Einführung in die molekulare Zellbiologie - JGW eV
←
→
Transkription von Seiteninhalten
Wenn Ihr Browser die Seite nicht korrekt rendert, bitte, lesen Sie den Inhalt der Seite unten
Einführung in die molekulare Zellbiologie
Uta Hardt, 2018
Dogma der Molekularbiologie
Desoxyribonukleinsäure (DNS, engl. DNA) trägt die genetische Information einer Zelle und
besteht aus tausenden von Genen (Bakterien/Archaeen: 1.000 - 6.000; Maus/Mensch:
20.000 - 25.000). Jedes Gen bestimmt, wie ein bestimmtes Protein (Synonym: Eiweiß)
aussehen soll. Proteine übernehmen wichtige Aufgaben für die Funktion und Struktur einer
Zelle. Somit bestimmt der genetische Informationsfluss von der DNA zum Protein die
Komposition und dabei die Funktion der Zelle. Diese Informationskette bezeichnen wir als
das Dogma (griech. Lehrsatz, Meinung) der Molekularbiologie (Abbildung 1). Die DNA der
Eukaryoten (siehe Abschnitt Was ist ein Eukaryot?) befindet sich im Zellkern und ist in
Chromosomen organisiert. Die DNA wird vor der Zellteilung verdoppelt, damit jede
Tochterzelle einen vollständigen Gensatz besitzt. Wenn ein bestimmtes Protein benötigt
wird, wird gezielt das entsprechende Gen in Ribonukleinsäure (RNS, engl. RNA)
umgeschrieben (Transkription). Die RNA wird zuerst weiterverarbeitet, indem nicht
codierende Teile entfernt werden und der entscheidende Teil gegen Abbau geschützt wird
(Prozessierung), und dann aus dem Zellkern heraus geschleust. Außerhalb des Zellkerns
werden entsprechend dem genetischen Code Proteine synthetisiert (Translation), die durch
komplexes Falten eine Struktur bekommen, die die Funktion des Proteins bestimmt.
Was ist ein Eukaryot?
Als Eukaryo(n)ten (griech. echter Kern) bezeichnet man alle Lebewesen, deren Zellen einen
Zellkern (lat. Nukleus) besitzen. Im Gegensatz zu Prokaryo(n)ten (griech. vor-Kern), die
keinen Zellkern besitzen und die Bakterien und Archaeen umfassen, sind Eukaryoten häufig
mehrzellig. Die eukaryotische Zelle ist außerdem größer und in komplexen
membranumhüllten Subkompartimenten organisiert. Diese spezialisierten Einheiten nennt
man Organellen.
Abbildung 1 Stark vereinfachte Übersicht über den Lehrsatz der Molekularbiologie. Beachte: Lehrsätze müssen nicht
zwingend der Wahrheit entsprechen!
1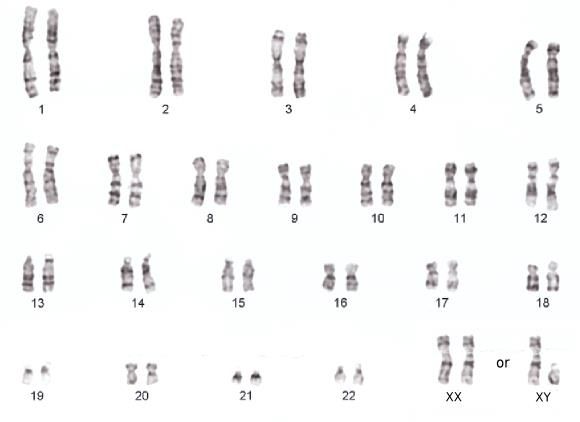
DNA
Das DNA-Gerüst formt eine Doppelhelix und besteht aus Basen und einem
Zuckerphosphatrückgrat. Die beiden Stränge der Helix sind zueinander komplementär. Das
bedeutet, dass jeder Strang als Vorlage für die Synthese des anderen Strangs herreicht. Die
strukturgebenden Bausteine der DNA sind Pentosen (Zucker mit 5 Kohlenstoffatomen),
Desoxyribosen, die über Phosphodiesterbindungen miteinander verknüpft sind. Diese
formen die zwei Stränge des Zuckerphosphatrückgrats an der Außenseite der Doppelhelix.
Jede Ribose bindet genau eine der vier Basen: Adenin (A), Guanin (G), Cytosin (C) oder
Thymin (T). Die komplementären Stränge werden durch Basenpaarung zusammengehalten:
G ist immer mit C über drei Wasserstoffbrückbindungen und A ist immer mit T über zwei
Wasserstoffbrückenbindungen gepaart (siehe Abbildung 2). Eine Base mit einem Zucker
wird als Nukleosideinheit bezeichnet, fasst man die Phosphatgruppe noch mit spricht man
von einem Nukleotid.
