Neutrophile extrazelluläre Netze und die Aktivität der Serumnukleasen bei Patienten mit Sepsis
←
→
Transkription von Seiteninhalten
Wenn Ihr Browser die Seite nicht korrekt rendert, bitte, lesen Sie den Inhalt der Seite unten

Medizinische Fakultät
der
Universität Duisburg-Essen
Aus der Klinik für Anästhesiologie und Intensivmedizin
Neutrophile extrazelluläre Netze und die
Aktivität der Serumnukleasen bei
Patienten mit Sepsis
Inaugural - Dissertation
zur
Erlangung des Doktorgrades der Medizin
durch die Medizinische Fakultät
der Universität Duisburg-Essen
Vorgelegt von
Linda Elisabeth Cox
aus Kamp-Lintfort
2020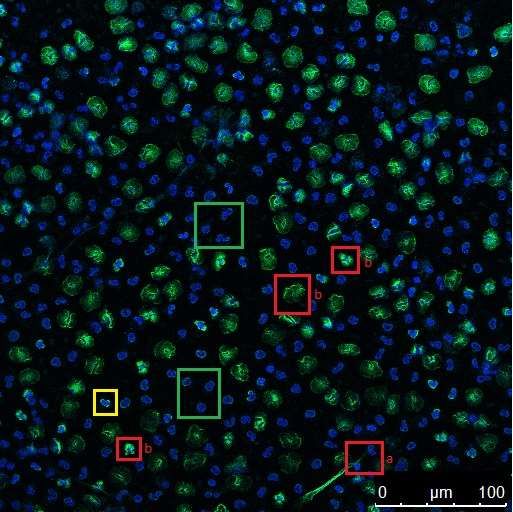
Diese Dissertation wird via DuEPublico, dem Dokumenten- und Publikationsserver der
Universität Duisburg-Essen, zur Verfügung gestellt und liegt auch als Print-Version vor.
DOI: 10.17185/duepublico/74400
URN: urn:nbn:de:hbz:464-20210623-120040-0
Alle Rechte vorbehalten.
Dekan: Herr Univ.-Prof. Dr. med. J. Buer
1. Gutachter: Herr Priv.-Doz. Dr. med. S. Schäfer
2. Gutachter: Herr Univ.-Prof. Dr. sc. hum. Dipl.-Chem. W. Brandau
Tag der mündlichen Prüfung: 23. April 2021
2
Publikationen 1 Publikationen Originalarbeiten: Neutrophil extracellular trap formation and nuclease activity in septic patients Linda E. Cox, Kai Walstein, Lena Völlger, Friederike Reuner, Alexandra Bick, An- nika Dötsch, Andrea Engler, Jürgen Peters, Maren von Köckritz-Blickwede & Simon T. Schäfer; BMC Anesthesiol. 2020 Jan 13;20(1):15. doi: 10.1186/s12871-019- 0911-7 Early suppression of peripheral mononuclear blood cells in sepsis in re- sponse to stimulation with cytomegalovirus, OKT3, and pokeweed mitogen. Malewicz NM, Walstein K, Heine T, Engler A, Bick A, Cox L, Dötsch A, Westendorf AM, Horn PA, Lindemann M, Peters J, Schäfer ST. J Appl Physiol (1985). 2019 Dec 1;127(6):1539-1547. doi: 10.1152/japplphysiol.00438.2019. Epub 2019 Sep 23 Hypoxia Inducible Factor-2 Alpha and Prolinhydroxylase 2 Polymorphisms in Patients with Acute Respiratory Distress Syndrome (ARDS). Dötsch A, Eisele L, Rabeling M, Rump K, Walstein K, Bick A, Cox L, Engler A, Bachmann HS, Jöckel KH, Adamzik M, Peters J, Schäfer ST. Int J Mol Sci. 2017 Jun 14;18(6):1266. doi: 10.3390/ijms18061266 Hypoxia inducible factor-1 alpha and prolinhydroxlase 2 polymorphisms in patients with severe sepsis: a prospective observational trial. Höcker A, Rabeling M, Bick A, Cox L, Kreuzer M, Engler A, Walstein K, Bachmann HS, Jöckel KH, Eisele L, Adamzik M, Peters J, Schäfer ST. BMC Anesthesiol. 2016 Aug 11;16(1):61. doi: 10.1186/s12871-016-0225-y
Publikationen
Abstracts:
Sepsis beeinflusst die Ausbildung von neutrophilen extrazellulären DNA-Net-
zen (Neutrophil Extracellular Traps/NETs) sowie die Serumnukleaseaktivität
Cox L.; Walstein K.; Voellger L.; Engler, A.; Höcker, A.; Naim H. Y.; von Köckritz-
Blickwede M.; Peters J.; Schäfer S.T.; Wissenschaftliche Arbeitstage der DGAI,
Würzburg, 2015.
Die Versuche wurden durchgeführt mit hilfsbereiter Unterstützung durch Frau Ur-
sula Brecklinghaus und Herrn Kai Walstein. Über Teile dieser Arbeit wurde eine
Masterarbeit durch Herrn Kai Walstein verfasst mit dem Titel: „Überwachung des
Immunstatus bei kritisch kranken Patienten: Erste Erkenntnisse abgeleitet aus der
Transplantationsimmunologie“.
