufiger führt aber der missbräuchliche Konsum von Alkohol selbst zur Entwicklung der Ab-
hängigkeit. (128)
Alkoholabhängigkeit wird nach der 10.Revision der „International Classification of Diseases
and Health Problems“ (ICD-10) diagnostiziert, „wenn ein Folgeschaden [oder] mindestens drei
von sechs Kriterien für Abhängigkeit [innerhalb] der letzten 12 Monate gleichzeitig“ vorlagen.
(7) Zu diesen Kriterien zählen die Entwicklung von Toleranz, das starke Verlangen nach Alko-
hol, Kontrollminderung, das Auftreten von Entzugserscheinungen, die Vernachlässigung ande-
rer Interessen zugunsten des Konsums und der anhaltende Konsum trotz eindeutiger körperlicher
oder psychischer Schäden. Es gibt jedoch nicht nur Merkmale für die Diagnose der Alkoholab-
hängigkeit, sondern auch der schädliche Gebrauch von Alkohol kann anhand von spezifischen
7Kriterien diagnostiziert werden. (153) Hierzu braucht es den „deutliche[n] Nachweis, dass der
Substanzgebrauch [von Alkohol] für die körperlichen oder psychischen Schäden, einschließlich
der eingeschränkten Urteilsfähigkeit oder des gestörten Verhaltens, das zu Behinderung oder zu
negativen Konsequenzen in zwischenmenschlichen Beziehungen führen kann“, verantwortlich
ist. (7) Auch hier ist die nötige Zeitspanne für eine Diagnose mit einem Monat oder wiederholt in
den letzten 12 Monaten auftretend, definiert. (153) (7) Neben der deutschen Klassifikation nach
ICD-10, gilt die „Diagnostic and Statistical Manual of Mental Disorders“ (DSM) -Klassifikation
der amerikanischen psychiatrischen Gesellschaft als weiteres diagnostischen Diagnosesystem.
In die Wertung fallen hier die folgenden Punkte:
• Konsum von Alkohol in größeren Mengen oder über einen längeren Zeitraum als geplant
• Bestehen eines fortdauernden oder erfolglosen Wunsches den Gebrauch von Alkohol zu
reduzieren oder zu kontrollieren
• Hoher Zeitaufwand für Beschaffung und Konsum oder Erholung von der Substanz
• Craving
• Wiederholter Alkoholkonsum, mit der Folge des Versagens bei unterschiedlichen wichti-
gen Verpflichtungen
• Kontinuierlicher Alkoholkonsum trotz anhaltender oder wiederkehrender gesellschaftli-
cher oder zwischenmenschlicher Probleme
• Vernachlässigung oder Einschränkung wichtiger Aktivitäten zugunsten des Konsums
• Alkoholgebrauch in Situationen, die aufgrund des Konsums eine physische Schädigung
zu Folge haben können
• Andauernder Substanzgebrauch trotz psychischer oder körperlicher Probleme
• Entwicklung von Toleranz
• Symptome durch Entzug oder die Abschwächung/Vermeidung dieser durch den fortge-
setzten Konsum von Alkohol
Mit Hilfe dieses Klassifikationssystems kann die Erkrankung in eine milde (2-3 Kriterien werden
erfüllt), moderate (4-5 Kriterien werden erfüllt) und eine schwere Störung (6 Kriterien werden
erfüllt) eingeteilt werden. (2) Selbst nach Jahren der Abstinenz können sich diese Verhaltens-
muster wieder zeigen. (128)
Wenn über die Diagnose „Alkoholabhängigkeit“ gesprochen wird, sollte man sich vor Augen
führen, dass diese Erkrankung weder ein Zeichen von Schwäche oder moralischer Verfehlung
8darstellt, sondern vielmehr durch ein komplexes Zusammenspiel von individuellen, sozialen,
kulturellen und biologischen Faktoren entsteht. Dies wird anhand von Zwillingsstudien deut-
lich. (21) Ein erhebliches Risiko für Alkoholabhängigkeit, so wird in diesen Studien ersichtlich,
ist vererbbar. (110) In weiteren Studien, die das Verhalten analysierten, wird zum Beispiel der
Wesenszug „sensation seeking“ mit einem problematischen Umgang mit Alkohol in Verbin-
dung gebracht. (80) Schwierigkeiten beim Umgang mit emotionalem Stress und schwierigen
Situationen im Leben können ebenso als Risiko gewertet werden. (152) (13) Resümierend lässt
sich sagen, dass genetische Faktoren zu 60% zu dem Risiko beitragen im späteren Leben ei-
ne alkoholbezogene Störung zu entwickeln. (121) Negativen Einfluss können auch das soziale
Umfeld, demographische Gegebenheiten und Einflüsse durch die Umwelt haben. (13) (90) (61)
Der kulturelle Einfluss sollte nicht vernachlässigt werden: In Kulturen, in denen der Konsum
von Alkohol sehr freizügig betrachtet wird, ja noch viel mehr toleriert und durch Werbung an-
getrieben wird, ist die Prävalenz von Alkoholerkrankungen höher. (95) (18) Denn hier ist „der
Alkoholkonsum [...] gesellschaftlich akzeptiert und ritualisiert.“ (7) Problematisch ist auch, dass
der Preis für die Beschaffung von Alkohol in Ländern, in denen der Konsum toleriert wird, häu-
fig deutlich niedriger ist als in Gesellschaften, die dem Konsum von Alkohol zurückhaltend
gegenüberstehen. (18)
Auch bei Heranwachsenden ist die Entwicklung eines pathologischen Trinkverhaltens multi-
faktoriell. Als ein Risiko gilt ein alkoholkranker Elternteil. Jedoch erhöht auch eine Kombina-
tion dieses genetischen Faktors mit einschneidenden Lebensereignissen wie Misshandlung im
Kindesalter, Gewalt in der Familie und geringer sozialer Unterstützung das Risiko für den eige-
nen schädlichen Missbrauch von Alkohol bzw. die Alkoholabhängigkeit. Dies ist nicht zuletzt
auf die neurobiologischen Veränderungen zurückzuführen, auf die das heranwachsende Gehirn
deutlich sensibler reagiert. (39) Nicht zuletzt kommt es durch den Konsum von Alkohol bei Ju-
gendlichen, die die Kriterien für eine alkoholbezogene Störung erfüllen, zu Veränderungen in
der Hirnmorphologie während des Wachstums. Deutlich wird dies besonders im Hippocampus.