Abbildung 2 Die chemische Struktur der DNA. Das Rückgrat besteht aus Phosphatgruppen (violett) und Zuckern (blau), die in
sich wiederholenden Einheiten aneinandergehängt sind und deren Atombindungen eine Helixstruktur, also eine Wendelung
des Rückgrates erzeugen (siehe Illustration am linken Rand). Am Zucker hängen Basen, die entweder aus einem Ring
(Pyrimidine) oder zwei Ringen (Purine) bestehen. Über Wasserstoffbrückenbindungen (positiv polarisierter Wasserstoff wir
von negativ polarisiertem Sauerstoff angezogen) bindet immer eine kleinere Pyrimidinbase an ein größere Purinbase.
Cytosin (C, grün) paart mit Guanin (G, gelb), Thymin (T, grau) paart mit Adenin (A, gelb-orange).
Chromosomen
Denkaufgabe 1: Das Genom einer menschlichen Zelle besteht etwa aus 3,2 x 109
Nukleotiden. Der Abstand zwischen den Zentren von zwei benachbarten Nukleotiden
beträgt etwa 0,34 nm. Wir nehmen an, dass der Nukleus einer Zelle kugelförmig ist und
einen Durchmesser von 6 μm hat. Suche einen makroskopischen Vergleich für diese
mikroskopischen Größenverhältnisse. Wie würdest du den Strang im Raum organisieren?
Die Zelle löst dieses Problem mit spezialisierten Proteinen, die DNA binden können und
dabei Schleifen und Windungen bilden. Viele dieser Proteine gehören zu der Klasse der
sogenannten Histone, die in so großen Mengen vorhanden sind, dass sie der Masse der DNA
nicht nachstehen. Den Komplex aus (Histon-)Proteinen und DNA nennt man Chromatin. Ein
Oktamer (griech. acht) aus Histonen bildet das Nukleosom, auf das die DNA aufgewickelt ist.
Diese Nukleosomen bilden einen elektronenmikroskopisch sichtbaren Chromatinfaden.
Dieser Chromatinfaden ist wiederum organisiert in einer Serie von Schlaufen, die letztlich
Chromosomen (griech. Farbe, Körper) bilden (siehe Abbildung 3). Ein diploider (griech.
doppelt) menschlicher Chromosomensatz enthält im Regelfall 46 Chromosomen inklusive
2der zwei Geschlechtschromosomen (XX bei Frauen oder XY bei Männern). Die
Chromosomen werden oft als X-förmige Strukturen dargestellt. Die beiden Teile oder Arme
des Chromosoms werden als Chromatiden bezeichnet und tragen die gleiche genetische
Information. Tatsächlich existieren Chromosomen aber nur für einen sehr kurzen Moment
in dieser X-Form im Lebenszyklus einer Zelle. Der genaue Zustand der Chromosomen
während des Zellzyklus wird im Abschnitt Zellteilung beschrieben.
Abbildung 3 Illustration der Verpackung der DNA in Chromosomen. Beschreibung im Text. Quelle: Purves et al., Life: The
Science of Biology, 7th edition.
Zellteilung
Zellteilung ist eine grundlegende Eigenschaft von Zellen und ist die Voraussetzung für
Wachstum, Reproduktion (lat. Vervielfältigung) und Regeneration (lat. Erneuerung).
Zellteilung erfolgt nach einem einheitlichen Mechanismus. In tierischen Zellen können zwei
Teilungsmechanismen beobachtet werden, die Mitose (griech. Faden) und die Meiose
(griech. Verminderung), die im Folgenden näher erläutert werden.
Mitose
Mitose ist ein Teilungsmechanismus, bei dem somatische Zellen verdoppelt werden. Die
Periode von einer bis zur nächsten Zellteilung bezeichnet man als Zellzyklus. Der Zellzyklus
kann in zwei Phasen unterteilt werden, die Interphase (lat. Zwischenphase) und die Mitose.
In der Interphase nimmt das Zellvolumen zu und das genetische Material wird verdoppelt.
Die zelluläre Aktivität dieser Phase zeichnet sich durch vielfältige Syntheseprozesse aus, die
morphologisch (Morphologie, griech. Gestaltenlehre) unscheinbar sind. Im Gegensatz dazu
ist die Mitose morphologisch erkennbar. Bei der Mitose werden die Chromatiden jedes
Chromosoms unter Aufbau eines mikrotubulären Spindelapparates auf die Tochterzellen
verteilt. Die Mitose wird durch die Durchschnürung des Cytoplasmas, die Cytokinese (griech.
3Zellteilung), beendet. Die Gesamtdauer des Zellzyklus variiert sehr stark in verschiedenen
Zelltypen. Die meisten Säugerzellen teilen sich alle 12 bis 36 Stunden.
