Klimawandel und Baumarten-Verwendung für Waldökosysteme
←
→
Transkription von Seiteninhalten
Wenn Ihr Browser die Seite nicht korrekt rendert, bitte, lesen Sie den Inhalt der Seite unten
Forschungsstudie
Klimawandel und
Baumarten-Verwendung für
Waldökosysteme
Auftraggeber: Stiftung Wald in Not
Projektleiter: Prof. Dr. Andreas Rolo
Bearbeiter: Dipl.-Forstwirtin Britt Grundmann
Technische Universität Dresden
Institut für Forstbotanik und Forstzoologie,
Professur für Forstbotanik
Pienner Str. 7
01737 Tharandt
rolo@forst.tu-dresden.de
www.forst.tu-dresden.de/Botanik
08t.08.2008Forschungsstudie TU-Dresden: Klimawandel und Waldbaumartenverwendung
Inhaltsverzeichnis
1 Einleitung 4
2 Methode 5
2.1 Frostresistenz . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6
2.2 Hitze- und Trockenheitsresistenz . . . . . . . . . . . . . . . . . . . . . . . 12
3 Ergebnisse und Diskussion 14
3.1 Betrachtung der Hauptbaumarten . . . . . . . . . . . . . . . . . . . . . . 14
3.1.1 Rot-Buche (Fagus sylvatica L.) . . . . . . . . . . . . . . . . . . . 14
3.1.2 Stiel-, Trauben-Eiche (Quercus robur L. / Quercus petraea [Matt.]
Liebl. bzw. Quercus robur L. subsp. robur / Quercus robur L.
subsp. sessili
ora (Salisb.) A. DC.) . . . . . . . . . . . . . . . . . 15
3.1.3 Gemeine Fichte (Picea abies [L.] Karst.) . . . . . . . . . . . . . . 16
3.1.4 Wald-Kiefer (Pinus sylvestris L.) . . . . . . . . . . . . . . . . . . 16
3.2 Andere Baumarten . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 17
3.2.1 Neben- und Mischbaumarten . . . . . . . . . . . . . . . . . . . . . 17
3.2.2 Baumarten südländischer Herkunft . . . . . . . . . . . . . . . . . 18
3.2.3 Baumarten fremdländischer Herkunft . . . . . . . . . . . . . . . . 19
3.3 Herausforderungen für den Waldbau im Klimawandel . . . . . . . . . . . 20
3.4 Bewertung der Baumarten . . . . . . . . . . . . . . . . . . . . . . . . . . 20
3.4.1 Beispiele . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20
3.4.2 Gesamtbewertung . . . . . . . . . . . . . . . . . . . . . . . . . . . 22
3.5 Physiologische Anpassungsfähigkeit . . . . . . . . . . . . . . . . . . . . . 26
4 Schlussfolgerungen und KLAM-Wald (KLimaArtenMatrix für
Waldbaumarten) 28
Literatur 38
2Forschungsstudie TU-Dresden: Klimawandel und Waldbaumartenverwendung
Zusammenfassung
In der vorgelegten Studie werden 47 Waldbaumarten hinsicht-
lich ihrer Verwendbarkeit für Waldökosysteme unter Bedingungen
des Klimawandels bewertet. Diese Zusammenstellung beruht auf
der integrativen Auswertung und Beurteilung bereits publizierter
Analysen und Daten zu den physiologischen und ökologischen Po-
tentialen der Arten.
Hierbei liegt das wesentliche Augenmerk auf der Toleranz gegen-
über andauernden Trockenphasen, aber auch auf der Frostresis-
tenz. Um eine allgemeine Frosthärte jeder Art zu ermitteln, wur-
de die Toleranz gegenüber Winterfrösten, ebenso wie gegenüber
Spätfrostereignissen herangezogen.
Im Ergebnis können 47 Baumarten mittels Benotung (1-4) für vier
Standorte unterschiedlicher Wasserversorgung bewertet werden.
Dies wird anschaulich zusammengefasst in der KLAM-Wald, der
KLimaArtenMatrix für Waldbaumarten. Danach zeigt sich, dass
die meisten der heimischen Arten auf geeigneten Standorten ein
hohes Potential haben, um auch in Zukunft stabile Wälder zu bil-
den. Bei Entscheidungen der Baumartenwahl sind selbstverständ-
lich auch weitere Faktoren wie z.B. Nährstoangebot, Höhenstufe
und bei einzelnen Baumarten auch mögliche Pathogenrisiken mit
einzubeziehen.
Eine Bewertung der Waldbaumarten unter dem Aspekt des Kli-
mawandels wie in dieser Studie wurde bisher noch nicht vorgenom-
men, so dass es sich um einen neuen Ansatz handelt, der hiermit
zur Diskussion gestellt werden soll.
3Forschungsstudie TU-Dresden: Klimawandel und Waldbaumartenverwendung
1 Einleitung
Die Prognosen zum Klimawandel fallen je nach angewendetem Szenario unterschiedlich
dramatisch aus [35]. Im Zeitraum von 1906-2005 stieg die globale Jahresmitteltempera-
tur um 0,74◦ C an. Sie soll sich relativ zu 1980-1999 nochmals um 2-4◦ C bis zum Jahr
2100 erhöhen. Mit einer erhöhten Temperatur geht eine Veränderung der Niederschläge
einher. Während in Europa die Niederschläge in den Wintermonaten zunehmen werden,
kommt es zu einem Rückgang der Niederschläge in der Vegetationsperiode um 10-25 %
(SRES Bezugsszenario A1B). Allerdings bestehen bei regionalen Niederschlagsprojek-
tionen noch erhebliche Unsicherheiten. Die Zunahme von Extremereignissen, wie lange
Trockenperioden, Orkane, Starkregen und Über
utungen, ist bereits spürbar. Die Zu-
nahme von Sturmereignissen deutet sich für Europa ebenfalls an [21, 48, 49].
Klimatisch bedingte Veränderungen unserer Waldökosysteme sind nur eingeschränkt vor-
hersagbar, da groÿräumige Klimafaktoren durch regionale und lokale Dierenzierungen
wie z.B. Wasserspeicherfähigkeit der Böden modi
ziert werden [2]. Die Wälder stehen
vor einem bisher noch nie da gewesenem Anpassungsdruck [10]. In der vergangenen Erd-
geschichte gab es immer wieder einen Wechsel zwischen Warm- und Kaltzeiten, doch
vollzog sich dieser niemals in einer Geschwindigkeit, wie sie jetzt prognostiziert wird
[68]. Auf besonderen Extremstandorten, wie im Kanton Wallis in der Schweiz, einem der
extremsten inneralpinen Trockengebiete, lassen sich bereits heute Veränderungen in der
Artenzusammensetzung beobachten [69]. Veränderungen im Konkurrenzgeschehen der
Baumarten in den verschiedenen Entwicklungsstufen (Blüte, Regeneration, Wachstum)
werden in Zukunft sehr wahrscheinlich unterschiedlich verlaufen [2, 54]. Die Arealgren-
zen der Baumarten werden sich in Zukunft verschieben, nicht nur horizontal, sondern
auch vertikal [19, 96, 101].
Welche Baumarten sind nun für die deutschen Wälder in Zukunft geeignet? Welche
Bäume reagieren besonders tolerant oder auch intolerant auf Witterungsextreme wie
z.B. Trockenperioden oder Spätfröste und sind dennoch an tiefe Wintertemperaturen
angepasst? Diese individuelle Toleranz ist nicht nur artabhängig, sondern auch die Her-
künfte einer Art unterliegen dem Ein
uss unterschiedlicher Lokaladaption [13, 81, 90].
Der zukünftige Waldbau muss daher Baumarten mit bestmöglicher Anpassungsfähigkeit
auswählen. Die zunehmende Hitze in der Vegetationsperiode und das Verschieben der
gröÿten Niederschlagsmengen in das Winterhalbjahr [2, 36] stellen zusätzliche Stresssi-
tuationen für die Bäume dar. Im bestmöglichen Fall führt dies zu Resistenzen der P
an-
zen, doch kann es auch zu Zuwachsverlusten, sichtbaren Schäden, sogar bis hin zum
4Forschungsstudie TU-Dresden: Klimawandel und Waldbaumartenverwendung
Absterben und zum gänzlichen Ausfall einer Art kommen. Künftig werden auf einigen
Standorten Baumarten nicht mehr geeignet sein und auf andere Standorte ausweichen,
oder es werden andere Arten hinzukommen [94, 95, 100]. Zudem ist davon auszugehen,
dass andere Nutzungsaspekte neben der Holznutzung an Bedeutung gewinnen werden,
z.B. ökologische und ästhetische Funktionen.