Ursächlich für diese Verminderung des Hippocampusvolumens bei den betroffenen Jugendli-
chen und jungen Erwachsenen sind die neurotoxischen Eigenschaften des Alkohols, die sich
störend auf dessen Wachstum auswirken. Vermittelt werden diese Effekte vermutlich durch ei-
ne reduzierte Aktivität der N-Methyl-D-Aspartat (NMDA)-Rezeptoren. Da der Hippocampus
bis in das junge Erwachsenenalter hin ausreift, ist seine Empfindlichkeit für Alkohol in die-
ser Phase sehr hoch. (29) Junge Patienten, die alkoholsüchtig sind, leiden häufig noch unter
weiteren Erkrankungen. So ist der Konsum von Cannabis und Halluzinogenen in dieser Grup-
pe erhöht. Verhaltens- und Aufmerksamkeitsstörungen, Major-Depressionen und Posttraumati-
sche Belastungsstörungen traten insgesamt unter den alkoholabhängigen Jugendlichen häufiger
in Erscheinung. Vor allem die Raten von Patienten mit einer Major-Depression waren bei der
Gruppe der alkoholabhängigen Patienten erhöht. (149) (29) Bei den Patienten, die sowohl an
einer Major-Depression als auch unter Alkoholabhängigkeit litten, konnten geringere Volumina
9des Hippocampus nachgewiesen werden. Nebenbefundlich zeigten sich deutlich erhöhte Wer-
te der Corticosteroide, wenn eine Major-Depression vorlag. (29) Ein bestehendes vermindertes
Volumen des Hippocampus gilt vermutlich ebenso als Risikofaktor für die Entwicklung einer
alkoholbezogenen Störung im Erwachsenenalter. (29)
Lange bestanden zwei Theorien für die Einflussnahme von Alkohol auf neuronale Strukturen:
die Proteintheorie und die Lipidtheorie. Letztere besagt, dass Alkohol im zentralen Nervensy-
stem (ZNS) durch eine Störung der Membranlipide Einfluss auf die Neurone nimmt. Als mög-
licher Mechanismus der Einflussnahme von Membranlipiden auf Membranproteine wie Ionen-
kanäle galt eine Störung des Fließverhaltens von Membranen und einer Unordnung der Phasen-
grenzen durch Alkohol. (128) Diese Theorie konnte jedoch nicht verifiziert werden, vielmehr
konnten die besagten Veränderungen der Fließeigenschaften der Membranen bei laboratorischen
Untersuchungen ebenfalls durch das Einwirken anderer Faktoren, wie Erwärmung, hervorgeru-
fen werden. Auch wurde in den Untersuchungen gezeigt, dass durch eine Störung des Fließ-
verhaltens von Membranen nicht alle durch Neurotransmitter gesteuerten Ionenkanäle, die für
durch Alkohol hervorgerufene Verhaltensveränderungen relevant sind, von dieser Veränderung
betroffen waren. (107) (99)
Die Proteintheorie vermutet, dass Alkohol spezifisch mit Membranproteinen wie Ionenkanälen
und Rezeptoren interagiert. (128) Grundlage für diese Annahme ist eine Publikation, die erst-
mals zeigte, dass die Funktion von NMDA-Rezeptoren durch Ethanol vermindert wird und die
schädlichen Eigenschaften des Ethanols direkt über die NMDA-Rezeptoren oder indirekt über
deren neurotrophe Funktionen vermittelt werden. So kann die Intoxikation mit Ethanol zu neu-
ronalen und kognitiven Einbußen durch Inhibition der NMDA-Rezeptor-Aktivierung führen.