4Inhaltsverzeichnis
2 Inhaltsverzeichnis
1 Publikationen .................................................................................................. 3
2 Inhaltsverzeichnis ........................................................................................... 5
3 Einleitung ........................................................................................................ 7
3.1 Sepsis ................................................................................................................ 7
3.2 Neutrophile Granulozyten ................................................................................ 8
3.3 Neutrophile extrazelluläre Netze.....................................................................10
3.4 Serumnukleasen ..............................................................................................11
3.5 Zielsetzung .......................................................................................................13
4 Material und Methoden ................................................................................. 15
4.1 Patienten und Probandendaten ......................................................................15
4.2 Blutentnahme...................................................................................................17
4.3 Isolation neutrophiler Granulozyten aus humanem Vollblut mittels
Dichtegradientenzentrifugation ......................................................................18
4.4 Hitzeinaktivierung der Staphylokokkus aureus .............................................21
4.5 Zellkultur zur Isolation von Mitochondrien ....................................................21
4.6 Isolation von mitochondrialer DNA aus kultivierten HepG2-Zellen..............22
4.7 Stimulation der NETs-Bildung ........................................................................23
4.8 NETs-Visualisierung mittels Immunfluoreszenz............................................25
4.9 Mikroskopische Beurteilung der NETs ..........................................................25
4.10 Bestimmung der Serumnukleaseaktivität ......................................................29
4.11 Statistische Methoden .....................................................................................31
5 Ergebnisse .................................................................................................... 32
5.1 Vergleich der NETs-Bildung aus arteriellem und venösem Blut ..................32
5.2 Vergleich der basalen NETs-Bildung von Probanden, Patienten mit Sepsis
und postoperativen Patienten.........................................................................33
5.3 Theoretische Gesamtbildung NETs................................................................35
5.4 Einfluss von PMA auf die NETs-Bildung von Probanden und Patienten mit
Sepsis ...............................................................................................................37
5.5 Einfluss von hitzeinaktiviertem Staphylokokkus aureus auf die NETs-
Bildung von Probanden und Patienten mit Sepsis .......................................40
5.6 Einfluss von CpG auf die NETs-Bildung gesunder Probanden ....................42
5.7 Einfluss mitochondrialer DNA auf die NETs-Bildung....................................43
5.8 Vergleich der Serumnukleaseaktivität von gesunden Probanden,
postoperativen Patienten und septischen Patienten ....................................45
5Inhaltsverzeichnis
5.9 Korrelation der Nukleaseaktivität mit pro- und antiinflammatorischen
Infektionsvariablen und dem SAPS II .............................................................47
5.10 Patienten- und Probandencharakteristika .....................................................47
6 Diskussion..................................................................................................... 51
7 Zusammenfassung ....................................................................................... 57
8 Literaturverzeichnis ...................................................................................... 58
9 Anhang .......................................................................................................... 65
9.1 Abkürzungsverzeichnis...................................................................................65
9.2 Tabellenverzeichnis.........................................................................................65
9.3 Abbildungsverzeichnis....................................................................................66
9.4 Danksagung .....................................................................................................68
9.5 Curriculum vitae ..............................................................................................69
6Einleitung
3 Einleitung
3.1 Sepsis
Alltäglich und rezidivierend sind Menschen Pathogenen aus der Umwelt ausgesetzt.
Um das Voranschreiten einer Infektion zu verhindern, ist unser Organismus auf eine
schnelle und adäquate Immunantwort angewiesen. Hierbei kann eine unzu-
reichende Antwort und Bekämpfung der Pathogene in einer Sepsis resultieren
(Seeley, Matthay & Wolters, 2012). Dabei kann der Auslöser sowohl bakteriellen,
viralen, mukösen als auch parasitären Ursprungs sein. Der Begriff „Sepsis“ ist keine
Erfindung der Neuzeit. Er existiert schon seit etwa 2700 Jahren und hat sich in sei-
ner Bedeutung kaum verändert. Ursprünglich leitet sich die Bezeichnung „Sepsis“
von dem griechischen Wort „sepo“ ab und bedeutet „ich verfalle“ (Geroulanos &
Douka, 2006).
Heutzutage ist die Sepsis immer noch ein sehr ernstzunehmendes Krankheitsbild
und eine der Haupttodesursachen auf nicht-kardiologischen Intensivstationen
(Rangel-Frausto et al., 1995). Die Anzahl der weltweit auftretenden Sepsisfälle wird
auf jährlich 31 Millionen und die einer schweren Sepsis auf 19 Millionen geschätzt
(Fleischmann et al., 2016). Damit ist die Sepsis in den USA der zweithäufigste
Grund für eine Aufnahme ins Krankenhaus (Torio & Moore, 2016). In den Industrie-
nationen erliegen immer noch etwa ein Viertel der an einer schweren Sepsis er-
krankten Patienten ihrem Krankheitsbild, was weltweit jährlich zu schätzungsweise
5,3 Millionen Todesfällen führt (Fleischmann et al., 2016). Betroffen sind in erster
Linie Personen über 65 Jahre. Auch aus finanzieller Sicht stellt die Sepsis eine
enorme Belastung dar. Mit der Behandlung der Sepsis gingen in den USA im Jahr
2013 Kosten in Höhe von 23,6 Milliarden Dollar einher (Torio & Moore, 2016).
Lange Zeit galt die Sepsis als eine überschießende Antwort des Immunsystems und
wurde mit Immunsuppressiva behandelt, um dem Krankheitsgeschehen Einheit zu
bieten. Durch die Entwicklung moderner Therapieverfahren konnte die Überlebens-
zeit unter einer Sepsis so weit verlängert werden, dass häufig von einem zweipha-
sigen Erkrankungsverlauf gesprochen wird. In der frühen Phase der Sepsis über-
wiegt die starke proinflammatorische Antwort des Immunsystems, wobei wir kürzlich
zeigen konnten, dass auch initial bereits immunsuppressive Mechanismen beste-
hen (Schäfer et al., 2016).
7Einleitung
Im weiteren Verlauf kommt es zu einer verstärkten, immunsuppressiven Phase
(Boomer et al., 2012; Boomer et al., 2011; Hotchkiss, Monneret & Payen, 2013;
Ward, 2011), in der es zum vermehrten Auftreten von Infektionen mit minderpatho-
genen Keimen kommt (Otto et al., 2011). In der frühen Phase (Tag 1-5) ereignen
sich ein Drittel aller Todesfälle und zwei Drittel in der späten (Tag 6-150) (Otto et
al., 2011). In dieser immunsuppressiven Phase ist vor allem die Ausbildung eines
(Multi-)Organversagens mit besonders schweren Verläufen einer Sepsis assoziiert.
Wenngleich dieses Modell zur Verständlichkeit beiträgt, sind die zwei Phasen keine
unabhängigen Entitäten, sondern können überlappend und bereits zu Beginn der
Erkrankung auftreten.
3.2 Neutrophile Granulozyten
Weiße Blutkörperchen, die Leukozyten, stellen den zellulären Anteil des Immunsys-
tems dar. Das Immunsystem teilt sich in einen angeborenen und einen im Verlauf
des Lebens erworbenen Anteil. Die größte Fraktion in der Blutzirkulation besteht mit
etwa 50-70 % bei Gesunden aus neutrophilen Granulozyten (Smith, 1994). Diese
gehören zum angeborenen Teil des Immunsystems. Es sind enddifferenzierte Zel-
len, mit einer sehr kurzen Lebenszeit von wenigen Stunden in der Zirkulation. Sie
besitzen einen in mehrere Läppchen geteilten Zellkern, welcher ihnen auch den Na-
men polymorphnukleäre Zellen verschaffte (Brinkmann & Zychlinsky, 2012). In ih-
rem Zytoplasma sind mehrere Granula enthalten, die unter anderem mit verschie-
denen antimikrobiell wirksamen Proteinen gefüllt sind (Nathan, 2006). Ihr Entste-
hungsort liegt im Knochenmark, wo etwa 109 Neutrophile pro kg Körpergewicht je-
den Tag ausreifen und das Knochenmark in die Blutzirkulation verlassen (Dancey,
Deubelbeiss, Harker & Finch, 1976).