Somatische (griech. körperliche) Säugerzellen sind diploid, d.h. sie besitzen zwei
Chromosomensätze, einen paternal (lat. väterlich) und einen maternal (lat. mütterlich)
vererbten Chromosomensatz. Diese Chromosomensätze sind zueinander homolog, d.h. sie
besitzen die gleichen Loci (lat. Genorte), aber nicht zwingend die gleichen Allele (griech.
gegenseitig) eines Gens. In der Interphase besitzen Zellen einen intakten Kern, dessen
Chromosomen nicht als eigenständige Elemente erkennbar sind. Die Interphase ist in drei
Stadien eingeteilt: In der G1-Phase (gap, engl. Lücke) findet vermehrt RNA- und
Proteinbiosynthese statt. Während dieser Phase liegt jedes der 46 Chromosomen als
einzelne Chromatide vor. Jede Chromatide existiert dabei zweimal, eine maternal und eine
paternal vererbte. Die DNA-Menge wird mit 2c (copies, engl. Kopien) angegeben. In der
darauffolgenden S-Phase (synthesis, engl. Synthese) wird die DNA re(du)pliziert (lat.
verdoppelt). Die DNA-Menge steigt dabei kontinuierlichen von 2c auf 4c. Am Ende der S-
Phase liegt jedes der 46 Chromosomen damit mit zwei Chromatiden vor. Die G2-Phase ist
wieder durch RNA- und Proteinbiosynthese gekennzeichnet. Die DNA-Menge von 4c bleibt
während der G2-Phase bis zur Mitose erhalten. Zellen, die nach der G1-Phase keine DNA-
Replikation und damit keine Mitose einleiten, verbleiben in der sogenannten G0-Phase.
Beispiele für Zellen im G0-Stadium sind die Lymphozyten im Blut, die sich nur
vorübergehend nicht teilen, oder Nervenzellen, die ihre Teilungsfähigkeit weitesgehend
verloren haben.
Das Mikrotubulinzytoskelett wird von den sogenannten Centrosomen organisiert (engl.
Microtubule organizing centers, MTOCs). Der Grundbaustein der Mikrotubuli ist das Protein
Tubulin. Durch die Anlagerung von zwei verschiedenen Tubulinen enstehen Heterodimere,
die wiederum durch Aneinanderlagerung fadenförmige Aggregate, Protofilamente, bilden.
Mikrotubuli sind Röhrchen, die meistens aus 13 Protofilamenten bestehen. Diese
Mikrotubuli sind im Centrosom verankert. Das Centrosom besteht aus einem Centriol, das in
das perizentrioläre Material (PCM) eingelagert ist. Das Centriol besteht wie das Zytoskelett
aus Mikrotubuli. Die Zusammensetzung des PCM ist komplex und über den Zellzyklus
hinweg variabel. In der G1-Phase von tierischen Zellen liegen die beiden Centriolen eines
Centrosoms im rechten Winkel zueinander und in der Nähe des Zellkerns. Im weiteren
Verlauf entfernen sich die sogenannten Elterncentriolen voneinander. In der S-Phase wird
senkrecht zu den zwei Elternzentriolen kontinuierlich je eine Tochtercentriole ausgebildet,
die als Diplosomen aus je zwei Centriolen in der G2-Phase bis zur Mitose erhalten bleiben.
Der Zellzyklus wird auf komplexe Art von Proteinen reguliert. Eine zentrale Rolle wird dabei
den Cyclinen zugeschrieben, die zyklisch in ihrer Konzentration variieren. Im Zellzyklus gibt
es zwei konzeptionelle Restriktionspunkte in der G1- und der G2-Phase, wo über das
weitere Schicksal der Zelle entschieden wird. Diese Restriktionspunkte stellen sicher, dass
keine geschädigte bzw. unvollständig replizierte DNA vorliegt, bevor die S-Phase bzw. die
Mitose eingeleitet wird.
Die Mitose in höheren Eukaryoten lässt sich konzeptionell in 5 Stadien einteilen: die
Prophase (griech. vor), die Prometaphase (griech. vor/zwischen), die Metaphase (griech.
zwischen), die Anaphase (griech. auf/auseinander) und die Telophase (griech. Ziel).