(84) NMDA-Rezeptoren werden wie Kainsäurerezeptoren und α-Amino-3-Hydroxy-5-Methyl-
4-Isoxazolpropionsäure (AMPA)-Rezeptoren durch Glutamat, einem bedeutenden erregenden
Neurotransmitter im ZNS, aktiviert. (6) (84) Neben NMDA-, AMPA- und Kainsäurerezeptoren
wirkt Ethanol auch auf spannungsabhängige Calciumkanäle hemmend. (99) (149)
Neben NMDA-Rezeptoren wurden weitere Rezeptoren und Ionenkanäle identifiziert, die Bin-
dungsstellen für Alkohol aufweisen. So erzeugt Ethanol eine Funktionssteigerung des γ-amino-
buttersäure A (GABAA )-Rezeptors. GABAA -Rezeptoren sind inhibitorische Neurotransmitter-
Rezeptoren, bestehend aus einem Chloridkanalkomplex und zusammengesetzt aus einer Viel-
zahl von Untereinheiten. (5) Daneben sind auch inhibitorische Glycin-Rezeptoren sensibel für
den Einfluss von Ethanol. (93) Neben einer Wirkungsverstärkung durch Interaktion mit diesen
genannten inhibitorischen Neurotransmittern verstärkt Alkohol auch den Effekt an 5-Hydroxy-
tryptamine-3 (5-HT3 )-Rezeptoren und neuronale nicotinerge Acetylcholinrezeptoren. (99) (90)
Letztlich kommt es nach den primären Effekten von Alkohol auf diese Rezeptoren und Kanäle
auch zu indirekten Effekten auf Monoamine, Opioide und Endocannabinoide. Dieser Vorgang
ist essentiell für die Etablierung des Trinkverhaltens durch Verstärkung und Belohnung. (128)
10Durch Alkohol und auch durch andere Suchtmittel findet besonders im Nucleus accumbens und
zu einem geringeren Anteil auch im Nucleus caudatus eine verstärkte dopaminerge Stimula-
tion statt. Die erhöhte Konzentration an Dopamin im synaptischen Spalt führt schließlich zur
Stimulation des Verhaltens und zum fortgesetzten Konsum. (35)
Der Konsum von Alkohol über eine lange Zeit hinweg kann in drei Phasen eingeteilt werden:
In der ersten Phase tastet man sich an den Konsum von Alkohol heran, der dann in der zweiten
Phase als kontrollierter Konsum fortgesetzt wird. In der dritten Phase geht dieser Konsum in
einen unkontrollierten Konsum über. (129) Abhängig von biologischen, psychologischen und
sozialen Komponenten und Risikofaktoren kommt es zum Wandel und Durchlaufen von Phase
zu Phase: Nicht-Konsum, Konsum, starker Konsum, Abusus und Abhängigkeit. (108) In Tier-
studien, die die Entscheidung zwischen dem Konsum von gering- oder hochkonzentriertem Al-
kohol untersuchten, konnte festgestellt werden, dass die Faktoren der Dauer des Konsums und
der Gewöhnung hierbei eine große Rolle spielen und den Konsum von hochkonzentriertem Al-
kohol begünstigen. Ebenso war es möglich durch Phasen der Bereitstellung von Alkohol und
Phasen der Alkoholdeprivation die Entwicklung des Konsums zu beschreiben. Die Aufnahme
von Alkohol erfolgt dann in größeren Mengen und häufiger zu unpassenden Zeitpunkten, in
einer unkontrollierten und impulsiven Weise. (128) Die Komplexität der Entstehung durch das
Zusammenspiel von Genetik, Umwelt und sozialem Umfeld ergibt eine große klinische Hete-
rogenität bezüglich der Symptome und des Schweregrades der Alkoholabhängigkeit sowie der
individuellen Ansprache auf eine Behandlung. (128)
Allgemein lässt sich die Behandlung der Alkoholerkrankung in verschiedene Phasen einteilen:
• Kontaktphase: Dient der Feststellung der Problematik, der Krankheitseinsicht, der Moti-
vation zur Therapie
• Entgiftung-/Entzugsphase: Beendigung des Alkoholkonsums, Diagnose und Behandlung
physischer und psychischer Begleiterkrankungen
• Entwöhnungsphase: Erreichen der dauerhaften Abstinenz und Wiedererlangen der Er-
werbsfähigkeit in stationärem oder ambulantem Rahmen
• Weiterbehandlungs-/Nachsorgephase (11)
Problematisch ist, dass trotz der sehr hohen Prävalenz, die Rate an behandelten Patienten deut-
lich zu niedrig ist. (17) Als am effektivsten gelten Ansätze, die eine medikamentöse Therapie
mit einer Verhaltenstherapie kombinieren. Trotzdem ist die Aufrechterhaltung einer dauerhaf-
ten Abstinenz der Patienten eine Herausforderung. Auch hier profitieren nur 20-30% der Pati-
enten von den aktuell vorhandenen Therapiemöglichkeiten. Diese Tatsache zeigt, wie wichtig
11es ist, Biomarker zu identifizieren, um eine individuelle pharmakologische Therapie zu etablie-
ren. (94) Unter allen behandelten Patienten sprechen nur 20-30% auf Medikamente an, die den
Wunsch, Alkohol zu konsumieren, unterdrücken oder Medikamente, die Rückfälle verhindern
sollen. (128) Zu den am häufigsten verordneten Medikamenten in der Therapie der Alkoholsucht
gehört als eines der ältesten Therapieoptionen Disulfiram, welches in den Alkoholmetabolismus
eingreift und die Aldehyddehydrogenase blockiert. Bei fortgesetztem Alkoholkonsum kommt
es dann zu starken Intoxikationserscheinungen. Diese Nebenwirkungen sind auch der Grund für
den abnehmenden Einsatz im Sinne einer Aversionstherapie. Nach Disulfiram wurde im Jahr
1994 das Medikament Naltrexon zugelassen. Durch Hemmung der Opioidrezeptoren im Gehirn
vermindert Naltrexon die euphorisierende Wirkung von Alkohol. Als dritter Wirkstoff wird seit
2004 Acamprosat eingesetzt, welches Glutamatrezeptoren blockiert und so die durch Alkohol
verstärkte neurotransitorische Aktivität des Glutamats vermindert. (94)
Epidemiologische und klinische Studien konnten zwei verschiedene Arten des Alkoholikers
identifizieren. So gibt es die sogenannten „reward drinker“, für die der Konsum eine Beloh-
nung darstellt. In dieser Gruppe ist häufiger eine familiäre Vergangenheit in Zusammenhang
mit Alkoholabhängigkeit vorzufinden. Sie selbst beginnen im jungen Erwachsenenalter mit dem
krankhaften Konsum. Diese Patienten sprechen häufiger auf den Wirkmechanismus des Medi-
kamentes Naltrexon an. Die zweite Gruppe der Alkoholiker sind die „relief drinker“. Diese trin-
ken, um negative innere Angespanntheit zu lösen. Die Entwicklung hin zur Abhängigkeit findet
seinen Weg über einen anfänglich moderaten Konsum und geschieht zwischen dem 30 und 40
Lebensjahr. Diese Typen reagieren wohl sensibler auf den Wirkmechanismus des Acamprosat.