Neutrophile Granulozyten nehmen eine entscheidende Rolle in unserem Immun-
system ein und sind die ersten Immunzellen am Ort einer Infektion (Nathan, 2006;
Schleimer et al., 1989; Smith, 1994). Bei einer Infektion kommt es zu einer akuten
Entzündungsantwort des angeborenen Immunsystems, durch die aktivierte Zellen
und Moleküle aus den Blutgefäßen zum Infektionsort wandern (Delves & Roitt,
2000). Die Neutrophilen folgen dabei einem Chemotoxin-Gradienten und sammeln
sich im infizierten Gewebe an (Delves & Roitt, 2000). Ihre Aufgaben sind vielfältig
8Einleitung
und bestehen zum einen aus der Kommunikation mit anderen Zellen des Immun-
systems und damit auch der Regulation der Aktivität von dendritischen Zellen, wie
Makrophagen und Monozyten (Bennouna, et al., 2003; Soehnlein et al., 2008). Zum
anderen zählt die Phagozytose zu ihren Aufgaben und somit die intrazelluläre Zer-
störung der Mikroben (Fuchs et al., 2007). Nach der Phagozytose werden die Mik-
roben in so genannten Phagosomen eingeschlossen, welche mit den Granula fusi-
onieren. Dadurch werden die phagozytierten Erreger einer hohen Konzentration an
toxischen Molekülen, wie Superoxid-Anionen, ausgesetzt und abgetötet (Delves &
Roitt, 2000). Können Superoxid-Anionen aufgrund eines genetischen Defektes der
NADPH-Oxidase nicht gebildet werden, wie bei Patienten mit einer chronischen gra-
nulomatösen Erkrankung, leiden sie an schweren Infektionen (Brinkmann V. and
Zychlinsky A., 2007; Heyworth, Cross & Curnutte, 2003), was die Bedeutung der
neutrophilen Granulozyten unterstreicht. Die wichtige Rolle der Neutrophilen spie-
gelt sich auch darin wider, dass ihre verminderte Aktivität bei Patienten mit einer
Sepsis die Entwicklung nosokomialer Infektionen begünstigt (Hotchkiss et al., 2013;
Stephan et al., 2002).
In der frühen proinflammatorischen Phase einer Sepsis kommt es zu einer massiven
Zytokinausschüttung, dem so genannten Zytokinsturm (Chong & Sriskandan,
2011). In dieser Phase sind Neutrophile stärker aktiviert als bei gesunden Proban-
den und eine schwächere Aktivität geht mit einer erhöhten Mortalität einher (Muller
Kobold et al., 2000). In diesem Zeitraum kommt es außerdem meist zu einer aus-
geprägten Rekrutierung von neutrophilen Granulozyten, durch vermehrte Aus-
schwemmung aus dem Knochenmark, sowie zu einer Verlängerung der Lebenszeit,
hervorgerufen durch eine verzögerte Apoptose (Tamayo et al., 2012). Dies führt
während der proinflammatorischen Phase meist zu stark erhöhten Neutrophilenzah-
len in der Zirkulation (Drifte et al., 2013; Hotchkiss et al., 2013; Phillipson & Kubes,
2011; Tamayo et al., 2012). In der späten immunsuppressiven Phase einer Sepsis
ist die Funktion der Neutrophilen eingeschränkt (Boomer et al., 2011; Stephan et
al., 2002) und es kann eine Neutropenie auftreten. Neben diesen klassischen Me-
chanismen ist seit einigen Jahren bekannt, dass neutrophile Granulozyten soge-
nannte neutrophile extrazelluläre Netze (NETs) bilden können, also extrazelluläre
DNA-Stränge apparent werden.
9Einleitung
3.3 Neutrophile extrazelluläre Netze
Im Jahr 2004 wurde erstmals von Brinkmann et al. ein neuer antimikrobieller Me-
chanismus der neutrophilen Granulozyten beschrieben. Sie sind der Lage, extrazel-
luläre netzartige Strukturen auszubilden und so auch extrazellulär Erreger zu immo-
bilisieren und abzutöten (Brinkmann et al., 2004). Diese neutrophil extracellular
traps oder NETs umgeben die Ursprungszelle wie eine Wolke und erstrecken sich
somit über einen Bereich, der zehn- bis fünfzehnmal so groß ist wie die Ursprungs-
zelle (Brinkmann and Zychlinsky, 2007; Brinkmann & Zychlinsky, 2012). Sie enthal-
ten sowohl nukleäre als auch granuläre Komponenten (Brinkmann et al., 2004). Ihre
Hauptbestandteile sind DNA und damit verknüpfte Histone. Außerdem enthalten sie
weitere toxische Bestandteile, wie neutrophile Elastase, Cathepsin G und Myelo-
peroxidase, welche granulären Ursprungs sind (Brinkmann et al., 2004). NETs ha-
ben einen Durchmesser von etwa 17 nm, mit kugelförmigen Verdickungen von 30-
50 nm, welche sich stellenweise zu Strängen mit bis zu 100 nm Durchmesser zu-
sammen lagern (Brinkmann and Zychlinsky, 2007; Brinkmann et al., 2004). Die Ver-
dickungen werden dabei von den granulären Proteinen gebildet (Brinkmann et al.,
2004) und ermöglichen die leichte Unterscheidung von ähnlichen Strukturen, wie
Fibrin (Brinkmann & Zychlinsky, 2012).
Die Bildung von NETs ist ein schneller Prozess, der schon zehn Minuten nach Ak-
tivierung der Neutrophilen beginnen kann (Brinkmann et al., 2004). Sind Erreger
erst einmal in den NETs gefangen und immobilisiert, sind sie einer hohen lokalen
Konzentration an antimikrobiellen Molekülen ausgesetzt (Brinkmann et al., 2004).
NETs sind so nicht nur in der Lage, Erreger in vivo einzufangen und mit Hilfe von
Proteasen, wie neutrophiler Elastase, unschädlich zu machen, sondern auch direkt
abzutöten (Brinkmann et al., 2004). Ihr Wirkspektrum ist dabei sehr vielfältig und
erstreckt sich über gram-positive und gram-negative Bakterien (Brinkmann et al.,
2004), sowie über Pilze (Urban et al., 2006) und auch Parasiten (Guimarães-Costa
et al., 2009). Die effektive bakterizide, also Bakterien abtötende, Fähigkeit von His-
tonen, einem der Hauptbestandteile von NETs, ist schon lange bekannt (Hirsch,
1958). Die antimykotische Wirkung wird durch die granulären Proteine ausgelöst
(Urban et al., 2006). Es gibt vermehrt Hinweise darauf, dass auch das Einfangen
der Erreger und damit die Verhinderung ihrer Ausbreitung, zu den entscheidenden
Aufgaben der extrazellulären Netze gehört (Meng et al., 2012; Urban et al., 2006).