4Während der Prophase kondensieren die Chromosomen, die als individuelle Einheiten
innerhalb des intakten Zellkerns vorliegen. Die Centrosomen wandern entlang der
Kernmembran bis sie an gegenüberliegenden Polen des Zellkerns zu liegen kommen, von wo
sie den Spindelapparat aus Mikrotubuli organsieren. Dabei werden die polaren Mikrotubuli
auch als Astern bezeichnet. In der Prometaphase beginnt sich die Kernmembran aufzulösen
und die Mikrotubuli strahlen in den Kerninnenraum ein. Spindelmikrotubuli weisen eine
sehr hohe dynamische Instabilität auf. Das heißt, dass das gesamte Mikrotubuligerüst des
Spindelapparates binnen 20-30s vollständig umgebaut ist (siehe Video). Diese Eigenschaft
macht die Suche nach Chromosomen zur Anheftung sehr effizient. Der Kontakt zwischen
Chromosom und Mikrotubulus wird durch Proteine im Chromatin stabilisiert. In der
Metaphase ist die Kernmembran vollständig zerfallen und man spricht von der offenen
Mitose. Die Chromosomen sind während der Metaphase in der Äquatorialebene zwischen
den Spindelpolen angeordnet. Man bezeichnet diese Anordnung, die senkrecht zur
Spindelachse liegt, auch als Metaphaseplatte. Bei genauerer Betrachtung liegen
insbesondere die Centromere der Chromosomen, die als Einschnürung erkennbar sind, in
dieser Ebene. Die Centromere sind die Chromosomensegmente, die mit dem Spindelapparat
interagieren. Sie bestehen in der Regel aus repetitiven nicht transkribierten DNA
Abschnitten. Ähnlich wie die Centrosomen sind auch die Centromere in Proteine
eingelagert. Diese sogenannten Kinetochore (griech. Bewegungsort) sind platten- oder
kugelförmig und dienen den Spindelmikrotubuli als Ansatzstellen. Die polwärtige
Ausstattung von Kinetochoren auf beiden Chromatiden eines Chromosoms wird für die
Anordnung in der Äquatorialebene zwischen den Centrosomen verantwortlich gemacht. In
der Anaphase wird jede Chromatide eines Chromosoms über den Spindelapparat polwärtig
zu den entgegengesetzten Spindelpolen mit einer Geschwindigkeit von 1 µm/min
transportiert. Die zwei wandernden Chromatidensätze werden analog zur Metaphaseplatte
auch als Anaphaseplatten bezeichnet. Außerdem vergrößert sich während der Anaphase der
Abstand zwischen den beiden Spindelpolen, der zu einer Spindelstreckung führt. Das
Spindelmikrotubulinsystem im Bereich der Äquatorialebene ist so streng angeordnet, dass
der Eindruck eine Parakristalls entsteht. Schließlich beginnt mit der Telophase der Abbau
der Kinetochormikrotubuli, es bilden sich die Kernmembranen der Tochterkerne und das
Chromatin dekondensiert.
Die Cytokinese setzt am Ende der Mitose ein. Die Teilungsebene wird durch die Lage der
Spindelpole festgelegt und liegt in der Ebene, in der auch die Metaphaseplatte entsteht.
Man vermutet, dass jede Zelle etwa gleiche Anteile der cytoplasmatischen Zellbestandteile
erhält. Zu diesen Bestandteilen gehören die Mitochondrien, der Golgi Apparat (nach dem
Entdecker Camillo Golgi) und das endoplasmatische (griech. Eigenplasma) Reticulum (lat.
Netz). Die enstehende Einstülpung der Zellmembran wird als Teilungsspalt bezeichnet. Die
Cytokinese verläuft in der Regel symmetrisch, d.h. dass beide Tochterzellen das gleiche
Volumen erhalten. Es gibt aber auch Ausnahmen, z.B. während der Embryogenese von
Invertebraten (lat. Wirbellose). Der kontraktile Ring, der zur Cytokinese führt, besteht aus
Aktinfilamenten (lat. Aktinfäden), die mit dem Motorprotein Myosin überlappen. Aktin ist
ein Protein, das ähnlich wie Tubulin fadenförmig polymerisiert. Der kontraktile Ring liegt
unmittelbar unterhalb der Zellmembran. Durch Kontraktion dieses Rings schnüren sich die
Tochterzellen voneinander ab. Allerdings kann der Ring die streng angeordneten
Mikrotubulibündel des Spindelapparates nicht zertrennen, sodass die Reste dieses
Bestandteils auf eine Tochterzelle übertragen werden.
5Abschließend lässt sich feststellen, dass die Mitose die Zahl der Chromosomen während des
Zellzyklus konstant hält.
Im obigen Text sind einige Begriffe durch Unterstreichen hervorgehoben. Beschriftet die
folgenden Abbildungen mit diesen Begriffen!
67
Meiose
Im Gegensatz zur Mitose wird die genetische Information bei der Meiose halbiert. Bei der
Meiose betrachtet man zudem nicht den Teilungsmechanismus während eines Zellzyklus,
sondern während des sexuellen Entwicklungszyklus. Dieser erstreckt sich über Generationen
beginnend mit der Befruchtung der Eizelle bis zur Produktion von eigenen Nachkommen.
Das Bedeutungsgewicht, das der sexuellen Fortpflanzung dabei zukommt, wird dadurch
verdeutlicht, dass der Fortpflanzung ein komplettes Organsystem zugeordnet ist, das für das
Individuum andernfalls völlig nutzlos ist.
In den Geschlechtsorganen entwickeln sich die Gameten (griech. Keimzellen), die ohne die
Hilfe von den umliegenden somatischen Zellen zu Grunde gingen. Gameten enstehen im
Zuge einer Meiose und tragen reduzierte Information in einem haploiden (griech. einfach)
Chromosomensatz. Durch die Befruchtung der Eizelle entsteht wieder eine Zygote (griech.
vereint) mit einem diploiden Chromosomensatz (Abbildung 4), die das Potenzial hat, einen
vollständigen Organismus zu bilden.