(117)
Die Problematik des unterschiedlichen Effektes der Medikamente wird vor allem bei Naltrexon
deutlich. Höchstwahrscheinlich kommt es durch genetische Unterschiede der µ-Opioid- Rezep-
toren der Patienten zu einem sehr unterschiedlichen Ansprechen auf den Wirkmechanismus die-
ses Medikamentes. So zeigen Patienten mit einer speziellen genetischen Variante ein besseres
Ansprechen auf Naltrexon. (28) Durch eine Publikation konnte zum ersten Mal eine Verbin-
dung zwischen den Genen der Patienten und der Antwortrate auf Naltrexon gezeigt werden,
was Hoffnung auf Verbesserungen in der individuellen Therapie macht. So gilt Topiramat, ein
antikonvulsives Medikament, eingesetzt in der Epilepsie- und Migränetherapie, momentan als
vielversprechend. Auch Topiramat inhibiert die Signaltransduktion von Glutamat und verbessert
die Wirkung von GABA. Der genaue Einfluss, den Topiramat auf der Trinkverhalten nimmt, ist
jedoch noch unbekannt. (94)
122.2 Rolle der Sphingolipide bei Alkoholsucht
Die pharmakologischen Effekte und die Wirkmechanismen von Alkohol im Gehirn werden im-
mer genauer erforscht und besser verstanden. Entgegen der Meinung, dass Alkohol unspezifi-
sche Rezeptoren im Gehirn aktiviert, gibt es Erkenntnisse, die nur für das Ansprechen einiger
spezifischer Ziel-Rezeptoren argumentieren. Als diese Rezeptoren konnten NMDA-, GABAA -,
Glycine-, 5-HT3 -, neuronale nikotinerge Acetylcholinrezeptoren, L-Typ Calciumkanäle und G-
Protein-abhängige Kalium-Kanäle identifiziert werden. (147) Durch die spezifische Aktivierung
dieser Rezeptoren einerseits und indirekte Effekte durch die Einflüsse auf Neurotransmitter und
Neuropeptide andererseits kommt es zu typischen Veränderungen durch Alkoholkonsum: von
Enthemmung über Sedierung bis hin zu Hypnose, je nach der Konzentration des Alkohols. (147)
Die verschiedenen Signalwege, die Alkohol oder seine Metaboliten aktivieren, führen sowohl zu
kurzfristigen Veränderungen von zellulären Prozessen als auch langfristig zu einer Veränderung
der Genexpression. (36) Als Konsequenz eines wiederholten Konsums von Alkohol treten mit
der Zeit zelluläre und neurophysiologische Veränderungen auf, die durch Triggerung im Verstär-
kungssystem des Gehirns das Verlangen nach Alkohol forcieren. Ausschlaggebend, ob sich eine
Abhängigkeit entwickelt, sind allerdings auch die genetische Konstellation des Individuums und
einige Umweltfaktoren. (128)
Nachweislich nimmt Ethanol auch Einfluss auf den Sphingolipidstoffwechsel. Im Tierversuch
konnte bereits gezeigt werden, dass die saure Sphingomyelinase (ASM) zu einem Anstieg der
Ceramid-Produktion führt, wenn Tiere Alkohol konsumieren. (136) (81) Es wurde sowohl in
Astrozyten (105) als auch in Hepatozyten (82) (31) nachgewiesen, dass Ethanol zum Zelltod
führt. In Astrozyten führt Ethanol durch Stimulation der ASM und der neutralen Sphingomyeli-
nase (NSM) und dem darauffolgenden Anstieg der Ceramidproduktion zu einer massiven Schä-
digung und folglich zum Zelltod. (105) Diese Rolle der ASM und NSM war ebenso als Ursache
des Ethanol-induzierten Zelltods in Hepatozyten reproduzierbar. (82) Auch hier kam es zur Ak-
tivierung der Enzyme mit einem Anstieg der Ceramid-Konzentration der Hepatozyten. (31) (81)
Bei der Alkoholabhängigkeitserkrankung des Menschen waren diese Ergebnisse für die Rolle
der ASM und NSM reproduzierbar. Für diese Untersuchungen wurde einer Versuchsgruppe im
Laufe ihres Alkoholentzuges wiederholt Blut entnommen und die Enzymaktivität der lysoso-
malen sauren Sphingomyelinase (L-ASM) bestimmt. In peripheren mononukleären Blutzellen
bei Patienten mit vorliegender akuter Alkoholintoxikation wurde hierbei eine erhöhte Aktivität
der L-ASM nachgewiesen. Verglichen wurden die Werte dieser Patienten mit akuter Intoxikation
mit den Werten von Patienten, die chronisch abhängig von Alkohol waren und sich in einem frü-
hen Stadium der Abstinenz befanden. Die ASM-Spiegel in der Kontrollgruppe waren anfänglich
deutlich niedriger. Die initial erhöhten Spiegel der ASM im Rahmen der Intoxikation nahmen
innerhalb von 24 Stunden im Laufe der Ausnüchterung ab. Am Ende des Entzugs zeigten sich
13die Werte der ASM in beiden Gruppen nochmals erhöht. (115) Vergleicht man die sekretorische
saure Sphingomyelinase (S-ASM) im Plasma chronisch alkoholabhängiger Patienten mit ge-
sunden Probanden, zeigen diese eine Erhöhung der S-ASM um das 3-fache, verglichen mit der
ausgewählten Kontrollgruppe. Diese erhöhte Aktivität sank im Rahmen des Entzugs innerhalb
einer Woche und glich sich den Werten der Kontrollgruppe an. Interessanterweise korreliert die
Aktivität der S-ASM signifikant positiv mit dem Enzym „Carbohydrate-deficient Transferrin“
(CDT), einem der klinischen Marker des Alkoholmissbrauchs. (114)
Die Aktivierung der ASM, NSM und auch der alkalischen Sphingomyelinase (alkSM) durch
Alkohol und die darauffolgende Überproduktion von Ceramid bei Menschen ist vermutlich
für die schädigende Wirkung einer akuten Intoxikation verantwortlich. (115) Der Anstieg des
Ceramidspiegels beruht nachweislich auf der vermehrten Expression einer Vielzahl von Genen
und Proteinen wie Ceramidsynthase (CerS) 1, CerS5 und CerS6. Dies konnte auch in einer Er-
höhung der für die ASM und NSM kodierenden Gene nachvollzogen werden. Es kommt bei