2 Methode
Eine sinnvolle Herangehensweise an eine objektive Bewertung wichtiger Baumarten hin-
sichtlich der Verwendungsmöglichkeiten für Waldökosysteme in naher Zukunft ist die
Analyse von Verbreitungsgebieten und Klimadiagrammen, sowie das Einschätzen des
physiologischen und ökologischen Potentials einer Art. Hierbei wurde besonderes Au-
genmerk auf die Toleranz gegenüber andauernden Trockenphasen gelegt. Hierzu wurden
verschiedene Literaturquellen herangezogen [11, 17, 41, 42, 70, 75, 77, 78, 82, 97, 98].
Ein weiterer Aspekt für nützliche Hinweise auf die Fähigkeiten einer Baumart, Trocken-
phasen zu überstehen, ist ihre Eignung als Stadtbaum [22, 7074, 76]. Die klimatischen
Bedingungen innerhalb der Städte wirken um ein Vielfaches verstärkt und unmittelbar
auf den Einzelbaum. Daher wurde diese Eignung ebenfalls in die allgemeine Bewertung
mit einbezogen. Der ökologische Waldbau hingegen musste sich schon immer an den
gegebenen Standortsverhältnissen orientieren. Daher
ieÿt das potenzielle Vorkommen
jeder einzelnen Baumart in natürlichen Waldgesellschaften, wohinter sich die Eignung
für frische oder trockene Standorte verbirgt, in die Bewertung mit ein [17, 80, 82]. Es
werden folgende vier Standortsbereiche unterschieden: nass bis sehr frisch, ziemlich frisch
bis frisch, mäÿig frisch bis mäÿig trocken und trocken bis sehr trocken.
In der vorliegenden Studie wurde ebenfalls der Ansatz der Klimahüllen berücksichtigt [5,
37]. Doch aufgrund der kritisch zu sehenden Eingangsdaten fallen die Einschätzungen des
Potentials einzelner Baumarten unterschiedlich zu unseren Untersuchungen aus (Kap.
3.1.4). Aus diesem Grund fanden diese Bewertungen keinen Eingang in die vorliegenden
Ergebnisse.
Um zusätzlich eine Einschätzung der Frosthärte jeder einzelnen Baumart zu gewinnen,
wurde mittels obiger Literatur die Toleranz gegenüber Winterfrösten, aber auch Spät-
frostereignissen ermittelt. Diese beiden Eigenschaften wurden gemittelt und zu einer
allgemeinen Frosthärte zusammengefasst. Wird die Frosthärte einer Baumart als sehr
gut oder gut eingeschätzt, hat sie auf die abschlieÿende Gesamtbewertung keinen wei-
teren Ein
uss. Ist eine Baumart jedoch in ihrer allgemeinen Winterfrostresistenz einge-
5Forschungsstudie TU-Dresden: Klimawandel und Waldbaumartenverwendung
schränkt, oder ist sie spätfrostemp
ndlich, so hat das eine wichtige Auswirkung auf die
Gesamteinschätzung: In solch einem Falle wurde die Bewertung um eine Stufe herabge-
setzt.
Im Folgenden sollen die wesentlichen Eigenschaften Frostresistenz sowie Hitze- und Tro-
ckenheitsresistenz, die zu einer Bewertung der Baumarten führen, erläutert werden.
2.1 Frostresistenz
Frostresistenz kann allgemein de
niert werden als die Fähigkeit einer P
anze, Tempera-
turen unter 0◦ C ungeschädigt zu überleben [46]. Sie ist in erster Linie genetisch bedingt
und weist Unterschiede zwischen Arten, Sorten und sogar Klonen auf. Weiterhin un-
terliegt sie einer typischen Jahresrhythmik, ist abhängig vom Abhärtungsvermögen und
verändert sich im Laufe des Lebensalters der P
anze. Auch exogene Faktoren wie Nähr-
stoangebot, Licht, Wasser und Immissionen beein
ussen die Frosthärte. Der Photo-
und Thermoperiodismus einer Baumart bestimmt in der Regel deren Frosthärte. Durch
hormonale Steuerung tritt der Baum im Herbst in das Stadium der Vorruhe, welche bei
vielen Arten durch Dauerbelichtung noch reversibel ist. Unter dem Ein
uss von weiteren
Hemmstoen beginnt anschlieÿend die Vollruhe. Diese Phase des Übergangs bestimmt
die Frosthärte, die durch komplexe Veränderungen in der plasmatischen Struktur der
Zellen gekennzeichnet ist. So kann durch eine künstliche Verkürzung der Tageslängen
eine P
anze früher ausreifen und dadurch einen höheren Frosthärtegrad erreichen. Dies
konnte bei Robinia pseudoacacia durch Moschkow bereits 1935 nachgewiesen werden.
Andererseits kann eine kurze Tageslänge und die damit eintretende Ruheperiode bei
Picea glauca die Dürreresistenz fördern [91].
Der Kältetod ist, wie auch der Hitzetod, die Folge von Biomembranläsionen und des
Zusammenbruchs der Energieversorgung der Zelle. Eine Schädigung durch Frost tritt
ein, wenn es zur Eisbildung in den Zellen kommt (intrazelluläre Eisbildung). Auch die
extrazelluläre Eisbildung in den Interzellularen (Kondenswasser) und Zellwänden kann
schädigen (Abb. 1), doch ist es artspezi
sch unterschiedlich, wie lange eine P
anze dies
ertragen kann.
Der Gefrierimpuls breitet sich über Leitbündel und homogene Gewebe schnell aus, Dis-
kontinuitäten (Lufträume, dichte verholzte oder cutinisierte Sekundärwände) behindern
die Eisausbreitung dagegen. Die physiologischen Folgen dieser Eisbildung sind die De-
hydration, die Veränderung der Permeabilität von Biomembranen und die Anreicherung
6Forschungsstudie TU-Dresden: Klimawandel und Waldbaumartenverwendung
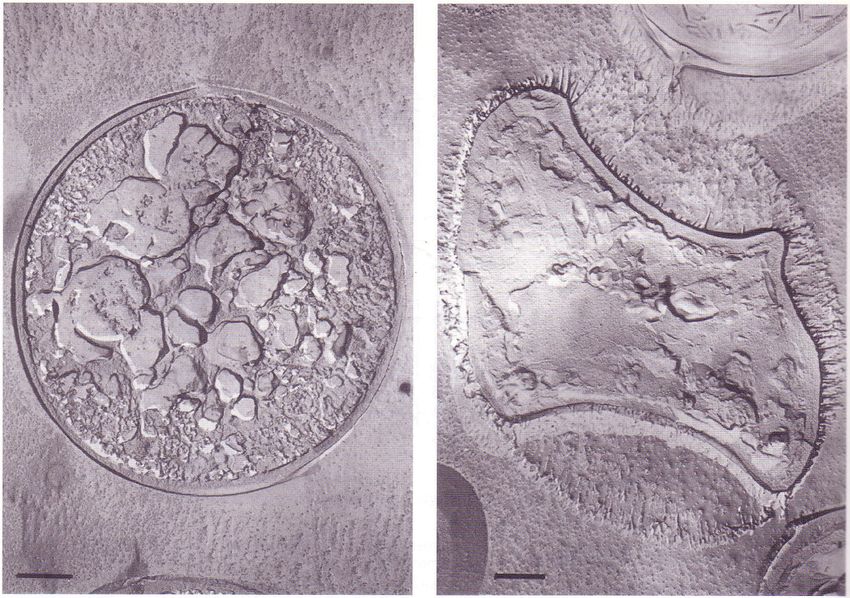
Abbildung 1: Intrazelluläres (links) und extrazelluläres Gefrieren (rechts) von Hefezellen.
Durch intrazellulär entstandenes Eis werden die protoplasmatischen Struk-
turen zerstört. Die extrazellulär gefrorene Zelle ist durch den Wasserentzug
geschrumpft und von Eiskristallnadeln umgeben, die den ursprünglichen
Umfang der Zelle erkennen lassen. (Längenmaÿ = 1 µm) Quelle: LAR-
CHER 2001
von kryotoxischen Substanzen. Dies kann im Zusammenhang mit der Inaktivierung mem-
brangebundener Enzymsysteme zum Zusammenbruch des Energieumsatzes sowie zu wei-
teren Schädigungen von Membranen führen.
P
anzen haben in ihrer Anpassung an frostgefährdete Standorte eine Vielzahl von Mög-
lichkeiten entwickelt, um Frostereignisse und Frostperioden zu überstehen. Dazu gehören
Maÿnahmen gegen den Gefrierstress, aber auch gegen Begleiterscheinungen winterli-
cher Kälte wie Photoinhibition, Frosttrocknis und Schneebelastung. Zur Fähigkeit einer
P
anze, Winterschäden vorzubeugen, gehört ebenso, sich der Frosteinwirkung so gut wie
möglich zu entziehen, räumlich wie auch zeitlich [89]. Hierzu gehört z.B. das Abwerfen
der frostemp
ndlichen Blätter.