Denkaufgabe 2: Warum kann die asexuelle Fortpflanzung mitotisch ablaufen, wohingegen
die sexuelle Fortpflanzung meiotisch ist?
Der Meiose geht eine Interphase voraus, während der jedes Chromosom sein genetisches
Material verdoppelt. Dieser Vorgang ist der Chromosomenreplikation vor der Mitose
ähnlich. Aus jedem Chromosom gehen zwei genetisch identische Schwesterchromatiden
hervor, die durch ihre Centromere miteinander verbunden sind. Die Centrosomen
replizieren sich ebenfalls.
Die meiotische Prophase I dauert länger und ist komplexer als die Prophase der Mitose. Die
Chromosomen beginnen sich zu verdichten, und die homologen Chromosomen – jede
bestehend aus zwei Schwesterchromatiden – paaren sich. Dieser Vorgang wird Synapsis
bezeichnet. Dabei verbindet eine Proteinstruktur – der Synaptonemal-Komplex – die beiden
homologen Chromosomen entlang deren Längsachse. Wenn der Synaptonemal-Komplex in
der späten Prophase verschwindet, wird jedes Chromosomenpaar im Mikroskop als Tetrade
sichtbar, einem Komplex aus vier Chromatiden. Über das ganze Chromosom verteilt
überkreuzen sich die Chromatiden homologer Chromosomen. Diese Überkreuzungen
werden Chiasmata genannt. Die Chiasmata halten die homologen Chromosomenpaare bis
zur Anaphase I zusammen. Die Chromosomen tauschen an den Chiasmata Segmente aus.
In der Zwischenzeit bereiten sich andere zelluläre Komponenten für die Kernteilung in
ähnlicher Weise vor, wie dies bei der Mitose zu sehen ist. Die Centromere weichen
voneinander, und Spindelmikrotubuli werden zwischen ihnen ausgebildet. Die
Kernmembran löst sich auf. Schließlich nehmen die Spindelmikrotubuli Kontakt mit den
Kinetochoren auf und die Chromosomen beginnen die Ausbildung der Metaphaseplatte. Die
Prophase I belegt typischerweise 90% der Zeit, die für die Meiose benötigt wird.
Die Chromosomen sind nun auf der Metaphaseplatte angeordnet, und noch immer sind die
Homologen gepaart. Die Kinetochormikrotubuli des einen Zellpols sind mit je einem
8Chromosom jedes Paares verbunden, während Mikrotubuli des anderen Zellpols mit dem
jeweils anderen Chromosom Kontakt aufnehmen.
Wie bei der Mitose transportiert der Spindelapparat die Chromosomen während der
Anaphase I zu den Polen. Die Schwesterchromatiden bleiben jedoch über ihr Centromer
miteinander verbunden und bewegen sich als eine Einheit zum selben Pol. Die homologen
Chromosomen bewegen sich zum entgegengesetzten Pol. Dies ist anders als während der
Mitose. In der Mitose erscheinen die Chromosomen eher einzeln auf der Metaphaseplatte
anstatt in Paaren, und es werden die Schwesterchromatiden jedes Chromosoms getrennt.
Während der Telophase I fährt der Spindelapparat fort, die homologen Paare zu trennen, bis
die Chromosomen schließlich die Zellpole erreichen. An jedem Pol sammelt sich nun ein
haploider Satz, aber jedes Chromosom – die jetzt in Wirklichkeit nicht mehr einzeln zu
erkennen sind – besteht immer noch aus zwei Chromatiden. Gewöhnlich ereignet sich die
Cytokinese gleichzeitig mit der Telophase I, wobei zwei Tochterzellen gebildet werden. Bei
Tierzellen ensteht durch Einschnüren der Plasmamembran eine Teilungsfurche. Vor dem
Abschluss der Meiose II erfolgt nun keine weitere Replikation des genetischen Materials.
In der Prophase II bildet sich ein neuer Spindelapparat, und die Chromosomen bewegen sich
zur Metaphaseplatte. Die Chromosomen ordnen sich auf der Metaphaseplatte an wie bei
einer Mitose, wobei die Kinetochore der Schwesterchromatiden jedes Chromosoms zu
entgegengesetzten Polen zeigen. In der Anaphase II trennen sich die Schwesterchromatiden
am Centromer und bewegen sich zu entgegengesetzten Zellpolen. Während der Telophase II
beginnen sich an den beiden Zellpolen die Kerne zu bilden, und es findet Zytokinese statt. Es
sind nun vier Tochterzellen vorhanden, wovon jede einen haploiden individuellen
Chromosomensatz trägt.
Abbildung 4 Der menschliche Chromosomensatz oder Karyotyp (griech. Kerngepräge) mit 23 diploiden Chromosomen.
Beachte, dass die Chromosomen verschieden gefärbt/ gebändert sind und unterschiedlich groß sind. Auch die Lage des
Centromers ist unterschiedlich.