Ethanol-induzierten toxischen Effekten auf die Leber hierfür zu einem Anstieg der Gene Sphin-
gomyelin phoshodiesterase 1 (Smpd1) und 3 (Smpd3). Neben einer vermehrten Anhäufung von
Ceramid kommt es Ethanol-induziert ebenso durch eine verminderte Expression der Cerami-
dase 2 (CerD2) zu einem geringeren Abbau von Ceramid. (68) All diese Mechanismen haben
eine Akkumulation von Ceramid in den entsprechenden Zellen zur Folge. Chronischer Alko-
holmissbrauch führt zu einer kompletten Dysregulation des Sphingomyelinstoffwechsels, was
wiederum auch die Nebenwirkungen während des Entzuges erklären kann. (115) (114) Diese
Erkenntnisse bestätigen die vorangegangenen in vivo- und in vitro Ergebnisse, die bereits eine
gesteigerte Aktivität des Sphingomyelinstoffwechsels durch den Konsum von Ethanol vermuten
ließen. (114)
Grundsätzlich ist bekannt, dass Alkohol durch eine erhöhte Produktion von Ceramid, deren Ak-
kumulation und der Aktivierung von Enzymen des Ceramidmetabolismus zu einer Vielzahl de-
generativer Vorgänge in Gehirn, Leber, Pankreas und weiteren Organen, auch beim Ungebore-
nen, führt. (30) (148) Welcher exakte Signalweg der Ceramid-Biosynthese durch die Wirkung
von Alkohol aktiviert wird, ist noch nicht abschließend geklärt. Untersuchungen der Leber las-
sen jedoch vermuten, dass der De-novo Signalweg der Ceramidherstellung für die alkoholindu-
zierte Zelldegeneration verantwortlich ist. (23) Diese Zellschädigung fördert wiederum durch
die vermehrte Synthese und Akkumulation von Ceramid die Entstehung einer Insulinresistenz.
Ceramid kann selbst direkt die Blut-Hirn-Schranke überwinden und zu einer Degeneration der
weißen Substanz führen. Alkohol wirkt somit nicht nur direkt, sondern auch indirekt über die
Akkumulation von Ceramid, neurotoxisch. (30)
142.3 Sphingolipide
Der deutsche Arzt und Vater der Neurochemie Johann Ludwig Thudichum entdeckte bereits im
Jahr 1874 bei seinen Studien des Nervengewebes eine neue Art der Lipide, die er als Sphingoli-
pide bezeichnete. Ausschlaggebend für die Namensgebung war die Gestalt der neu entdeckten
Moleküle, die ihn an die griechische Sagenfigur der Sphinx erinnerte, wohingegen ihm die Funk-
tion der Sphingolipide ein Rätsel blieb. Auch für viele Jahre waren Sphingolipide nur in ihrer
Rolle als Membranbestandteile bekannt, was wohl auch an der Schwierigkeit lag, amphiphile
Moleküle genau zu untersuchen. Heute ist man sich jedoch darüber im Klaren, dass Sphingo-
lipide, neben ihrer Aufgabe als Bestandteile der Zellmembran, auch viele wichtige Aufgaben
als Mediatoren in fast allen zellulären Signalkaskaden erfüllen. (144) (56) (53) Diese Funktio-
nen sind dahingehend essentiell für das Überleben der Zelle, das die gezielte Abschaltung der
Sphingolipidsynthese im Genom den Zelltod zur Folge haben würde. (56) Die Komplexität der
sogenannten Lipidomik und all ihrer Funktionen übertrifft nach neuen Erkenntnissen sogar der
des Genoms oder Proteoms. Dieses sich entwickelnde Feld der Erforschung bioaktiver Sphin-
golipide wird heute immer mehr mit den Haupterkrankungen der Menschheit in Verbindung
gebracht. Die Entwicklung von Definitionen der molekularen Mechanismen und Enzymregu-
lation sowie die weitere Erforschung von Interaktionen zwischen Lipiden und Proteinen, die
Etablierung von Biomarkern und die Nutzung als Therapieansatz, sind aktuell Gegenstand der
Forschung. (53)
2.3.1 System der Sphingolipide
Für die Einordnung der Sphingolipide ist die Zugehörigkeit dieser zu der großen Familie der
Lipide relevant. Lipide können durch die Kombination mit Fettsäuren und verschiedenen wei-
teren Strukturen eine Vielzahl von Aufgaben im Organismus erfüllen. Hierzu gehören, neben
der Aufrechterhaltung der Membranstruktur und der Speicherung von Energie, auch die Signal-
transduktion oder die Regulation der Exprimierung von Genen. (68) Die höchste Konzentration
an Lipiden ist im ZNS vorzufinden. Daher basiert auch der reibungslose Ablauf essentieller
Funktionen des Gehirns auf dem funktionierenden Stoffwechsel der Lipide. (141)
Die Sphingolipide gelten als eine komplexe Familie mit verschiedenen Verbindungen (41), de-
nen als Grundstruktur der Aminoalkohol Sphingosin dient. An diese Struktur ist neben einer
Fettsäure die polare Kopfgruppe gebunden. (56) (51) Zu den Sphingolipiden gehören die Cer-
amide, die Sphingophospholipide, auch Sphingomyeline genannt, und die Glykosphingolipide,
neben weiteren Spezies wie Protein-Addukten. (68) Unterschieden werden diese drei großen
Hauptgruppen der Sphingolipide durch ihre verschiedenen Kopfgruppen. Bei Ceramiden findet
sich ein Wasserstoffatom, bei Sphingomyelinen das Phosphocholin und bei Glycosphingolipi-
15den das Saccharid. (56) (Siehe Abbildung 2.1)
Abbildung 2.1: Allgemeine chemische Struktur der Sphingolipide nach (42). Die Grundstruktur bildet meist der
Aminoalkohol Sphingosin, bestehend aus 18 Kohlenstoffatomen. Über eine Amidbindung ist Sphingosin mit einer
Fettsäure verbunden. An der polaren Kopfgruppe (dargestellt durch R = Rest) können verschiedene Reste gebunden
sein und so die drei großen Hauptgruppen der Sphingolipide unterscheiden: bei Ceramiden entspricht R einem
Wasserstoffatom, bei Sphingomyelin entspricht R dem Phosphocholin und bei Glycosphingolipiden entspricht R
einem Saccharid.