10Einleitung
Die großflächige Ausdehnung ermöglicht es ihnen vermutlich auch Keime abzutö-
ten, die zu groß sind für eine Phagozytose (Urban et al., 2006). Die antimikrobielle
Aktivität der extrazellulären Netze ist der Phagozytose in ihrer Effektivität nicht un-
terlegen (Fuchs et al., 2007). Sie ermöglichen es den Neutrophilen sogar noch über
ihren Tod hinaus Erreger wirkungsvoll abzutöten (Fuchs et al., 2007).
Die Bildung von NETs ist ein NADPH-abhängiger und aktiver Prozess, der zum Tod
der Neutrophilen führt. Diese Form des gesteuerten Zelltodes wird als NETose be-
zeichnet und ist abzugrenzen von den Mechanismen der Apoptose und Nekrose
(Fuchs et al., 2007). Bei der NETose wird die Integrität der nukleären und granulä-
ren Zellmembranen aufgelöst und es kommt zur Fusion von granulären Komponen-
ten mit dem Chromatin. Die Zelle bleibt dabei so lange lebendig, bis die Zellmem-
bran ebenfalls rupturiert und die DNA-Stränge aus der Zelle ausgeschleust werden
(Fuchs et al., 2007).
Es wurde bereits eine Vielzahl an Erregern identifiziert, die zu einer Bildung von
NETs führen (Brinkmann & Zychlinsky, 2012). Dazu gehören verschiedene gram-
positive (Fuchs et al., 2007) und gram-negative Bakterien, Pilze, Parasiten und so-
gar das HI-1-Virus (Saitoh et al., 2012).
Trotz der effektiven antimikrobiellen Wirkung der NETs gibt es vermehrt Hinweise
darauf, dass neutrophile Granulozyten auf diesen Mechanismus erst zurückgreifen,
wenn sich der Organismus in einer sehr ernsten Lage befindet (Clark et al., 2007).
Vor allem in der frühen Phase einer Sepsis scheinen NETs eine wichtige Rolle zu
spielen, was im Mausmodell bereits bewiesen wurde (Meng et al., 2012). Hier wur-
den die höchsten NETs-Serumspiegel 24 Stunden nach Induktion der Sepsis ge-
messen. Diese führen zu einer abgeschwächten systemischen Entzündungsreak-
tion, verhindern die Ausbreitung der Erreger, vermindern die Schädigung lebens-
wichtiger Organe und reduzieren die Mortalität in dieser Phase (Meng et al., 2012).
3.4 Serumnukleasen
Unter dem Begriff Nukleasen wird eine Gruppe von Enzymen zusammengefasst,
die in der Lage ist, Nukleinsäuren, also DNA oder RNA hydrolytisch zu spalten und
dadurch abzubauen. Desoxyribonuklease I (DNase I) ist die in menschlichen Kör-
perflüssigkeiten, wie Serum oder Urin, am häufigsten vorkommende Nuklease. Ihr
11Einleitung
Produktionsort liegt in den Speicheldrüsen und im Pankreas, welche für gewöhnlich
einen Serumspiegel um 3 ng ml-1 aufrecht erhalten (Prince et al., 1998). Eine Auf-
gabe der DNase liegt in dem Abbau von fremder DNA, die über den Verdauungs-
trakt in den Körper gelangt. Mit Entdeckung von NETs wurde klar, dass diese DNa-
sen auch endogene extrazelluläre DNA abbauen, indem sie das „DNA-Rückgrat“
der NETs degradieren (Brinkmann et al., 2004).
Der Abbau von DNA-Netzen in der Zirkulation ist von großer Bedeutung. Denn kann
die DNase I ihren Aufgaben nicht vollständig nachkommen, wie bei Patienten mit
Systemischem Lupus Erythematodes (SLE) nachgewiesen, führt dies durch die
Ausbildung einer sekundären Lupusnephritis zu einem besonders schweren Krank-
heitsverlauf (Hakkim et al., 2010). NETs haben also nicht nur eine positive Seite.
Eine zu starke NETs-Bildung kann dem Organismus Schaden zuführen. Ein weite-
res Beispiel hierfür ist die Zystische Fibrose (CF), eine erbliche Erkrankung, die zur
Produktion von zähflüssigem Schleim und rezidivierenden bakteriellen Lungenin-
fektionen führt. Für die hohe Viskosität des Schleims werden darin enthaltene NETs
verantwortlich gemacht (Manzenreiter et al., 2012; Papayannopoulos, Staab &
Zychlinsky, 2011). Aus diesem Grund erzielt man durch die Behandlung der CF-
Patienten mit DNase eine Verflüssigung des Schleims (Manzenreiter et al., 2012)
und damit die Milderung der Symptomatik. Auch in der Blutgerinnung, der Koagula-
tion, spielen NETs eine Rolle, die fatale Folgen haben kann. Die für die primäre
Blutgerinnung verantwortlichen Thrombozyten induzieren eine zügige Bildung von
NETs (Clark et al., 2007). Andersherum führen NETs zu einer Anlagerung von
Thrombozyten und einer Aktivierung der Blutgerinnung (von Bruhl et al., 2012).
Nimmt die Koagulation Überhand, kommt es zur Verstopfung von Blutgefäßen.
NETs werden daher zum Beispiel auch für die Entstehung einer tiefen Beinven-
enthrombose (TVT) mitverantwortlich gemacht (Fuchs, Brill & Wagner, 2012; von
Bruhl et al., 2012). Im Mausmodell konnte bereits gezeigt werden, dass sich die
Verabreichung von DNase I protektiv auf eine TVT auswirkt (Brill et al., 2012), was
die Bedeutung von Serumnukleasen noch einmal unterstreicht.
In einer Sepsis stehen NETs außerdem im Verdacht, an der Entstehung eines
(Multi-)Organversagens beteiligt zu sein, indem sie eine Minderdurchblutung und
Zerstörung des Leberparenchyms hervorrufen (Clark et al., 2007). Somit könnten
12Einleitung
sie mit der hohen Sterblichkeit während der immunsuppressiven Phase im Zusam-
menhang stehen.