9Denkaufgabe 3: Wie sieht das Karyogramm eines Patienten mit Downsyndrom aus und wie
heißt die entsprechende Chromosomenabberation (lat. Abweichung)? In welcher Phase der
Meiose kann eine solche Chromosomenabberation entstehen? Recherchiere gerne im
World Wide Web.
Zum Abschluss sollen hier noch einmal die grundlegenden Unterschiede der Mitose und der
Meiose zusammengefasst werden: Die Chromosomenzahl wird bei der Meiose auf die Hälfte
reduziert, nicht aber bei der Mitose. Die genetischen Konsequenzen dieses Unterschiedes
sind wichtig. Während durch die Mitose zwei Zellen entstehen, die genetisch identisch mit
der elterlichen somatischen Zelle sind, gehen aus der Meiose vier Zellen hervor, die sich
genetisch sowohl von der elterlichen Gameten und als auch untereinander unterscheiden.
RNA
RNA besteht genauso wie DNA aus Nukleotiden, die miteinander verbunden sind. Ein
Nukleotid besteht aus einer Base, einem Zucker und einem Phosphatrest. Trotzdem gibt es
Unterschiede zwischen DNA und RNA:
- Während DNA aus zwei komplementären Strängen besteht und eine Doppelhelix
bildet, ist RNA einzelsträngig und nimmt verschiedene Formen an.
- Die vier Basen der DNA Nukleotide sind Adenin, Guanin, Thymin und Cytosin. Bei
RNA wird Thymin durch Uracil (U) ersetzt.
- Die Pentose in DNA ist Desoxyribose, wohingegen die Pentose in RNA Ribose ist.
Es gibt drei Haupttypen von RNA:
1. messenger-RNA (mRNA), eine Blaupause der DNA, die die DNA Information zur
Proteinsynthese übermittelt.
2. ribosomale RNA (rRNA), die zusammen mit Proteinen Ribosomen (siehe
Proteinbiosynthese/Translation) bilden.
3. transfer-RNA (tRNA), die zur Proteinsynthese Aminosäuren zum Ribosom liefern.
Proteinbiosynthese
Die drei Teile der Proteinbiosynthese, postuliert im Dogma der Molekularbiologie, werden in
den folgenden Videos näher beschrieben.
Transkription: https://www.youtube.com/watch?v=Fy_3gpIkoNs
Prozessierung: https://www.youtube.com/watch?v=HSdK7l9Qk1w
Translation: https://www.youtube.com/watch?v=3wFfj6D0_nQ
Proteine
Proteine (=Eiweiße) sind wichtige Effektormoleküle und entscheidend für die
Expressionsregulation des verfügbaren genetischen Materials. Sie stabilisieren Zellen, lesen
Gene ab oder werden als Signale an andere Zellen versandt. In diesem Abschnitt wird
10erklärt, wie ein Protein aufgebaut ist, welche Strukturen es annehmen kann und in welche
Klassen wir Proteine einteilen können.
Aminosäuren
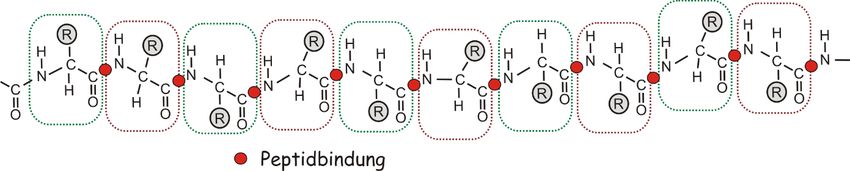
Wie in den Videos zur Proteinbiosynthese gezeigt, bilden Ketten von Aminosäuren (engl.
amino acid, AA) die Peptide. Längere Peptide mit mehr als 100 AA werden als Proteine
bezeichnet. In Proteinen verbinden sich Aminosäuren derartig, dass ein repetitives
Stickstoff-Kohlenstoff-Kohlenstoff-Rückgrat das gesamte Protein durchzieht. An diesem
Rückgrat stehen in regelmäßigen Abständen Reste ab, die je nach AA variieren können
(Abbildung 5). Diese Reste sind es, die durch ihre chemische Eigenschaften einen
wesentlichen Teil der Struktur des Proteins bestimmen. So gibt es zum Beispiel hydrophile
Reste, die mit einer wässrigen Lösung interagieren können (z. B. Serin, Glutamin) und
deshalb bevorzugt auf Proteinoberflächen liegen, denn die zelluläre Umwelt ist zumeist
wässriger Natur. Auch gibt es hydrophobe Reste, die von Wasser abgestoßen werden (z. B.
Valin und Alanin), die bevorzugt im Inneren von Proteinen und in Membranproteinen
(zelluläre Membrane sind Lipiddoppelschichten) positioniert sind. Weitere Reste bilden
starke Ionenbindungen (z. B. Glutamat, Arginin) oder sogar echte Atombindungen
(ausschließlich Cystein). Menschliche Zellen verwenden 20 Aminosäuren in ihren Proteinen.