Sphingolipide besitzen eine Vielzahl von Funktionen. Sie zeigen sich in allen eukaryotischen
Zellmembranen, wie auch im Gehirn, und stellen mit etwa 10-20% einen kleinen, aber lebens-
wichtigen Anteil der Membranlipide dar. (59) Die häufigste Gruppe an Lipiden in der grauen
Substanz und in den Neuronen sind die Ganglioside, die wiederum zur Gruppe der Glykosphin-
golipide gehören. (68)
In den Membranen von Zellen bilden Sphingolipide wichtige Strukturbausteine des äußeren
Blattes ab (51) (116), können zwischen intrazellulären Organellen transportiert werden (145)
oder dienen als Bestandteil der Lipoproteine (126). Neben der Funktion der Sphingolipide als
Membranbestandteil erfüllen diese auch wichtige Aufgaben in der Signalübermittlung, im Zell-
wachstum und in der -differenzierung, bei Entzündungsreaktionen durch die Produktion von
Eicosanoiden und bei der Apoptose. (19) (27) (63) (103) (109)
Den Sphingolipiden kommt eine wichtige Rolle in der Tumorentstehung und -unterdrückung
zu. So werden Sphingosin und Ceramid als Lipide der Tumorsuppression angesehen und dienen
durch die Modulation von Proteinkinasen als physiologische Kontrollinstanz des Zellzyklus,
des Zellarrest und der Apoptose. Phosphoryliertes Sphingosin, Sphingosin-1-Phosphat (S1P),
agiert in entgegengesetzter Weise als ein Induktor der Zellproliferation, des Zellwachstums, des
Zellüberlebens und der Zellmigration. Ebenso fördert es das Überleben der Zellen im Sinne
einer Apoptoseresistenz. S1P dient der Angiogenese, der Vaskulogenese, der Neuritogenese und
den Funktionen des Immunsystems. (104) (139) (132) Zu den weiteren Funktionen des S1P
zählen sowohl die Interaktion mit G-Protein abhängigen Rezeptoren (46) als auch unabhängigen
16Rezeptoren. (132)
Das Lipidgleichgewicht des Gehirns unterliegt Einflussfaktoren wie Alter, Geschlecht, Umwelt-
faktoren, physischer Aktivität, Erkrankungen oder auch der Ernährung. Die Zusammensetzung
der Lipide des Gehirns ist folglich nicht statisch, vielmehr passt sie sich ständig an. Diese Dy-
namik erlaubt den Lipiden Einfluss auf die Signaltransduktion zu nehmen und auch selbst als
Neurotransmitter zu fungieren. Studienergebnisse der vergangenen Jahre bekräftigen die Hy-
pothese der Rolle von Membran- und Signallipiden in psychischen und neurologischen Vor-
gängen und somit auch in der Pathogenese von psychiatrischen Erkrankungen wie Depressi-
on, bipolaren Störungen, Schizophrenie und Substanzabhängigkeit. Durch Substanzen, die die
Blut-Hirnschranke überwinden, kann auf den Lipidmetabolismus des Gehirns aktiv Einfluss ge-
nommen werden. Diese Möglichkeit der Intervention kann neben Therapieansätzen auch der
Prävention psychiatrischer Krankheiten dienen. (120)
2.3.2 Entstehung von Ceramiden
Ceramid gilt als Schlüsselmolekül des Sphingolipidstoffwechsels. (Siehe Abbildung 2.2) Cera-
mid wird zum einen De-novo durch Kondensation von L-Serin und einer aktivierten Fettsäu-
re, in der Regel Palmitoyl-Coenzym A (CoA), durch das Enzym Serin-Palmitoyltransferase
(SPT) an der cytosolischen Seite des endoplasmatischen Retikulums gebildet. (56) Zum ande-
ren kann Ceramid durch eine weitere Acylierung seines Abbauproduktes Sphingosin entstehen.