Durch die Degradation durch Nukleasen verlieren die NETs nicht nur ihre Struktur
(Brinkmann et al., 2004), sondern auch ihre antimikrobielle Aktivität (Fuchs et al.,
2007; Guimarães-Costa et al., 2009). Einige Bakterien, wie Gruppe A-Streptokok-
ken, machen sich diesen Mechanismus zu Nutze, indem sie eine potente DNase
(Sda1) produzieren, um dem Einfang durch NETs und ihrer Toxizität zu entkommen
(Buchanan et al., 2006). Dies könnte einen wichtigen Beitrag zu ihrer hohen Virulenz
leisten und erklären, wie sie lebensbedrohliche Erkrankungen, wie die nekrotisie-
rende Fasziitis oder das Streptococcal toxic shock syndrome (STSS), auslösen kön-
nen (Buchanan et al., 2006).
3.5 Zielsetzung
Die Studienlage zur Bildung von NETs durch neutrophile Granulozyten während ei-
ner Sepsis ist bislang noch lückenhaft. Eine vermehrte NETs-Bildung in der frühen
Phase der Sepsis ist wahrscheinlich, da das Immunsystem insgesamt stark aktiviert
ist. Um zu untersuchen ob auch noch andere immunaktivierende Ereignisse, wie
große Operationen, einen Einfluss auf die NETs-Bildung aufweisen, untersuchten
wir nicht nur die Bildung von NETs bei septischen Patienten, sondern auch bei Pa-
tienten nach großen viszeralchirurgischen Eingriffen.
Die NETs-induzierende Wirkung von PMA auf neutrophile Granulozyten von septi-
schen Mäusen in vitro wurde bereits beweisen (Meng et al., 2012). Die in diesem
Modell verwendeten Neutrophilen wurden jedoch aus dem Knochenmark von sep-
tischen Mäusen isoliert. Es bleibt damit offen, ob diese sich so verhalten wie hu-
mane neutrophile Granulozyten und anders auf die PMA-Stimulation reagieren, als
Neutrophile isoliert aus der Blutzirkulation, welche also die tatsächlich aktiven
Neutrophilen in einer Sepsis repräsentieren.
Bekannt ist bereits, dass vitale Staphylokokkus aureus (S. aureus) eine Induktion
der NETs-Bildung bewirken (Fuchs et al., 2007). Unklar ist bislang noch, worauf
diese NETs-Induktion beruht. Ein Ziel der Arbeit ist es zu prüfen, ob die NETs-Bil-
dung durch Oberflächenproteine von S. aureus hervorgerufen wird.
13Einleitung
Die DNA von Bakterien enthält eine deutlich höhere Anzahl an CpG (Cytosine-Gu-
anosin) Dinukleotidmotiven als die DNA von Säugetieren. Synthetisch hergestellte
CpG Oligonukleotide (ODN), die eine hohe Anzahl an CpG Dinukleotiden enthalten,
werden daher verwendet, um die Wirkung bakterieller DNA zu repräsentieren (ODN
2006, Invivogen, San Diego,USA). CpGs wirken über den TLR-9-Rezeptor und füh-
ren zu einer Aktivierung von neutrophilen Granulozyten (Zimmermann et al., 2015).
Ob sie auch zu einer NETs-Ausschleusung führen, ist bislang noch unbekannt.
Mitochondrien sind Zellorganellen mit einer eigenen DNA. Mitochondriale DNA
(mtDNA) wird daher bei der Schädigung von Zellen frei. In einer Sepsis kommt es
zum vermehrten Zelluntergang und auch zu erhöhten Serumkonzentrationen an
mtDNA, welche sich sogar als relevanter prognostischer Faktor herausgestellt hat
(Kung et al., 2012). Laut der Endosymbiontentheorie sind Mitochondrien aus proka-
ryotischen Vorläuferzellen entstanden, die phagozytiert wurden (Müller et al., 2012).
Zu den Prokaryoten zählen auch Bakterien. In mtDNA ist aufgrund ihres Ursprungs
eine hohe Anzahl an CpG Dinukleotiden enthalten (Cardon et al., 1994). Ihre proin-
flammatorische Wirkung ist bereits bekannt (Zhang et al., 2010). Ob sich mtDNA
auf die Ausbildung von NETs auswirkt, ist bislang noch nicht geklärt.
Serumnukleasen bauen NETs in der Zirkulation ab. Im Mausmodell konnte bereits
gezeigt werden, dass mit einer Erhöhung der zirkulierenden freien DNA (cfDNA)-
Spiegel auch eine erhöhte Serumkonzentration an Nuklease einher geht (Meng et
al., 2012). Unklar ist hingegen, wie NETs bei Patienten mit Sepsis reguliert sind.
Daher testeten wir nachfolgende Hypothesen:
1) Die NETs-Bildung von neutrophilen Granulozyten septischer Patienten ist in
vitro im Vergleich zu gesunden Probanden erhöht.
2) Auch chirurgische Eingriffe führen zu einer Induktion der NETs-Bildung.
3) Neutrophile sind auch bei Patienten mit Sepsis im Stande, NETs durch Sti-
mulation mit PMA in vitro zu bilden.
4) Auch hitzeinaktivierte Staphylokokken führen zu einer Induktion der NETs-
Bildung.
5) CPG und mtDNA haben Einfluss auf die Bildung von NETs in vitro.
6) Die Aktivität der Serumnukleasen ist bei septischen und postoperativen Pa-
tienten im Vergleich zu gesunden Probanden erhöht.
14Material und Methoden
4 Material und Methoden
4.1 Patienten und Probandendaten
Nach positivem Votum der Ethikkommission (Nr.: 09-4154, Universität Duisburg-
Essen, Essen, Deutschland) und Studienregistrierung (German clinical trials data-
base, DRKS no. 00007694) wurde eine prospektive Beobachtungsstudie eingelei-
tet, in die sowohl gesunde Probanden als auch Patienten nach großen chirurgischen
Eingriffen und Patienten mit Sepsis aufgenommen wurden.