In der wissenschaftlichen Literatur werden sie in einer Kurzschreibweise dargestellt, die
entweder aus drei oder sogar nur aus einem Buchstaben besteht. Alanin mit dem Rest -CH3
zum Beispiel wird als Ala oder schlicht mit A abgekürzt. Solche Tabellen muss man nicht
wissen, denn es gibt sie tausendfach im Web abzurufen.
Abbildung 5 Das Rückgrat eines Proteins. Die gestrichelten Kästchen grenzen einzelne Aminosäuren voneinander ab, die an
den Stellen der Peptidbindung (roter Kreis) miteinander verbunden sind. Die Aminosäuren unterscheiden sich voneinander
durch die chemischen Gruppen, die an der Reststelle (grauer Kreis) angehängt sind.
Struktur der Proteine
Die Abfolge der Aminosäuren bestimmt die Form oder Konformation des Proteins. Dabei ist
der Faltungsvorgang selbst derart komplex, dass selbst mit Hochleistungscomputern nicht
die endgültige Konformation aus der Aminosäuresequenz bestimmt werden kann. Die
Konformation wird durch vielerlei Wechselwirkungen zwischen Resten des
Proteinrückgrates festgelegt, wobei das fertige Protein zumeist ein energetisches Minimum
erreicht. Die komplexe Form der Proteine wurde der Einfachheit halber in verschiedene
Strukturklassen unterteilt, die in Abbildung 6 dargestellt sind. Die Primärstruktur beschreibt
die bloße Abfolge der Aminosäuren. Um die beiden Enden der AA-Kette unterscheiden zu
können, sprechen wir von einem N-Terminus (dort, wo die Kette mit einem Stickstoff- Atom
beginnt, siehe Abbildung 5) und einem C-Terminus (dort, wo die Kette mit einem
11Kohlenstoffatom endet). Die Sekundärstruktur ergibt sich aus
Wasserstoffbrückenbindungen zwischen den AA, die nicht von den Resten erzeugt werden,
sondern vom Sauerstoff am C-terminalen Kohlenstoff einer jeden AA und dem Wasserstoff
am Stickstoffatom. Aus diesen Wechselwirkungen resultieren zwei mögliche Formen der AA-
Kette: die α-Helix und das β-Faltblatt. Die Wechselwirkungen zwischen den Resten der AA,
die oben geschildert wurden, spielen auf der nächsten Komplexitätsebene die
entscheidende Rolle, der Tertiärstruktur.
Abbildung 6 Konfigurationen eines Proteins. Primärstruktur = Abfolge der AA; Sekundärstruktur = alpha-Helix und beta-
Faltblatt; Tertiärstruktur = Interaktion der AA-Reste.
Wenn wir als Forscher mehr über die Struktur oder Funktion eines Proteins erfahren wollen,
so verwenden wir häufig die Seite http://uniprot.org. Mit dieser Seite lassen sich folgende
Aufgaben lösen: Welche Primärstruktur oder Sequenz besitzt die Homeobox protein Hox-
B13? Suche nach dem Protein, indem du die Abkürzung HXB13_HUMAN im Suchfeld von
Uniprot eingibst. Welche Strukturelemente der Sekundärstruktur erkennst du wieder?
Schaue dir auch die gefaltete Struktur als Bild unter
https://www.ebi.ac.uk/pdbe/entry/pdb/5EGO an. Findest du die α-Helices und die DNA?
Welches andere Homeobox Protein heterodimerisiert mit dem Homeobox Protein B13?
Proteinfunktion
Proteine sind die zentralen Bausteine jeder Zelle. Eine Übersicht über die mannigfaltigen
Proteinfunktionen gibt folgende Tabelle.
Klasse Funktion Beispiel
Strukturproteine Bausteine von Zellen und Aktin ist ein Strukturprotein
Geweben des Zytoskeletts
Signalproteine Signalübertragung zwischen Wnt-Faktoren regulieren eine
oder innerhalb von Zellen Vielzahl von
Entwicklungsprozessen
Regulatorische Proteine Transkriptionsfaktoren, die Oct-4 ist Teil eines Cocktails
DNA binden und dabei Gene mit dem Stammzellen
an und ausschalten induziert werden
Transportproteine Transport kleiner Moleküle Connexine tragen zum
oder Ionen interzellulären Signalaustausch
über die Membran hinweg bei
Enzyme Katalyse biochemischer Hyaluronidase kann
Reaktionen extrazelluläre Matrix des
Bindegewebes verdauen
12Denkaufgabe 4: Welcher Klasse in dieser Tabelle ist das oben genannte Homeobox Protein
B13 zuzuordnen? Recherchiere gerne auf Uniprot!