Eine Frostresistenz kann auf zwei verschiedene Wege erreicht werden:
1. durch die Fähigkeit, Eisbildung in Zellen und Geweben (intrazelluläre Eisbildung)
zeitlich zu verzögern oder gegen tiefere Temperaturen zu verschieben (Gefrierver-
zögerung);
7Forschungsstudie TU-Dresden: Klimawandel und Waldbaumartenverwendung
2. durch die Fähigkeit, extrazelluläre Eisbildung zu tolerieren (Gefrierbeständigkeit).
Die meisten Bäume der gemäÿigten Breiten besitzen die Eigenschaft, dass einzelne ihrer
Gewebe und Organe oder auch zeitweilig die gesamte P
anze extrazelluläre Eisbildung
ertragen. Dies erreichen sie hauptsächlich durch die Stabilisierung der Membransysteme.
Folgende biochemisch-physiologischen Vorgänge lassen sich dabei beobachten [47, 89]:
1. Bildung von Kryoprotektoren (Substanzen, die das osmotische Potential der Zellen
beein
ussen und somit zur Gefrierpunktsenkung beitragen);
2. Veränderung und Stabilisierung der Membranen selbst (durch Anreicherung pola-
rer Lipide, Desaturation von Membranlipiden, um die Membran
uidität bei nied-
rigen Temperaturen aufrechtzuerhalten und durch Anreicherung von Proteinen);
3. Vermehrung und Vergröÿerung von Biomembran
ächen (Schutzfunktion bei Struk-
turveränderungen durch extrazelluläre Gefrierprozesse);
4. Persistente Unterkühlung in Geweben mit Nukleationsbarrieren (dicke und dichte
Zellwände ermöglichen eine anhaltende Gefrierverzögerung bis zur Unterschreitung
einer Temperaturschwelle. Bei manchen Blättern bis auf -10◦ C und -12◦ C, bei
Knospen und Holz verschiedener Wald- und Obstbäume der gemäÿigten Zone bis
auf -30◦ C bis -70◦ C).
Die Fähigkeit der Frostresistenz der untersuchten Baumarten ist Tabelle 1 zu entnehmen.
Hierbei wurden Noten von 1 - 4 vergeben, wobei gilt
• 1 = extrem frostresistent
• 2 = gut frostresistent
• 3 = bedingt frostresistent
• 4 = frostemp
ndlich.
Diese Bewertung wurde getrennt vorgenommen für die Spätfrostresistenz und die all-
gemeine Winterfrostresistenz. In der letzten Spalte wurde der Mittelwert aus beiden
Eigenschaften gebildet, um eine Gesamteinschätzung für jede Art zu gewinnen. Diejeni-
gen Baumarten, bei denen entweder die allgemeine Frosthärte oder die Spätfrostresistenz
mit 3 oder 4 bewertet wurde, sind fett hervorgehoben und sind somit als frostgefährdet
gekennzeichnet.
8Tabelle 1: Bewertung der Frostresistenz, Baumarten mit eingeschränkter Winterhärte oder Spätfrostresistenz sind fett her-
vorgehoben (Erläuterungen der Ziern oben im Text)
Forschungsstudie TU-Dresden: Klimawandel und Waldbaumartenverwendung
Lateinischer Name Deutscher Name Spätfrost Winterfrost Mittelwert
Abies alba Mill. Weiss-Tanne 4 2 3
Abies grandis (Dougl. ex D.Don) Lindl. Küsten-Tanne 3 2 2,5
Acer campestre L. Feld-Ahorn 2 2 2
Acer platanoides L. Spitz-Ahorn 2 1 1,5
Acer pseudoplatanus L. Berg-Ahorn 2 1 1,5
Alnus glutinosa (L.) Gaertn. Schwarz-Erle 2 2 2
Alnus incana (L.) Moench Grau-Erle 1 1 1
Betula pendula Roth Sand-Birke 1 1 1
Betula pubescens Ehrh. Moor-Birke 1 1 1
Buxus sempervirens L. Buchsbaum 2 1 1,5
9
Carpinus betulus L. Hainbuche 2 1 1,5
Castanea sativa Mill. Edel-Kastanie 3 2 2,5
Fagus sylvatica L. Rot-Buche 3 2 2,5
Fraxinus excelsior L. Gemeine Esche 3 2 2,5
Fraxinus ornus L. Blumen-Esche 3 3 3
Ilex aquifolium L. Stechpalme 4 2 3
Juglans regia L. Gemeine Walnuss 4 2 3
Larix decidua Mill. Europäische Lärche 2 1 1,5
Malus sylvestris (L.) Mill. Wild-Apfel 3 2 2,5
Picea abies (L.) Karst. Gemeine Fichte 2 2 2
Fortsetzung ...Lateinischer Name Deutscher Name Spätfrost Winterfrost Mittelwert
Pinus cembra L. Zirbel-Kiefer 1 1 1
Forschungsstudie TU-Dresden: Klimawandel und Waldbaumartenverwendung
Pinus nigra Arnold Schwarz-Pappel 1 2 1,5
Pinus strobus L. Weymouths-Kiefer 1 1 1
Pinus sylvestris L. Wald-Kiefer 1 1 1
Populus nigra L. Schwarz-Pappel 2 2 2
Populus tremula L. Zitter-Pappel 1 1 1
Prunus avium L. Vogel-Kirsche 2 2 2
Prunus padus L. Trauben-Kirsche 2 1 1,5
Pseudotsuga menziesii (Mirb.) Franco Douglasie 2 1 1,5
Pyrus pyraster (L.) Burgsdorf Wild-Birne 3 2 2,5
Quercus cerris L. Zerr-Eiche 3 2 2,5
Quercus petraea (Matt.) Liebl. Trauben-Eiche 2 2 2
10
Quercus pubescens Willd. Flaum-Eiche 2 3 2,5
Quercus robur L. Stiel-Eiche 2 1 1,5
Quercus rubra L. Rot-Eiche 2 2 2
Robinia pseudoacacia L. Robinie 2 2 2
Salix alba L. Silber-Weide 2 1 1,5
Sorbus aria (L.) Crantz Mehlbeere 2 1 1,5
Sorbus aucuparia L. Eberesche 1 1 1
Sorbus domestica L. Speierling 2 2 2
Sorbus torminalis (L.) Crantz Elsbeere 2 2 2
Taxus baccata L. Gemeine Eibe 2 2 2
Tilia cordata Mill. Winter-Linde 2 1 1,5
Fortsetzung ...Lateinischer Name Deutscher Name Spätfrost Winterfrost Mittelwert
Tilia platyphyllos Scop. Sommer-Linde 2 2 2
Forschungsstudie TU-Dresden: Klimawandel und Waldbaumartenverwendung
Ulmus glabra Huds. emend. Moss. Berg-Ulme 2 1 1,5
Ulmus laevis Pall. Flatter-Ulme 2 2 2
Ulmus minor Mill. emend. Richens Feld-Ulme 2 2 2
11Forschungsstudie TU-Dresden: Klimawandel und Waldbaumartenverwendung
2.2 Hitze- und Trockenheitsresistenz
Im Allgemeinen wird unter Hitzeresistenz die Fähigkeit verstanden, hohe Temperaturen
ohne irreversible Schäden zu überstehen. Die maximalen Temperaturen, die Gehölze in
den gemäÿigten Breiten ertragen können, liegen bei 45◦ C bis 55◦ C [89]. Unter Dürre
versteht man dagegen eine niederschlagsarme Zeit, während der der Wassergehalt des
Bodens so stark absinkt, dass die P
anzen unter Wassermangel leiden. Häu
g ist Bo-
dentrockenheit mit Lufttrockenheit und starker Einstrahlung gekoppelt, wodurch eine
hohe Verdunstungsbelastung entsteht. Groÿräumig ist Trockenheit das Ergebnis des
Zusammenwirkens von Niederschlagsmangel und hoher Verdunstung. In Trockengebie-
ten herrscht Dürre mit solcher Regelmäÿigkeit und von solcher Dauer, dass die jährliche
Verdunstung die Jahressumme der Niederschläge übersteigt [47].