Die gesunden Probanden wurden kliniksintern innerhalb eines Zeitraums vom
07.09.2012 bis zum 27.02.2015 rekrutiert. Sie erfüllten folgende Kriterien:
- Es besteht keine akute Infektion und keine bekannte therapiebedürftige Vor-
erkrankung, sowie kein Status nach einer Impfung innerhalb der letzten 14
Tage
- Nichtraucherstatus und kein exzessiver Nikotinkonsum in der Anamnese
- Keine Dauereinnahme von Medikamenten (bei Frauen wurde von der Ein-
nahme oraler Kontrazeptiva abgesehen)
- Volljährigkeit
Für die postoperativen Patienten mussten folgende Kriterien erfüllt sein:
- Durchführung eines großen abdominellen Eingriffs innerhalb der letzten 24
Stunden
- Kein Einsatz einer Herzlungenmaschine während der Operation
- Keine Chemotherapie in der Anamnese
- Kein bekanntes metastasiertes Tumorleiden
- Keine vorliegende Immunsuppression
Außerdem wurden septische Patienten eingeschlossen, die im Zeitraum vom
13.08.2012 bis 11.02.2015 auf einer Intensivstation des Universitätsklinikums Es-
sen behandelt wurden und die Diagnosekriterien einer Sepsis erfüllt haben, entspre-
chend den 2012 festgelegten Leitlinien der Surviving Sepsis Campaign (Dellinger
et al., 2013):
15Material und Methoden
Nachweis oder Annahme einer Infektion
Diagnose einer Infektion über den mikrobiologischen Nachweis oder durch klini-
sche Kriterien und einige der folgenden Kriterien:
a. Allgemeine Variablen
- Fieber (> 38,3 °C) oder Hypothermie (≤ 36 °C)
- Tachykardie: Herzfrequenz ≥ 90 min-1
- Tachypnoe
- Vigilanzminderung
- Signifikante Ödeme oder positive Flüssigkeitsbilanz (>20ml kg-1 über 24h)
- Hyperglykämie (>140 mg dl-1 oder 7,7mmol l-1) ohne Diabetes mellitus
b. Inflammatorische Variablen
- Leukozytose (≥ 12.000 weiße Blutkörperchen µl-1) oder Leukopenie (≤ 4.000
µl-1)
- ≥ 10 % unreife neutrophile Granulozyten im Differentialblutbild
- CRP-Konzentration im Plasma mehr als zwei Standardabweichungen über
dem Normwert
- PCT-Konzentration im Plasma mehr als zwei Standardabweichungen über
dem Normwert
c. Hämodynamische Variablen
- Arterielle Hypotension (systemischer Blutdruck < 90 mmHg, mittlerer arteri-
eller Druck < 70 mmHg oder Abfall > 40 mmHg bei Erwachsenen oder weni-
ger als zwei Standardabweichungen unter dem Altersnormwert
d. Variablen einer Organdysfunktion
- Arterielle Hypoxämie (paO2/fiO2 < 300)
- Akute Oligurie (Diurese < 0,5 ml kg-1 h-1) für mindestens zwei Stunden trotz
adäquater Flüssigkeitstherapie)
- Kreatininanstieg > 0,5 mg dl-1 oder 44,2 µmol l-1)
- Koagulopathie (INR > 1,5 oder aPTT > 60 s)
- Ileus (fehlende Darmgeräusche)
- Thrombozytopenie (< 100.000 µl-1)
- Hyperbilirubinämie (Plasma Bilirubin > 4 mg dl-1 oder 70 µmol l-1)
e. Variablen der Gewebedurchblutung
- Laktatanstieg (> 1 mmol l-1)
- Verlängerte Rekapillarisierungszeit oder Marmorierung
16Material und Methoden
Anzumerken ist, dass der Einschluss der Patienten in unsere Studie nach den zum
Zeitpunkt der Studie gültigen Sepsiskriterien erfolgte. Diese wurden in der Zwi-
schenzeit modifiziert (Singer et al., 2016) und daher können wir unsere Ergebnisse
nicht 1:1 auf Patienten übertragen, die nach der neuen Definition eine Sepsis auf-
weisen.
Während der oben genannten Zeiträume konnten 31 gesunde Probanden, 8 posto-
perative Patienten sowie 21 Patienten mit Sepsis rekrutiert werden. Alle Probanden,
Patienten bzw. deren gesetzliche Betreuer haben schriftlich ihr Einverständnis zur
Teilnahme an der Studie erklärt. Zusätzlich wurden Patientencharakteristika wie Al-
ter, Geschlecht, BMI, Hauptdiagnosen und der Krankheitsverlauf (Krankenhausauf-
enthaltsdauer, Mortalität) dokumentiert und in eine Datenbank aufgenommen. Mik-
robiologisch gesicherte Angaben über Keime sowie hämodynamische und labor-
chemische Variablen, wie die Entzündungsvariablen Procalzitonin (PCT), C-reakti-
ves-Protein (CRP) und Leukozyten, wurden ebenfalls mit in die Datenbank aufge-
nommen. Außerdem wurde der intensivmedizinische Komplexbehandlungsscore
Simplified Acute Physiology Score II (SAPS II) berechnet und dokumentiert. Inner-
halb von 24 Stunden nach Diagnosestellung der Sepsis, bzw. Durchführung des
chirurgischen Eingriffs, wurde den Patienten 20 ml Vollblut entnommen, das für die
weiteren Versuche umgehend verarbeitet wurde.
4.2 Blutentnahme
Die Isolation der neutrophilen Granulozyten erfolgte aus 20 ml venösem oder arte-
riellem Blut in Lithium-Heparin-Monovetten (S-Monovette® 9 ml, Sarstedt AG & Co.,
Nümbrecht, Deutschland). Bei den septischen sowie postoperativen Patienten er-
folgte die Blutentnahme bei Vorliegen eines arteriellen Zugangs vorzugsweise arte-
riell, alternativ über einen zentralen Venenkatheter. Die arterielle Blutentnahme
wurde bevorzugt, um eine mögliche Verfälschung der Neutrophilenaktivität durch
infundierte Medikamente zu vermeiden. Von einer erneuten peripheren Venenpunk-
tion sollte abgesehen werden. Sämtlichen gesunden Probanden wurde über eine
einmalige frische periphere Venenpunktion Blut entnommen. Die Heparin-Monovet-
ten wurden äußerst behutsam befüllt, um eine Verfälschung des Ergebnisses durch
17Material und Methoden
Voraktivierung der Neutrophilen, wie durch sehr schnellen Blutfluss unter hohem
Druck oder Einschluss von Luftbläschen, zu vermeiden.
Da die Neutrophilen entweder aus arteriellem oder aus venösem Blut stammten,
musste zunächst ausgeschlossen werden, dass der Zugang zur Blutentnahme ei-
nen Einfluss auf die NETs-Bildung besitzt. Bei einigen postoperativen und septi-
schen Patienten erfolgte zeitgleich die Entnahme über beide Zugangswege und der
Vergleich der gebildeten NETs-Menge (N=9).