(74) Ebenso wird Ceramid im Sphingomyelinzyklus durch Hydrolyse von Sphingomyelin oder
Glycosphingolipid geschaffen. Daneben kann Ceramid im Sinne eines „salvage pathways“ in
Lysosomen oder Endosomen anfallen. (142) (56) (116) (9) Der Beitrag dieser unterschiedlichen
Herstellungswege zur Summe der vorhandenen Ceramide ist nicht gleichmäßig verteilt. (75)
17Abbildung 2.2: Ceramidstoffwechsel modifiziert nach (74) (68). Die drei großen Hauptgruppen der Sphingo-
lipide werden durch Ceramid, Sphingomyelin und Glycosphingolipiden abgebildet, die zumeist aus dem Ami-
noalkohol Sphingosin und einer Fettsäure aufgebaut sind und sich durch ihre polaren Kopfgruppe unterschei-
den. Sphingolipide können zum einen De-novo durch Kondensation von L-Serin und Palmitoyl-Coenzym A
(CoA) durch Katalyse des Enzyms Serin-Palmitoyltransferase (SPT) gebildet werden. Im ersten Schritt entsteht
3-Ketodehydrosphinganin, welches unter Verbrauch von protoniertem Nicotinamid-Adenin-Dinucleotid-Phosphat
(NADPH) an der Ketogruppe durch die 3-Dehydrosphinganinreduktase (KDSR) reduziert wird und so Dihydro-
sphingosin gebildet wird. Durch Katalyse der Ceramidsynthase kommt es zur Reaktion mit Acyl-CoA und Dihy-
droceramid entsteht. In einer Flavin-Adenin-Dinukleotid (FAD)-abhängigen Reaktion wird Dihydroceramid durch
Dihydroceramid-Desaturase schließlich zu Ceramid oxidiert. Ceramid dient nun als Ausgangssubstanz für die Her-
stellung weiterer Sphingolipide, wie Sphingomyelin oder den Glycosphingolipiden. Alternativ kann Ceramid zu
Ceramid-1-Phosphat (C1P) oder Sphingosin hydrolysiert werden. Durch Phosphorylierung entsteht aus Sphingosin
Sphingosin-1-Phosphat (S1P). Da die meisten Schritte des Ceramidstoffwechsels reversibel sind, kann Ceramid
rasch De-novo oder durch Recycling wieder zur Verfügung gestellt werden. Wird S1P jedoch durch die Sphingosin-
1-Phosphat-Lyase (S1P-Lyase) degradiert, können die entstandenen Moleküle, Ethanolaminphosphat und Hexa-
decanal, nicht wieder dem Ceramidstoffwechsel zugeführt werden und stehen somit nicht mehr als Substrate für
die Ceramidherstellung zur Verfügung. (92) (56) (74) (68)
Bei der pyridoxalphosphatabhängigen Kondensation von L-Serin und Palmitoyl-CoA entsteht
unter Abspaltung von Kohlenstoffdioxid (CO2 ) durch Katalyse der SPT im ersten Schritt 3-
Ketodehydrosphinganin. Im darauffolgenden Schritt wird dieses Zwischenprodukt unter Ver-
brauch von protoniertem Nicotinamid-Adenin-Dinucleotid-Phosphat (NADPH) an der Keto-
gruppe durch die 3-Dehydrosphinganinreduktase (KDSR) reduziert und Dihydrosphingosin wird
gebildet. Dieses reagiert anschließend unter dem Einfluss von Ceramidsynthase mit Acyl-CoA
18und lässt Dihydroceramid entstehen. (92) (56) Schließlich wird Dihydroceramid in einer Flavin-
Adenin-Dinukleotid (FAD)-abhängigen Reaktion durch Dihydroceramid-Desaturase zu Cera-
mid oxidiert. (56) (68)
Weiterhin kann aus Ceramid mit Hilfe des Enzyms Ceramidase Sphingosin hergestellt werden.
Sphingosin besitzt die Eigenschaft die Proteinkinase C inhibieren zu können. Sphingosin kann
dann durch eine von zwei vorhandenen Sphingosinkinasen (SphK1 und SphK2) zu S1P phos-
phoryliert werden. (56) (19) (74) (68) Im Gegenzug kann Sphingosin bei Bedarf wieder aus
S1P mit Hilfe der spezifischen Sphingosin-1-Phosphat-Phosphatase regeneriert werden. (19)
Ceramid, Sphingosin und S1P gelten alle als bioaktive Lipide. Sie vermitteln Zellproliferation,
Differenzierung, Apoptose, Adhäsion und Migration durch die Kontrolle des Gleichgewichtes
zwischen der Hydrolyse von Ceramid und der Herstellung von Sphingosin und S1P. Das ent-
scheidende Enzym für dieses Gleichgewicht ist die Ceramidase. Im menschlichen Genom wur-
den fünf verschiedene Ceramidasen identifiziert, für die fünf verschiedene Gene codieren. Je
nach ihrem pH-Optimum können die saure Ceramidase (AC), die neutrale Ceramidase (NC) und
die alkalischen Ceramidasen (ACER1-3) unterschieden werden. Ein Zusammenhang zwischen
einem Funktionsverlust der Ceramidasen und diversen Erkrankungen konnte bereits festgestellt
werden. (85) (19) Zwei der Metaboliten des Ceramid, S1P und Ceramid-1-Phosphat (C1P), sind
untereinander konvertierbar. (19) Seit dem Jahr 2005 ist bekannt, dass C1P ebenfalls für die
Kontrolle physiologischer Abläufe auf Zellebene relevant ist und ebenfalls als bioaktives Mo-
lekül fungiert. (19) (56)
Da ein Großteil der enzymatischen Schritte des Sphingolipidzyklus reversibel sind, kann Cera-
mid neben der De-novo-Synthese auch zügig durch den Abbau von komplexen Membranlipiden
mit Hilfe von Glucosylceramidase oder durch die Wiederverwertung von Sphingosin oder S1P