Der Schädigungsverlauf durch Trockenheit beginnt mit einer Volumenverminderung der
Zelle durch Wassermangel. Es folgt eine Erhöhung der Zellsaftkonzentration und eine
Entquellung des Protoplasmas. Der Turgor der Zelle geht somit zurück und es kommt
zu einer Verlangsamung der Wachstumsvorgänge. Trockenschäden sind unvermeidbar,
sobald das Wurzelwachstum aufgrund Wassermangels eingestellt wird. Wie schon zuvor
genannt ist der Hitzetod die Folge von Biomembranläsionen und des Zusammenbruchs
der Energieversorgung der Zelle.
Trockenheitsvermeidung können P
anzen durch Einziehen oder Abwerfen austrocknungs-
emp
ndlicher Organe (alle vegetativen Organe, Sprosssysteme, Blätter) erreichen. An-
dererseits können Überdauerungsorgane mit spezi
schen Mechanismen der Trockenre-
sistenz ausgestattet werden.
Zur Aufrechterhaltung des lebensnotwendigen Wassergehalts dienen folgende funktionel-
le und strukturelle Eigenschaften [89]:
1. Schutzeinrichtungen gegen unkontrollierbare Wasserverluste
• Kutikula und Wachsau
age auf den Blättern
• Behaarung von Blättern, vorrangig tote Haare
• Blattstellung
• Kork und Borke an Stämmen
• Endodermis und Exodermis an Wurzeln
• Reduktion transpirierender Flächen
12Forschungsstudie TU-Dresden: Klimawandel und Waldbaumartenverwendung
2. Einschränkung der Wasserabgabe durch Regulation der Stomata
3. Sicherung der Wasseraufnahme
4. Verbesserung des Wassertransports
5. Wasserspeicherung
Fähigkeiten der P
anzen, die der Austrocknung auf zellulärer und subzellulärer Ebene
entgegenwirken können, sind folgende:
1. Aufrechterhaltung des Turgordrucks von Zellen durch Akkumulation gelöster, häu-
g osmotisch wirksamer Substanzen;
2. Elastizität von Geweben;
3. Fähigkeit des Protoplasmas zur nahezu vollständigen Austrocknung bei Aufrecht-
erhaltung der Aktivität löslicher Enzyme für Photosynthese und Respiration.
Die Befähigung der begutachteten Bäume, Hitze- und Trockenstress zu ertragen, ist in
deren Eignung, auf unterschiedlich frischen Standorten vorkommen zu können, enthal-
ten. Diese generelle Eignung wurde nach umfangreichem Literaturstudium und eigenen
Erhebungen für folgende Standorte eingeschätzt:
• nass bis sehr frisch
• ziemlich frisch bis frisch
• mäÿig frisch bis mäÿig trocken
• trocken bis sehr trocken
Die Benotung wurde in gleicher Form von 1 bis 4 vorgenommen, wobei gilt:
• 1 = sehr gut geeignet
• 2 = gut geeignet
• 3 = bedingt geeignet
• 4 = ungeeignet
13Forschungsstudie TU-Dresden: Klimawandel und Waldbaumartenverwendung
3 Ergebnisse und Diskussion
Zu den Möglichkeiten der Vorbereitung von bestehenden Waldbeständen unter Beibe-
haltung der Baumartenzusammensetzung auf zukünftige Bedingungen gehören Anpas-
sungen der waldbaulichen Behandlung wie das Absenken der Umtriebszeiten oder Weit-
bestandeskonzepte. Daneben ist eine Strategie zur Risikominimierung durch ökologisch
ausgerichteten Waldumbau eine der wichtigsten Optionen [50]. Dies erfordert ein Min-
destmaÿ an Mischung sowie den Anbau von Baumarten, die bei aller Unsicherheit des
Kommenden am ehesten in der Lage zu sein scheinen, sowohl mit den Veränderungen in
der Zeit des Übergangs, als auch mit den später eventuell konstanteren aber veränderten
Klimabedingungen zurecht zu kommen, und natürlich eine standortgerechte, klimaange-
passte Baumartenwahl. Eine weitere Option des Waldbaus stellt das Naturverjüngungs-
potential dar. So wird die natürliche Selektion auf Individuen begünstigt, die gegenüber
den veränderten Bedingungen die gröÿte Toleranz besitzen [2]. Grundsätzlich wird die
Klimaveränderung keinen bis wenig Ein
uss auf das Baumwachstum haben, wenn sich
eine Baumart an einem Standort in ihrem Optimum be
ndet. Lediglich an Standorten,
wo Bäume an ihre physiologischen Grenzen geraten, muss die Forstwirtschaft eingreifen
[15].
3.1 Betrachtung der Hauptbaumarten
Die Bewertung des Verwendungspotentials der Baumarten für Waldökosysteme soll an-
hand einer Benotung in Tabellenform erfolgen. Doch zunächst werden die vier ökono-
misch wichtigsten Baumarten Rot-Buche (Fagus sylvatica L.), Stiel- und Trauben-Eiche
(Quercus robur L. / Quercus petraea [Matt.] Liebl.), Gemeine Fichte (Picea abies [L.]
Karst.) und Wald-Kiefer (Pinus sylvestris L.) näher betrachtet.
3.1.1 Rot-Buche (Fagus sylvatica L.)
Eine der wichtigsten Laubbaumarten Deutschlands und Mitteleuropas war bisher die
Rot-Buche, und sie wird ihre Stellung voraussichtlich auch in naher Zukunft behaup-
ten können [1, 19]. Die Buche wächst auch heute schon auÿerhalb ihres physiologischen
Optimums in bestandsbildender Funktion, und es wird ein weiteres Vordringen auf tro-
ckenwarme Kalkstandorte verzeichnet. Auch das Verschieben der Arealgrenze kann beob-
achtet werden. Dies zeigten u.a. Peñuelas und Boada (2003) für die Buche im Montseny
Gebirgszug in Nordspanien. Hier hat sich die Verbreitung um 70 Höhenmeter erweitert.
14Forschungsstudie TU-Dresden: Klimawandel und Waldbaumartenverwendung
Die Buche besitzt eine erstaunlich hohe Trockenstresstoleranz und kann nicht zu lang an-
haltende trockene Phasen gut tolerieren [9, 13, 14]. Gegenüber Kiefer und Eiche zeichnet
sie sich durch eine hohe Konkurrenzkraft aus, welche sie nur bei langen und häu
gen Tro-
ckenperioden verliert. Doch gibt es auch konträre Ansichten zur zukünftigen Verbreitung
der Buche. So soll die Konkurrenzfähigkeit in zunehmend kontinentalerem Klima und
weiterhin bestehender Spätfrostgefährdung gegenüber Eiche, Linde und Gemeine Hain-
buche abnehmen [52]. Jedoch wird das Anpassungspotential der Buche an Trockenstress
oft unterschätzt. So kann es zu reversiblen Veränderungen von Gestaltsmerkmalen in
Form von Verengung des Spross-Wurzel-Verhältnisses, aber auch zu Wachstumsreduk-
tionen kommen [9]. Diese Möglichkeit zu evolutionären Anpassungsprozessen besteht
aufgrund der sehr hohen genetischen Variabilität der Buche und durch Änderung gene-
tischer Strukturen. Auf extremen Trockenstandorten wird jedoch sehr wahrscheinlich der
Anteil der Buchenwaldgesellschaften gegenüber den Eichen-Trockenwäldern zurückgehen
[3].
3.1.2 Stiel-, Trauben-Eiche (Quercus robur L. / Quercus petraea [Matt.] Liebl.
bzw. Quercus robur L. subsp. robur / Quercus robur L. subsp. sessili
ora
(Salisb.) A. DC.)
Die Eiche (Stiel- und Trauben-Eiche) wird oftmals als die Baumart angesehen, die an-
gesichts des Klimawandels und des zukünftig erhöhten Anteils an Trockenstandorten
pro
tieren wird [58]. Sie ist mit ihrer tiefreichenden Pfahlwurzel prädestiniert für die
Trockenstandorte im Nordostdeutschen Tie
and, wo sie gemeinsam mit der heute dort
vorherrschenden Kiefer stabile Mischwälder bilden könnte. Ebenso im Süden Branden-
burgs und im Norden Sachsens wird es zukünftig teilweise für die Buche zu trocken
werden. Auch hier wäre die Eiche eine sehr gut geeignete Baumart. Dennoch verblei-
ben auch oene Fragen, wie die der möglichen Veratmung von gespeicherter Stärke in
warmen Wintern, oder auch die Wirkung von Kiefern-Eichen-Mischwäldern auf das Bo-
densubstrat. Doch hier zeichnen sich bereits einige wertvolle Vorteile dieser Baumarten-
mischung ab. Die oberen Bodenschichten in solchen Mischwäldern neigen weniger stark
zu Versauerung als bei fehlender Laubstreu, und das organische Material wird deutlich
besser in den Mineralboden eingetragen. Weiterhin verbessert die Laubstreu die Benetz-
barkeit des Bodens, denn ein Boden, der über einen bestimmten Schwellenwert hinweg
ausgetrocknet ist, nimmt nicht so leicht wieder Wasser an.