4.3 Isolation neutrophiler Granulozyten aus
humanem Vollblut mittels Dichtegradien-
tenzentrifugation
Für die Isolation der neutrophilen Granulozyten wurden zunächst 20 ml Polymorph-
prep (Progen, Heidelberg, Deutschland) in ein steriles 50 ml Falcontube (BD
Biosciences, Erembodegem, Belgien) vorgelegt und dieses behutsam mit 20 ml
Vollblut mittels einer Pipettierhilfe (Hirschmann Laborgeräte, Eberstadt, Deutsch-
land) überschichtet, so dass eine Vermischung der beiden sich ergebenden Schich-
ten vermieden wurde. Das Falcontube wurde daraufhin für 30 Minuten bei 512 x g
und Raumtemperatur (22 °C) ohne Bremse zentrifugiert. Nach der Zentrifugation
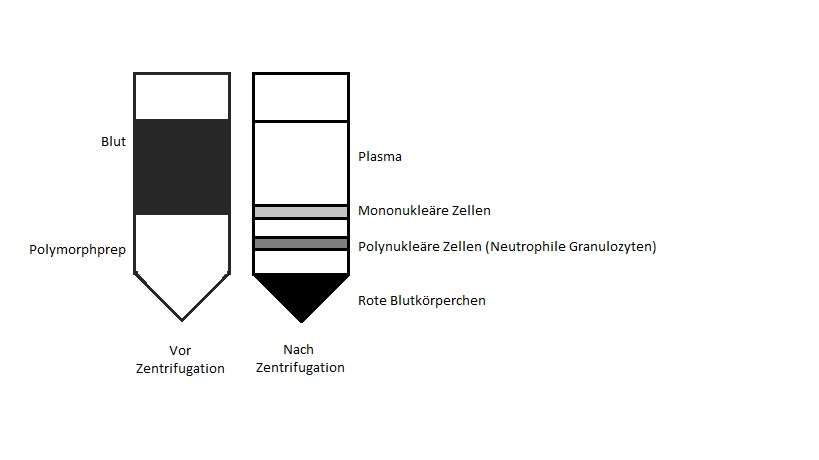
lagen die Zellen in dem Falcontube, wie in Abbildung 4.3.1 dargestellt, in drei von-
einander getrennten Schichten vor:
18Material und Methoden
Abb. 4.3.1 Isolation von neutrophilen Granulozyten mittels Dichtegradientenzen-
trifugation:
Vor Zentrifugation liegen zwei Phasen vor, in der unteren Polymorphprep, in der oberen
frisch entnommenes Vollblut. Nach der Zentrifugation liegen die verschiedenen Zellarten in
drei Schichten vor: In der oberen die mononukleären Zellen, in der mittleren die neutrophi-
len Granulozyten und in der unteren die Erythrozyten.
19Material und Methoden
Nach der Zentrifugation wurden zunächst das Blutplasma und die mononukleären
Zellen aus den oberen Schichten mit einer Pipette abgenommen und verworfen.
Anschließend wurde die mittlere Zellschicht aus neutrophilen Granulozyten mit ei-
ner Pipette in ein neues 50 ml Falcontube überführt und mit Phosphate Buffered
Saline (PBS) (Gibco, Life Technologies, Darmstadt, Deutschland) auf 50 ml aufge-
füllt. Es erfolgte ein weiterer zehnminütiger Zentrifugationsschritt mit Bremse bei
512 x g und 22 °C, nach dem sich ein Sediment aus neutrophilen Granulozyten und
einigen verbliebenen Erythrozyten am Boden des Falcons absetzte. Der Überstand
wurde verworfen und das Sediment in 5 ml sterilem Wasser (Merck Millipore, Darm-
stadt, Deutschland) resuspendiert und somit die vorhandenen Erythrozyten lysiert.
Nach ca. fünf Sekunden wurde das Falcontube, zum Stoppen der Lyse und zur Ver-
hinderung einer Schädigung der neutrophilen Granulozyten, zügig mit 45 ml PBS
aufgefüllt. Anschließend fand ein weiterer zehnminütiger Zentrifugationsschritt bei
512 x g und Raumtemperatur statt. Nach Beurteilung der Farbe des Zellsediments
fand ggf. ein erneuter Schritt zur Lyse von noch vorhandenen Erythrozyten sowie
eine Wiederholung des letzten Zentrifugationsschrittes statt, wobei maximal zwei-
mal lysiert wurde. Daraufhin wurde der Überstand abgenommen und die neutrophi-
len Granulozyten wurden in 1 ml Zellkulturmedium (RPMI-1640 ohne Zusätze,
Gibco, Life Technologies, Darmstadt, Deutschland) resuspendiert. Zur Beurteilung
der Zellvitalität sowie zur Bestimmung der Zellzahl wurden 10 µl der Zellsuspension
mit 90 µl 0,4-prozentiger Trypanblau-Lösung (Thermo Fisher Scientific Inc.,
Schwerte, Deutschland) gemischt, in eine Neubauer Zählkammer (LO-Laboroptik
GmbH, Friedrichsdorf, Deutschland) gegeben und anschließend unter einem Licht-
mikroskop (Axio Vert.A1, Zeiss, Hamburg, Deutschland) ausgezählt. Die Zellzahl
wurde mit RPMI-Medium auf zwei Millionen Zellen pro ml eingestellt.
Sämtliche Arbeiten fanden unter einer sterilen Werkbank (Thermo Fisher Scientific
Inc., Schwerte, Deutschland) statt, um jegliche Kontamination der Zellkultur zu ver-
hindern. Bei allen Pipettierschritten wurde darauf geachtet, dass diese behutsam
durchgeführt wurden, um eine Vorstimulation und somit ggf. Verfälschung der Er-
gebnisse zu vermeiden.
20Material und Methoden
4.4 Hitzeinaktivierung der Staphylokokkus
aureus
Für die Abtötung der Staphylokokkus aureus (S. aureus) wurden 5 ml BHI (Brain
Heart Infusion Broth) mit einer Bakterienkolonie (S. aureus USA 300 LAC AH 1263
(Wt)) beimpft und über Nacht bei 37 °C und 200 rpm inkubiert. Die Bakterien wurden
anschließend 1 : 100 in BHI verdünnt (1 ml Bakterien + 99 ml BHI) und wieder bei
37 °C und 200 rpm inkubiert bis zu einem OD600 von 0,7. 45 ml Bakterienlösung
wurden in ein 50 ml Falcon überführt und zur Hitzeinaktivierung 30 Minuten lang in
ein 95 °C heißes Wasserbad gegeben. Durch Aussähen von Bakterien auf einer
Blutagarplatte über Nacht wurde die erfolgreiche Abtötung überprüft. Die restlichen
Bakterien wurden in 1,5 ml Eppendorf Tubes (Eppendorf AG, Hamburg, Deutsch-
land) pipettiert und bis zur weiteren Verwendung bei -20 °C gelagert.