Einführung in die Immunologie
Die konstante Interaktion mit der Umwelt hat zur Ausbildung eines komplexen
Immunsystemes geführt. Das menschliche Immunsystem besteht konzeptionell aus zwei
Teilen, dem angeborenen Immunsystem, das eine erste unspezifische Barriere bildet, und
dem adaptiven Immunsystem, das ein Gedächtnis besitzt.
Angeborenes Immunsystem
Das angeborene Immunsystem dient der schnellen Abwehr jeglicher Erreger. Die ersten
Barrieren, die vor Erregern schützen, sind physikalischer, chemischer und mechanischer
Natur: die Haut trennt uns physikalisch von unserer Umwelt, die Magensäure zersetzt
mögliche Erreger, die mit der Nahrung aufgenommen werden chemisch, und der Niesreiz
hält unsere Lunge von Partikeln mechanisch frei. Wenn ein Erreger diese Barrieren
überquert hat und in das Gewebe eindringt, werden spezielle Immunzellen aktiv. Zu diesen
Immunzellen gehören Granulozyten, Makrophagen (griech. Fresszellen) und natürliche
Killerzellen (Abbildung 7). Zu den Hauptaufgaben der Granulozyten und Makrophagen
gehört es, einen Entzündungsherd zu etablieren und mit Enzymen die Erreger anzugreifen
und zu verdauen. Die primäre Aufgabe natürlicher Killerzellen ist es, infizierte körpereigene
Zellen abzutöten, sodass sich ein Erreger nicht weiter ausbreiten kann.
Abbildung 7 Granulozyt (links), Macrophage (mittig) und natürlicher Killerzelle (rechts) gehören zum angeborenen
Immunsystem. Quelle: https://images.fineartamerica.com/images-medium-large-5/immune-cells-illustration-spencer-
sutton.jpg
Adaptives Immunsystem
Das adaptive Immunsystem ist ein individuell spezialisiertes Immunsystem, das Erreger-
spezifisch reagiert. Es entwickelt sich erst nach der initialen Immunantwort des
angeborenen Immunsystems, hilft aber bei einer wiederkehrenden Infektion schnell und
gezielt die Immunabwehr zu verstärken. Zu den spezialisierten Zellen des adaptiven
Immunsystems gehören die dendritischen (griech. Baum) Zellen, zwei Arten von T-
Lymphozyten (griech. weiße Blutzellen) und die B-Lymphozyten (Abbildung 8). Dendritische
13Zellen haben eine vergrößerte Oberfläche, mit der sie Antigene des Erregers aufnehmen
können. Nach der Prozessierung der Antigene präsentieren sie den T-Helfer Lymphozyten
Erreger-spezifische Peptide. T-Helfer Lymphozyten, die diese Erreger erkennen, werden
positiv selektioniert und helfen die Immunantwort der B-Lymphozyten zu verstärken. Die
Hauptaufgabe von B-Lymphozyten ist die Produktion von Antikörpern. Diese können
extrazellulär Erregerantigene spezifisch binden, sodass der Organismus den Erreger
schneller erkennen und eliminieren kann. Zytotoxische T-Lymphozyten identifizieren
Erreger-spezifisch infizierte Zellen und töten sie ähnlich wie natürliche Killerzellen ab.
Abbildung 8 Dendritische Zelle (links), T-Zelle (mittig), B-Zelle (rechts) gehören zum adaptiven Immunsystem. Quelle:
https://images.fineartamerica.com/images-medium-large-5/immune-cells-illustration-spencer-sutton.jpg
Motivation!
Geschafft! Raucht euch jetzt der Kopf? Macht euch keine Sorgen, wenn ihr irgendwo nicht
weitergekommen seid. Mit Sicherheit habt ihr jetzt mehr Fragen als zuvor. Entweder ihr
hebt euch eure Fragen bis zur Akademie auf, wo wir unser bestes geben werden, sie gut zu
beantworten. Oder ihr schreibt uns eure Fragen an uta-hardt[at]msn[dot]com.
Bis zum 23. Juli 2018 in Papenburg!
Viele Grüße,
Eure Kursleiter Uta und Johanna
14Quellen und Anmerkungen
Molecular Biology of the Cell; Alberts, Johnson, Lewis, Raff, Roberts, Walter; 52008, Garland
Science.
http://www.nobelprize.org/educational/medicine
Taschenlehrbuch Biologie; Munk; 12008, Thieme.
Biologie; Campbell, Reece, Markl; 62006, Pearson Studium.
http://www.jörn-online.de/
https://study.com/academy/course/high-school-biology-help-course.html
http://www.zum.de/
Anmerkungen:
- Der Text zur Mitose orientiert sich stark am Taschenlehrbuch Biologie, der Text zur
Meiose ist beinahe gänzlich einer Abbildung aus dem Lehrbuch Biologie entnommen.
- Florian Schober hat den Text zu den Proteinen verfasst.
15Sie können auch lesen

















