Zimmermann et al. (2006) modellierten für verschiedene Klimaszenarien das Vorkom-
men und die Ausbreitung verschiedener Gehölze in der Schweiz. Generell wird es zu einer
15Forschungsstudie TU-Dresden: Klimawandel und Waldbaumartenverwendung
Verschiebung vieler Areale in höhere Lagen kommen, und aufgrund der abnehmenden
Sommerniederschläge werden nicht trockenheitstolerante Laubbaumarten benachteiligt
werden. Bei einer mittleren Temperaturerhöhung um 2,3 bis 2,8◦ C erwarten sie eine
Ausdehnung der wärmetoleranten Eichen- und Eichen-Buchenwälder im Schweizer Mit-
telland. Desweiteren stellen sie fest, dass das Hauptverbreitungsgebiet der Trauben-Eiche
in der Schweiz eine massive Verschiebung in Richtung der Voralpen erfahren wird. In
vielen Gebieten in der mittleren Schweiz wird es nach dem SRES Szenario A1FI (+6,0◦ C
± 0,29◦ C) [35] jedoch vermutlich sogar für die Trauben-Eiche zu trocken werden.
3.1.3 Gemeine Fichte (Picea abies [L.] Karst.)
Die Baumart, die für Deutschland wohl das schlechteste Anpassungspotential mit sich
bringt, ist die Gemeine Fichte. Als eine unserer ökonomisch bisher wichtigsten Baumar-
ten wird sie viele Standorte nicht mehr ohne Probleme besiedeln können, was jedoch
hauptsächlich im Anbau auf ungeeigneten Standorten begründet ist [85]. Ein weiteres
Problem stellt ebenso die hohe Kalamitätsanfälligkeit der Fichte dar [51]. Dennoch wird
verzeichnet, dass die Fichte vom zunehmenden CO2 -Gehalt pro
tiert [30], doch führen
warme und trockene Perioden zu einem Rückgang in Oberhöhe und Gesamtzuwachs.
Die Fichte ist eine der Baumarten, die besonders emp
ndlich auf heiÿ-trockene Sommer
reagiert [16]. Im Rekordsommer 2003 zeigte sie auf den langfristigen Level II-Flächen
von ICP Forests in Zentraleuropa im Vergleich zu Buche und Tanne die stärksten Rück-
gänge im Zuwachs [20]. Aber genau wie die Weiÿ-Tanne oder auch der Berg-Ahorn ist
die Fichte weiterhin ein wichtiger Bestandteil der Bergmischwälder [23, 38, 62, 67]. Nach
Döbbeler und Spellmann (2002) sowie Prot et al. (2006) wird im Tertiären Hügelland,
im Sauerland, im Niedersächsische Harz und im Thüringer Wald die Fichte auch in
Zukunft die Hauptbaumart bleiben.
3.1.4 Wald-Kiefer (Pinus sylvestris L.)
In einem groÿen Teil Deutschlands, im Nordostdeutschen Tie
and, ist die Wald-Kiefer
die dominierende Waldbaumart. Daher wird ihre zukünftige Stellung viel diskutiert.
Hofmann und Jenssen (2007) schreiben, dass mit zunehmender Bodennährkraft (Stick-
stoeinträge), Bodenfrische und Ozeanität des Klimas der relative Anbauwert der Kiefer
in selbstorganisierten Waldbeständen aufgrund der wachsenden Konkurrenzkraft hoch-
wüchsiger und ausdunkelnder Laubbaumarten sinkt. Walentowski et al. (2007) sehen
das Hauptverbreitungsgebiet der Kiefer im borealen Nadelwaldgürtel und bestätigen ihr
16Forschungsstudie TU-Dresden: Klimawandel und Waldbaumartenverwendung
ein hohes Anpassungspotential an Fröste, Trockenheit und Waldbrand. Dennoch soll
im westlichen und zentralen Europa die Kiefer unter dem Ein
uss der Erwärmung und
der Eutrophierung weiter zurückgehen. Weiterhin kann es dazu kommen, dass im süd-
lichen Teil Mitteleuropas ihre Konkurrenzkraft gegenüber Laubbäumen abnimmt und
die Anfälligkeit gegenüber Krankheitserregern zunimmt. Auch beobachten Rigling et al.
(2006) im Wallis in der Schweiz ein Zurückdrängen der Kiefer durch die Flaum-Eiche.
Der erhöhte Druck durch Trockenheit, Bestandeskonkurrenz, Misteln, Insekten, Phyto-
pathogene und Nematoden lässt die Kiefer auf diesem Extremstandort sogar absterben
[29], wobei die Mortalität der Kiefern im Wallis mit der Trockenheit korreliert.
Dennoch bestätigen wir der Wald-Kiefer ein besonders hohes Anpassungspotential und
bewerten ihre zukünftige Stellung in Mischwäldern als weiterhin sehr stark. Nach Döb-
beler und Spellmann (2002) wird sie im Ostniedersächsischen Tie
and, im Nordbran-
denburger Jungmoränenland und im Mittelbrandenburger Talsand- und Moränenland
auch zukünftig die Hauptbaumart bleiben. Durch die Klimahülle der Wald-Kiefer von
Kölling (2007) wird ihr keine besondere Stellung in unseren Wäldern mehr zuteil. Ein
wesentlicher Kritikpunkt an dieser Art der Herangehensweise liegt jedoch in der verwen-
deten Datengrundlage. Zur Einschätzung der Areale der Baumarten wurde die Karte
der natürlichen Vegetation Europas von Bohn und Neuhäusl (2000/2003) herangezo-
gen. Doch in dieser wird die ökologische Amplitude der Arten dargestellt und nicht die
physiologische. Die ökologische Amplitude stellt jedoch das Vorkommen einer Art unter
Konkurrenzdruck anderer Arten dar, wodurch die meisten Baumarten auf Nischenstand-
orte verdrängt werden. Somit wird das tatsächliche Potential vieler, vor allem seltener
Arten unterschätzt. Weitere Faktoren, die bisher keinen Eingang in die Klimahüllen ge-
funden haben, sind die jahreszeitliche Verteilung von Temperatur und Niederschlag und
Bodeneigenschaften, wie vor allen Dingen die Wasserspeicherkapazität.
3.2 Andere Baumarten
3.2.1 Neben- und Mischbaumarten
Auf besondere Weise wird die Rolle von Neben- und Mischbaumarten, wie z.B. Spitz-
Ahorn, Winter-Linde, Gemeine Walnuss, Vogel-Kirsche, Speierling oder Elsbeere, deren
klimatische Eignung als besonders hoch eingeschätzt werden, an Bedeutung gewinnen
[11, 12, 31, 39, 43, 60, 63, 8386, 88]. In der Regel benötigen diese Baumarten jedoch
Böden mit besserer Nährstoversorgung, so dass deren Auswahl wie auch die sämtlicher
17Forschungsstudie TU-Dresden: Klimawandel und Waldbaumartenverwendung
anderer Arten standortsgemäÿ erfolgen sollte. In Laubmischwäldern mit der Buche kön-
nen einige dieser Arten erheblich unter Konkurrenzdruck durch die dominante Buche
leiden. Arten wie die Echte Mehlbeere oder Elsbeere können sich dennoch gut in der
Mittel- und auch Unterschicht etablieren [83]. Die Förderung jeglicher Naturverjüngung
wird besonders empfohlen, da diese die natürliche Selektion auf Individuen begünstigt,
die gegenüber den veränderten Bedingungen die gröÿte Toleranz besitzen. Auch das
aktive Einbringen von Neben- und Mischbaumarten an Waldrändern führt durch nach-
folgende selbstständige Ausbreitung mittel- bis langfristig zu einem erhöhten Anteil von
diesen. Durch die Wahl geeigneter Herkünfte dieser Arten können hervorragende ver-
wertungsrelevante Eigenschaften erreicht werden.