im Sinne eines „salvage pathways“ generiert werden. (74) S1P kann hierbei durch Dephospho-
rylierung mit Hilfe der Sphingosin-1-Phosphat-Phosphatase zu Sphingosin abgebaut werden,
welches wiederum durch die Ceramidsynthase Ceramid bereitstellt. (68) (74) (32) Wird S1P
durch die Sphingosin-1-Phosphat-Lyase zu Ethanolaminphosphat und Hexadecanal abgebaut,
können diese Metaboliten nicht wieder in den Sphingolipidzyklus eingeschleust werden. Somit
stehen sie nicht mehr für die Ceramidherstellung zur Verfügung. (32)
Wie beschrieben kann Ceramid auch im Sphingomyelinzyklus aus Sphingomyelin hydrolysiert
werden. Hierfür bedarf es der Katalyse durch spezifische Sphingomyelinasen, auch Sphingo-
myelin Phosphodiesterasen (SMPD) genannt, die unter Abspaltung des Phosphorylcholinrestes
zur Bildung von Ceramid führen. Sphingomyelinasen sind sphingomyelinspezifische Formen
der Phophatase C (67) und als drei große Gruppen vorliegend, die je nach ihrem pH-Optimum
klassifiziert werden: die ASM, die alkSM und die NSM. (67) (32) (20) Die ASM zeigt sich vor-
rangig in den Endosomen und Lysosomen aller Zelltypen, die NSM ist in verschiedenen Zell-
kompartimenten zu finden. Ganz im Gegensatz zur alkSM, die ausschließlich in den Zellen des
19Intestinums auffindbar ist. (32) In erster Linie erfolgt dieser Abbau durch die entsprechenden
Hydrolasen in den Lysosomen. (56)
Das im endoplasmatischen Retikulum entstandene Ceramid wird anschließend durch Einschluss
in Vesikel oder mit Hilfe von einem Ceramid-Transfer-Protein (CERT) in den Golgi-Apparat
transportiert. Soll Sphingomyelin produziert werden, erfolgt der Transport von Ceramid über
das CERT1. Ist hingegen die Synthese von Glucosylceramid erforderlich, wird Ceramid über
den vesikulären Weg in den Golgi-Apparat befördert. (157) (32) (146) Hier kann nun die Syn-
these von Glucosylceramid oder Sphingomyelin stattfinden, also den Sphingolipiden, die zu
den fundamentalen Bestandteilen der Plasmamembran zählen. (74) Wenn das im Golgi-Apparat
vorhandene Ceramid nicht als Strukturbaustein der Zellen benötigt wird, kann es ebenso durch
die Ceramidkinase (CERK) zu C1P phosphoryliert werden. C1P gilt als wichtiger Aktivator der
Phospholipase A2 und ist in der Lage die Apoptose der Zellen zu hemmen. (56) (19) (68) Durch
Ceramidkinasen gebildetes C1P ist zwar bereits seit Beginn der 1990 bekannt, jedoch sind seine
weiteren Funktionen bis heute nicht komplett erforscht. Bekannt ist jedoch, dass CERK auch
große Ähnlichkeit mit SphK hat, welche wiederum die Phosphorylierung von Ceramid anregt,
um S1P zu bilden. (135)
An der luminalen Seite der Golgi-Membran ist es möglich, Ceramid durch die Glucosyl-Ceramid-
Synthase zu Glucosylceramid (GlcCer) zu glykosylieren. Dies schafft wiederum die Ausgangs-
substanz für mehr als 300 verschiedene komplexe Glykosphingolipide. (68) (32) Dazu zählen
Glucosylceramide, Galactosylceramide und Sulfatide. Im ersten Schritt der Synthese wird Urid-
indiphosphat (UDP) bei der Reaktion von Ceramid mit UDP-Galactose abgespalten. Nach Sub-
stitution mit Galactose entsteht das Sphingolipid Galactosylceramid. Wird nun hierauf ein Sul-
fatrest, meist das 3’-Phosphoadenosin-5’-phosphosulfat, an das C-Atom 3 der Galactose einge-
führt, ist die Sulfatidbiosynthese erfolgt und Sulfatid entstanden. Für die Synthese von Ganglio-
sid wird kommt es an der cytosolischen Seite des Golgi-Apparats zur Bindung eines Glucosylrest
an das Ceramid und es entsteht Glucosylceramid. Der Glucosylrest entstammt der UDP-Glucose.
Werden weitere Monosaccharidreste ergänzt ist die Biosynthese der Ganglioside komplemen-
tiert. In besonders hoher Anhäufung sind Ganglioside im Nervensystem vorzufinden. (56) Dane-
ben spielt sich an der luminalen Seite der Golgi-Membran die Synthese von Sphingomyelin ab.
Dafür kommt es zur Reaktion von Ceramid mit Cytidin-5’-diphosphocholin (CDP-Cholin) oder
Phosphatidylcholin unter Katalyse des Enzyms Sphingomyelinsynthase. Neben Sphingomyelin
entsteht Diacylglycerin. (56) (19) Nach Abschluss der Biosynthese werden Sphingomyelin und
die Glykosphingolipide wieder zur Plasmamembran zurückgeführt. Somit können sie an die-
ser Stelle ihre Aufgabe in der Formung von Mikrodomänen erfüllen. (68) Neben der De-novo-
Synthese von Sphingolipiden und dem Sphingomyelinzyklus gibt es noch einen weiteren Weg
der Ceramidgewinnung: das Recycling über Endosomen und Lysosomen durch Rezirkulation.
Dieser Weg wird durch die Proteinkinase C aktiviert. (9) Durch die Aufnahme und den Ab-
bau von Gangliosiden und der folgenden Freisetzung von Galactose, N-Acetyl-Galactosamin,
20Sie können auch lesen