3.2.2 Baumarten südländischer Herkunft
Bereits seit mehreren Jahrzehnten wird vermehrt eine Zunahme der Konkurrenzkraft von
Baumarten aus südlichen Gebieten Mitteleuropas beobachtet, wie Flaum-Eiche, Edel-
Kastanie, Gemeine Stechpalme oder Zerr-Eiche [45, 95, 96]. Doch es existiert auch die
Befürchtung, dass der Klimawandel schneller voranschreitet als diese Baumarten sich
bei uns ausbreiten und bestandsbildend auftreten werden [53]. Waldökosysteme sind
grundsätzlich relativ träge. Es können also weit schneller einzelne Arten ausfallen, als
dass andere hinzukommen [92]. Auÿerdem ist auch bei südländischen Baumarten die
Trockenheitsresistenz stark provenienzabhängig. Dies berichten Barthold et al. (2004)
von der Edel-Kastanie, die im Juli 2003 auf der Alpensüdseite eine Braunfärbung der
Blätter zeigte, doch diese Erscheinung trat vorwiegend auf Extremstandorten auf. Sehr
wahrscheinlich kann sie, je nach Herkunft, auf
achgründigen Standorten oder Böden
mit schlechter Wasserspeicherkapazität für Sommerdürre anfällig sein. Diese Anfälligkeit
für Dürrestress hat nicht nur eine Wachstumsreduktion zur Folge, sondern ruft auch ei-
ne Prädisposition gegenüber dem Erreger des Kastanienrindenkrebses hervor, der auch
schon auf der Alpennordseite beobachtet wurde [28], oder der Tintenkrankheit. Daher
sei auch hier bei der Baumartenwahl auf den besonderen Ein
uss der Herkunft hinge-
wiesen.
Die weit verbreitete Vermutung, dass sich das Klima nördlich der Alpen längerfristig
dem südländischen Mittelmeerklima angleichen würde, ist mit Vorsicht zu betrachten.
Mit groÿer Wahrscheinlichkeit werden viele Standorte ähnliche Witterungsbedingungen
erreichen, doch werden besonders die winterlichen Bedingungen noch länger ein Hin-
dernis für die Etablierung südländischer Baumarten darstellen. Die in Zukunft auch
18Forschungsstudie TU-Dresden: Klimawandel und Waldbaumartenverwendung
weiterhin auftretenden Spätfröste können Blüte, Fruchtreife und somit eine Naturver-
jüngung immens beeinträchtigen. Das zukünftige Klima in Deutschland, und besonders
dasjenige der Übergangsphase, wird eines sein, dass in der Form derzeit in Europa nicht
groÿräumig zu
nden ist. Es wird noch am ehesten mit Regionen Südosteuropas wie
z.B. Ungarn, dessen Klima von heiÿ-trockenen Sommern und gleichzeitig nass-kalten
Wintern geprägt ist, zu vergleichen sein. Auch wird sich das bereits heute sehr dieren-
zierte Klima innerhalb Deutschlands regional sehr unterschiedlich entwickeln. Die süd-
und westlichen Gebiete Deutschlands können auch dem Klima des Zentralmassivs in
Süd-Frankreich ähnlich werden, mit heiÿ-trockenen Sommern und nicht ganz so kalten
Wintern wie in Ungarn.
3.2.3 Baumarten fremdländischer Herkunft
Bisher wurden in Deutschland gute Erfahrungen mit einigen fremdländischen Baumar-
ten, wie Douglasie, Küsten-Tanne oder Rot-Eiche [4, 2426, 59, 99], gemacht. Das bereits
hohe klimatische Anpassungspotential dieser Baumarten und die wirtschaftliche Bedeu-
tung lieÿen sich durch Einführen noch besser geeigneter Herkünfte weiter steigern. Be-
reits etablierte Baumarten fremdländischer Herkunft wie diese bereits genannten oder
neue, trockenheitstolerante Arten aus Europa können sich als sinnvolle Ergänzung un-
seres Baumartenpools erweisen. Doch sollte der Anbau nicht heimischer, trockenheits-
toleranter Arten nach Ammer und Kölling (2007) nur versuchsweise erfolgen, da in der
klimatischen Übergangszeit noch ausschlieÿende Faktoren wie Spätfröste, Leistungsfä-
higkeit und die Auswirkungen der neuen Baumarten auf den Boden und die belebte
Umwelt zu untersuchen sind.
Ein gutes bzw. schlechtes Beispiel für das invasive Potential einer mehr oder weniger
unbekannten Spezies in einem fremden Ökosystem liefert die Nordamerikanische Dreh-
Kiefer (Pinus contorta Dougl. ex Loud.), die in den 1920er Jahren in Schweden eingeführt
wurde [18]. An diesem Versuch konnte beobachtet werden, dass sich eine neue Art un-
kontrolliert ausbreiten, heimische Arten verdrängen und für die Einfuhr fremdländischer
Insektenarten, die zu Schädlingen werden können, verantwortlich sein kann. Eine neue
Baumart kann ein bisher ausgewogenes Verhältnis zwischen heimischer Fauna und Flora
stören. Aufgrund mangelnder Erfahrung mit Baumarten aus Amerika und Asien, wel-
che bisher nicht dem Konkurrenzgefüge unserer heimischen Waldökosysteme ausgesetzt
waren, sollte auf die Einfuhr neuer Arten aus diesen Teilen der Erde weitgehend ver-
zichtet werden. Weiterhin sollten neue fremdländische Baumarten grundsätzlich, wenn
19Forschungsstudie TU-Dresden: Klimawandel und Waldbaumartenverwendung
überhaupt nötig, dann nur auf Extremstandorten eingesetzt werden, die von heimischen
Arten nicht mehr ohne Schwierigkeiten besiedelt werden können.
3.3 Herausforderungen für den Waldbau im Klimawandel
Als unverzichtbar wird sich ebenso in Zukunft die Kontrolle der Wilddichte erweisen,
denn nur so lassen sich die erwünschten Bestockungsziele erreichen. Das Wild ist oftmals
ein Verhinderer von Biodiversität, aber genauso ein Förderer dieser, je nach Selektions-
druck. Daher muss der Mensch diese Selektion in die gewünschte Richtung leiten.
Dagegen stellt die Problematik der Schadinsekten ein nahezu unkalkulierbares Risi-
ko dar, so dass deren zukünftige Entwicklung sich nicht einschätzen lässt. Aufgrund
steigender Temperaturen kann es bei einigen Arten zu Veränderungen in den Entwick-
lungszyklen kommen [87]. Da viele Arten in ihrer Entwicklung jedoch hauptsächlich von
der Photoperiode abhängig sind, kann die Situation auch konstant bleiben oder sich so-
gar zu Ungunsten der Insekten verändern. Ebenso können neue, fremdländische Arten
bei uns heimisch werden [7, 34, 40, 65]. Somit kann auch nicht ermessen werden, welche
Baumarten in Zukunft welchen pathogenen Risiken ausgesetzt sein werden. Bei Wild-
p
anzen mit natürlicher Verbreitung existiert ein Gleichgewicht zwischen Schaderregern
und Wirtsp
anzen, gelangen Krankheitserreger jedoch in neue Gebiete mit potenziellen
neuen Wirtsp
anzen, sind Epidemien nicht ausgeschlossen [27]. Die Klimaerwärmung
kann also wärmeliebende Pathogene begünstigen, und Umweltstress wie Trockenstress,
Staunässe und Bodenversauerung können die Abwehr der Bäume reduzieren. Lediglich
durch gut strukturierte, horizontal wie auch vertikal, und artenreiche Mischwälder so-
wie der Wahl geeigneter Herkünfte kann dem Risiko von Insekten- und Pilzkalamitäten
entgegengewirkt werden.
3.4 Bewertung der Baumarten
3.4.1 Beispiele
Als Ergebnis dieser Studie wurde jede einzelne Baumart auf der Basis umfassender Li-
teraturstudien hinsichtlich ihrer Frostresistenz beurteilt (Kap. 2.1) und anhand ihres
natürlichen Verbreitungsgebietes und ihres physiologischen Potentials, Standorte unter-
schiedlicher Wasserversorgung zu besiedeln, bewertet. Diese Wertung wurde durch Noten
von 1 = sehr gut geeignet bis 4 = ungeeignet (Kap. 2.2) vorgenommen. Baumarten, die
20Forschungsstudie TU-Dresden: Klimawandel und Waldbaumartenverwendung
eine eingeschränkte Frostresistenz aufweisen, wurden in die nächst tiefere Wertung abge-
stuft. Zum besseren Verständnis sollen hier zwei Beispiele aufgeführt werden, Schwarz-
Erle und Weiÿ-Tanne, um zu erläutern, wie sich die Gesamtbewertung ergibt.
Wie in Kapitel 2.1 beschrieben, wurde zunächst die allgemeine Frostresistenz ermittelt
(Tab. 2, Spalte 3). Anschlieÿend wurde durch die Analyse der natürlichen Vorkommen
und des ökophysiologischen Potentials die Eignung für vier verschiedene Standortstypen
bewertet (Spalte 2). Die Frostresistenz der Schwarz-Erle wurde mit 2 bewertet, so dass
die Standortseignung nicht eingeschränkt wird und so eine Gesamtbewertung für jeden
Standort entsteht. Die Schwarz-Erle ist demnach sehr gut bis gut geeignet für nasse bis
frische Standorte, doch für mäÿig frische bis sehr trockene Standorte ist sie ungeeignet.
Tabelle 2: Bewertung des Potentials von Schwarz-Erle (Alnus glutinosa ) für vier Stand-
orte unterschiedlicher Wasserversorgung, die Gesamtbewertung ergibt sich aus
der Standortseignung und der Frostresistenz
Standort Potential für Mittelwert Gesamtbewertung
Standort Frostresistenz
nass bis 1 2 1
sehr frisch
ziemlich frisch bis 2 2 2
frisch
mäÿig frisch bis 4 2 4
mäÿig trocken
trocken bis 4 2 4
sehr trocken
21Forschungsstudie TU-Dresden: Klimawandel und Waldbaumartenverwendung
Die Weiÿ-Tanne ist relativ winterhart, doch sie ist besonders spätfrostemp
ndlich. Dem-
nach ist ihre Frostresistenz eingeschränkt. Daher wird ihre Standortseignung herabge-
stuft. Sie ist nach dieser Bewertung eine Baumart, die für nasse wie auch für trockene
Standorte nicht geeignet ist. Doch die für sie geeigneten Standorte sind ziemlich frische
bis hin zu mäÿig trockenen, vorrangig der Mittelgebirge. (Tab. 3).
Tabelle 3: Bewertung des Potentials von Weiÿ-Tanne (Abies alba ) für vier Standorte
unterschiedlicher Wasserversorgung, die Gesamtbewertung ergibt sich aus der
Standortseignung und der Frostresistenz
Standort Potential für Mittelwert Gesamtbewertung
Standort Frostresistenz
nass bis 4 3 4
sehr frisch
ziemlich frisch bis 1 3 2
frisch
mäÿig frisch bis 1 3 2
mäÿig trocken
trocken bis 4 3 4
sehr trocken
3.4.2 Gesamtbewertung
Anhand von Spätfrostresistenz und Winterhärte wurde eine allgemeine Frostresistenz
eingeschätzt (Kap. 2.1), die mit Noten von 1 bis 4 bewertet wurden, wobei eine Abstu-
fung von 1 = extrem frostresistent bis zu 4 = frostemp
ndlich vorgenommen wurde. Die
Benotungen 1 und 2 bezeugen der Baumart also eine ausreichend gute Frostresistenz, die
in der abschlieÿenden Gesamtbewertung nicht berücksichtigt werden muss. Bei einzelnen
Baumarten, bei welchen entweder die Spätfrostresistenz oder die allgemeine Winterhärte
mit der Benotung 3 oder 4 eingeschätzt wird, ist diese Frostemp
ndlichkeit zu beach-
ten und führt demzufolge in der Gesamtbewertung zu einer um eine Stufe schlechteren
Benotung.
In der folgenden Tabelle 4 sind die Ergebnisse dieser Studie in Form der Benotung von 47
Baumarten zusammengefasst, die im zukünftigen Waldbau in Deutschland wesentliche
Rollen spielen könnten.
22Tabelle 4: Gesamtbewertung der Baumarten für ihre Verwendung auf nassen bis sehr trockenen Standorten unter dem Aspekt
des Klimawandels. Die Noten entstehen zunächst aus der grundsätzlichen Eignung für Standorte unterschiedlicher
Forschungsstudie TU-Dresden: Klimawandel und Waldbaumartenverwendung
Wasserversorgung, hinter welcher sich die Trockenresistenz verbirgt.
Hierbei gilt: 1 = sehr gut geeignet, 2 = gut geeignet, 3 = bedingt geeignet, 4 = ungeeignet.
Bei Baumarten, die in ihrer Frostresistenz eingeschränkt sind, wurde die Gesamtbewertung dementsprechend eine
Stufe schlechter eingeschätzt (*).
Art nass bis ziemlich frisch mäÿig frisch trocken bis
sehr frisch bis frisch bis m. trocken sehr trocken
Abies alba Weiÿ-Tanne * 4 2 2 4
Abies grandis Küsten-Tanne * 4 2 2 2
Acer campestre Feld-Ahorn 4 2 1 1
Acer platanoides Spitz-Ahorn 4 2 1 1
Acer pseudoplatanus Berg-Ahorn 3 1 1 2
23
Alnus glutinosa Schwarz-Erle 1 2 4 4
Alnus incana Grau-Erle 1 2 2 3
Betula pendula Sand-Birke 3 1 1 1
Betula pubescens Moor-Birke 1 2 2 3
Buxus sempervirens Gem. Eibe 4 3 2 2
Carpinus betulus Hainbuche 4 2 1 1
Castanea sativa Edel-Kastanie * 4 4 2 2
Fagus sylvatica Rot-Buche * 4 2 2 3
Fraxinus excelsior Gem. Esche * 2 2 2 3
Fraxinus ornus Blumen-Esche * 4 3 2 2
Ilex aquifolium Stechpalme * 4 3 2 3
Fortsetzung ...Art nass bis ziemlich frisch mäÿig frisch trocken bis
Forschungsstudie TU-Dresden: Klimawandel und Waldbaumartenverwendung
sehr frisch bis frisch bis m. trocken sehr trocken
Juglans regia Gemeine Walnuss * 4 2 2 2
Larix decidua Europ. Lärche 4 2 1 2
Malus sylvestris Wild-Apfel * 4 3 2 2
Picea abies Gem. Fichte 4 2 3 4
Pinus cembra Zirbel-Kiefer 4 2 1 3
Pinus nigra Schwarz-Kiefer 4 3 1 1
Pinus strobus Weymouths-Kiefer 4 2 2 1
Pinus sylvestris Wald-Kiefer 3 2 1 1
Populus nigra Schwarz-Pappel 1 1 2 4
Populus tremula Zitter-Pappel 3 1 1 1
Prunus avium Vogel-Kirsche 4 3 2 1
24
Prunus padus Trauben-Kirsche 1 1 1 3
Pseudotsuga menziesii Douglasie 4 2 2 3
Pyrus pyraster Wild-Birne * 4 3 2 2
Quercus cerris Zerr-Eiche * 4 4 2 2
Quercus petraea Trauben-Eiche 3 1 1 1
Quercus pubescens Flaum-Eiche * 4 3 2 2
Quercus robur Stiel-Eiche 2 1 2 2
Quercus rubra Rot-Eiche 4 1 1 2
Robinia pseudoacacia Robinie 4 2 1 1
Salix alba Silber-Weide 1 1 3 4
Sorbus aria Mehlbeere 4 3 1 1
Fortsetzung ...Art nass bis ziemlich frisch mäÿig frisch trocken bis
Forschungsstudie TU-Dresden: Klimawandel und Waldbaumartenverwendung
sehr frisch bis frisch bis m. trocken sehr trocken
Sorbus aucuparia Eberesche 1 1 1 2
Sorbus domestica Speierling 4 2 1 1
Sorbus torminalis Elsbeere 4 2 1 1
Taxus baccata Gem. Eibe 4 3 1 2
Tilia cordata Winter-Linde 3 1 1 1
Tilia platyphyllos Sommer-Linde 3 1 1 2
Ulmus glabra Berg-Ulme 4 1 2 2
Ulmus laevis Flatter-Ulme 2 2 2 3
Ulmus minor Feld-Ulme 2 2 2 3
25Forschungsstudie TU-Dresden: Klimawandel und Waldbaumartenverwendung
3.5 Physiologische Anpassungsfähigkeit
Das Ziel dieser Studie, bestmögliche Hinweise zur Verwendungsmöglichkeit der Baumar-
ten für zukünftig weiterhin stabile Waldökosysteme zu liefern, kann nicht mit einer
ausschlieÿlichen Bewertung der Potentiale der Arten enden. Die alleinige Einschätzung
der Anfälligkeit jeder einzelnen Baumart gegenüber p
anzenphysiologischen Stressfakto-
ren wie Temperaturextremen, Trockenheit oder Sturm, führt nicht zu einer Aussage, ob
der gesamte Waldbestand den zukünftigen Bedingungen gewachsen sein wird oder nicht.
Wie das Wort bereits besagt, handelt es sich bei Waldökosystemen um Systeme, die
auch als Ganzes bewertet werden müssen. Faktoren wie Baumartenzusammensetzung,
Konkurrenzbeziehungen, Standortgerechtigkeit und Wilddruck bestimmen die Rahmen-
bedingungen und somit die Sensitivität und Anfälligkeit des Waldes [44].
Die Anpassungsfähigkeit von Baumpopulationen beschreibt das Potential, auf bekannte
oder unbekannte Umweltbedingungen reagieren zu können. Diese physiologische Leis-
tung ist genetisch determiniert, und jener Toleranzbereich, in dem Individuen unter
gegebenen Bedingungen überleben können, wird durch die artspezi
sche, genetische Re-
aktionsnorm bestimmt. Die Grundlage einer hohen Anpassungsfähigkeit ist also eine
breite physiologische (genetische) Reaktionsnorm. Bei sich ändernden Umweltbedingun-
gen muss sich diese Norm demzufolge ändern bzw. erweitern. Dies kann auf der Ebene
der Individuen und Populationen nur durch eine genetische Neukombination und Muta-
tionen erfolgen. Doch Savolainen et al. (2007) fürchten angesichts des Klimawandels, dass
diese genetischen Anpassungsprozesse nicht schnell genug sein könnten. Bisher konnten
solche positiv wirkenden Mutationen bei Waldbaumarten nicht entdeckt werden. Doch
im Allgemeinen deuten die Freiland-Untersuchungen zur physiologischen Angepasstheit
von Baumarten auf eine hohe Variabilität der Anpassungsreaktionen hin, welche auf eine
hohe genetische Vielfalt auf Populationsebene zurückzuführen ist. Grundsätzlich schei-
nen Baumarten, die zu den Pionierbaumarten gezählt werden können, Vorteile durch die
frühere und häu
gere Frukti
kation zu haben. Denn durch diese Vermehrungsstrategie
entstehen immer wieder neue Genotypen, aus denen durch den klimatisch bedingten
Selektionsdruck die bestgeeigneten Individuen ausgewählt werden [44].
In dieser Studie, wie auch den meisten bekannten Veröentlichungen, wurde zur Ein-
schätzung des Potentials der Bäume ein statischer Bewertungsansatz verwendet. Dies
bedeutet, dass der derzeitige Anpassungsstatus der Gehölze an wesentliche Klimafakto-
ren bewertet wird. Doch bleibt hierbei unberücksichtigt, wie sich die genetischen Reak-
tionsnormen in Raum und Zeit verändern können. Wesentliche Faktoren wie Mutations-
raten, Neurekombinationen und besonders die Verbreitung und erfolgreiche Verjüngung
26Forschungsstudie TU-Dresden: Klimawandel und Waldbaumartenverwendung
nden Eingang in dynamische (evolutive) Ansätze, die jedoch bisher noch zu unkonkret
bleiben, um in eine konkrete Bewertung Eingang
nden zu können.
Klimaplastische Wälder Der Begri der Klimaplastischen Wälder wird von Jenssen
et al. (2007) verwendet, um die erwünschte Eigenschaft der Wälder,
exibel auf Klima-
änderungen zu reagieren, zu umschreiben. Doch was genau steckt da hinter? Dazu muss
zunächst der Begri ökologischen Plastizität erläutert werden. Es ist die [...] Eigen-
schaft von Ökosystemen, sich ohne längere Sukzessionen und bei gleichzeitigem Erhalt
ihrer Lebensfunktionen (C-Speicherung, Produktivität, Mikroklima etc.) unter verän-
derlichen Umweltbedingungen strukturell selbst zu organisieren. (Jenssen et al., 2007).
Mit anderen Worten: das Ökosystems passt sich an sich verändernde Umweltbedingun-
gen an. Dagegen steht der Begri der ökologischen Resilienz [33], welcher die Fähigkeit
von Waldökosystemen beschreibt, nach einer kurzfristigen Störung in den Ausgangszu-
stand zurückzukehren. Doch es ist davon auszugehen, dass sich in Zukunft kein Gleich-
gewichtszustand des Klimas einstellen wird, sondern eine neue, fortlaufende Entwicklung
statt
nden wird [92].
Die Plastizität eines einzelnen Baumes wird durch seine physiologische Angepasstheit
bestimmt und ist daher relativ eingeschränkt. Betrachtet man jedoch eine gesamte Po-
pulation, so ist die Plastizität durch deren Anpassungsfähigkeit bereits wesentlich höher,
in Abhängigkeit von der genetischen Diversität und der Breite der ökologischen Amplitu-
de der wesentlichen Baumarten. Ein klimaplastischer Wald sollte nun zusammengesetzt
sein aus Baumarten, [...] die eine gröÿere Amplitude von Umweltbedingungen in un-
terschiedlicher Richtung abpuern können und die sich einem dauerhaft ausbildenden
Trend aus sich heraus, weitgehend in Selbstorganisation oder zumindest mit einem ge-
ringen Aufwand an forstlicher Begleitung, anpassen können. (Jenssen et al., 2007).
Die klimatische Amplitude, beschreibt daneben ausschlieÿlich die klimatischen Be-
dingungen, unter denen eine Art vorkommen kann (sie ist nicht zu verwechseln mit
der ökologischen Amplitude, in welcher auch Faktoren wie Konkurrenz zwischen den
Baumarten integriert sind). Die meisten heimischen Baumarten decken aufgrund ihrer
bemerkenswert vielgestaltigen Herkunftsgebiete eine breite Amplitude von klimatischen
Verhältnissen ab [5557, 64]. Diese Eigenschaft ist die grundlegende Voraussetzung, um
mit den heimischen Baumarten klimaplastische Wälder aufzubauen.
27Forschungsstudie TU-Dresden: Klimawandel und Waldbaumartenverwendung
4 Schlussfolgerungen und KLAM-Wald (KLimaArtenMatrix für
Waldbaumarten)
Grundsätzlich zeigt sich, dass das physiologische Anpassungspotential von
Bäumen an diese Witterungsextreme deutlich höher ist als üblicherweise an-
genommen. Entscheidend ist, dass die physiologische Vielfalt der Reaktions-
muster hoch ist, so dass immer ausreichend Baumindividuen vorhanden sind,
die den Stress tolerieren. (Kätzel, 2008)
Die heimischen Waldökosysteme werden also auch in Zukunft Bestand haben und ihre
vielfältigen Funktionen erfüllen können, wenn auch ihre Artenzusammensetzung und ihre
Struktur teilweise anders gestaltet sein können oder auch müssen. Besonders die Bedeu-
tung der heute noch seltenen Neben- und Mischbaumarten wie Spitz- und Feld-Ahorn,
Vogel-Kirsche, Wild-Apfel, Elsbeere oder Speierling wird zunehmen, denn dies sind zum
gröÿten Teil Baumarten, welche die erwünschten Eigenschaften mit sich bringen. Um den
Anteil dieser Arten zu erhöhen, bietet sich an, jegliche aufkommende Naturverjüngung
zu erhalten, wie auch ein aktives Einbringen dieser Arten zunächst an den Waldrän-
dern, von wo aus sie sich selbstständig ausbreiten werden. Beide Vorgehensweisen haben
gegenüber der Saat oder P
anzung einen kosteneinsparenden Aspekt und zudem den
Vorteil des klimatisch bedingten Selektionsdruckes, der die bestgeeigneten Individuen
fördert.
Eine der Baumartenwahl gleichgewichtige Entscheidung bei der Bestockung eines Stand-
ortes ist die richtige Wahl der Herkunft. Dies wird u.a. deutlich bei der Betrachtung
der bisherigen Erfahrungen mit Baumarten südlicher Herkunft. Auch bei vermeintlich
besonders trockenheitsresistenten Arten wie Edel-Kastanie oder Flaum-Eiche ist diese
Eigenschaft stark provenienzabhängig. Bei fremdländischen Baumarten, die schon seit
Jahrzehnten bei uns etabliert sind, wie der Douglasie oder der Rot-Eiche, lieÿe sich durch
eine verbesserte Herkunftswahl eine weitere Steigerung der Wertigkeit erreichen. Bei der
Einfuhr neuer fremdländischer Arten ist jedoch sehr vorsichtig vorzugehen, da Gehölze
aus Amerika oder Asien bisher nicht dem Konkurrenzgefüge unserer heimischen Arten
ausgesetzt waren. Es fehlen z.T. Erfahrungen bezüglich der Auswirkungen einer fremden
Art auf bestehende Ökosysteme hinsichtlich deren invasiven Potentials, der Beeinträch-
tigung des Bodens oder der potenziell gleichzeitigen Einfuhr neuartiger Pathogene, die
das Gleichgewicht zwischen heimischer Flora und Fauna stören könnten. Auch sollten
fremd- und südländische Baumarten nur auf extremen Standorten eingesetzt werden, die
von den heimischen Arten nicht mehr ohne Probleme besiedelt werden können.
28Sie können auch lesen

















