4.5 Zellkultur zur Isolation von Mitochond-
rien
Um größere Mengen mitochondrialer DNA zu gewinnen, wurde die humane Zelllinie
HepG2 verwendet. Es handelt sich dabei um hepatozelluläre Karzinomzellen, die in
vitro einen hohen Grad an funktionaler und morphologischer Differenzierung auf-
weisen. Die Kultivierung der Zellen erfolgte im Inkubationsschrank bei 37 °C und 5
% CO2 in RPMI-Medium, das mit 10 % fetalem Kälberserum (FKS) und 1 mM Nat-
riumpyruvat (Thermo Fisher Scientific Inc., Schwerte, Deutschland) versetzt war. Je
nach Konfluenz wurden die Zellen nach sechs bis sieben Tagen passagiert. Dazu
wurde zunächst das Medium abgesaugt und die Zellen mit 10 ml PBS gewaschen.
Zur Ablösung der Zellen vom Boden der Kulturflasche wurden 10 ml TrypLE Ex-
press (Thermo Fisher Scientific Inc., Schwerte, Deutschland) hinzugegeben und die
Zellen dann für 20 Minuten bei 37 °C inkubiert. Nach Ablauf der Inkubationszeit
konnten die Zellen durch Schwenken der Zellkulturflasche vom Boden gelöst wer-
den. Die so erhaltene Zellsuspension wurde in 50 ml Falcontubes überführt und für
sieben Minuten bei 700 x g und 22 °C zentrifugiert. Das abzentrifugierte Zellsedi-
ment wurde resuspendiert in 20 ml frischem Zellkulturmedium und gleichmäßig auf
21Material und Methoden
vier neue Zellkulturflaschen (TPP Techno Plastic Products AG, Trasadingen,
Schweiz) aufgeteilt.
Das Ernten der Zellen erfolgte erst bei maximaler Konfluenz. Dafür wurden die Zel-
len ebenfalls mit PBS gewaschen, mit TrypLE Express gelöst und abzentrifugiert.
Die in der Zellsuspension enthaltene Zellzahl wurde während der Zentrifugationszeit
in einer Neubauer Zählkammer ausgezählt. Nach der Zentrifugation wurde das Me-
dium, welches den Überstand bildete, verworfen und das Zellsediment in einer pas-
senden Menge an Bambanker-Medium (Nippon Genetics Europe GmbH, Düren,
Deutschland) resuspendiert, so dass sich eine Zellzahl von 1 x 107 Zellen in der
Endlösung ergab. Diese Zellen wurden bei -80 °C in 2 ml Aliquots eingefroren, um
aus ihnen im Verlauf mitochondriale DNA zu isolieren.
4.6 Isolation von mitochondrialer DNA aus
kultivierten HepG2-Zellen
Für einen Ansatz wurden die Zellen aus einem 2 ml Aliquot eingesetzt, entspre-
chend einer Zellzahl von 2 x 107 Zellen. Die bei -80 °C gelagerte Zellsuspension
wurde zunächst bei 37 °C im Wasserbad aufgetaut, in neue 2 ml Eppendorf Tubes
überführt und sofort weiterverarbeitet. Die Isolation der Mitochondrien aus den kul-
tivierten HepG2-Zellen wurde mittels Mitochondria Isolation Kit for Cultured Cells
(Thermo Fisher Scientific Inc., Schwerte, Deutschland) entsprechend den Herstel-
lerangaben durchgeführt. Die Arbeits- und Inkubationsschritte wurden auf Eis durch-
geführt. Zunächst wurden die 2 x 107 Zellen durch zweiminütige Zentrifugation bei
850 x g pelletiert und der Überstand verworfen. Hinzugefügt wurden 800 µl Mito-
chondria Isolation Reagent A. Nach genau zwei Minuten Inkubationszeit wurden 5
µl Mitochondria Isolation Reagent B dazu gegeben und fünf Minuten lang inkubiert.
Es wurden 800 µl Reagent C hinzu pipettiert und das Tube zehn Minuten lang bei
700 x g und 4 °C zentrifugiert. Der Überstand wurde in ein neues Falcontube über-
führt und bei 3.000 x g 15 Minuten lang zentrifugiert. Zu dem gebildeten Pellet wur-
den 500 µl Reagent C gegeben und es erfolgte ein weiterer fünfminütiger Zentrifu-
gationsschritt bei 12.000 x g. Nach Abnahme des Überstandes verblieb ein Mito-
chondrien-Sediment.
22Material und Methoden
Zur Isolation der DNA aus den aufgereinigten Mitochondrien wurde das DNeasy
Blood & Tissue Kit (QIAGEN GmbH, Hilden, Deutschland) verwendet. Die Isolation
erfolgte entsprechend den Herstelleranweisungen für kultivierte Zellen. Wenn die
mtDNA nicht direkt weiter isoliert werden konnte, mussten die Mitochondrien für
eine Lagerung zunächst aufgebrochen werden. Dazu wurden die ersten zwei Ar-
beitsschritte der mtDNA-Isolation durchgeführt. Die Mitochondrien-Pellets wurden
resuspendiert in 200 µl PBS. Es wurde 20 µl Proteinase K hinzugegeben, sowie 200
µl Puffer AL. Die Suspension wurde durch vortexen gemischt und für zehn Minuten
bei 56 °C inkubiert. Anschließend konnten die Mitochondrien zur weiteren DNA-Iso-
lation bei -20 °C eingefroren werden. Nachdem die Mitochondriensuspension auf-
getaut war, wurden 200 µl 100 %-Ethanol zugefügt. Es folgten zwei Zentrifugations-
schritte bei 6.000 x g mit zwischenzeitlicher Zugabe von 500 µl Puffer AW1 und
AW2. Die Trocknung der DNeasy Membran erfolgte durch Zentrifugation bei 20.000
x g. Schließlich wurde die isolierte mtDNA in Puffer AE aufgenommen. Zur Konzent-
rationsbestimmung der gewonnenen mtDNA wurde die optische Dichte bei 280 nm
Wellenlänge photometrisch detektiert (Biophotometer Plus, Eppendorf, Hamburg,
Deutschland). Die mtDNA wurde bis zu ihrer Verwendung für die Stimulation neutro-
philer Granulozyten bei -20 °C gelagert.
4.7 Stimulation der NETs-Bildung
Zunächst wurden zwei Zellkulturplatten mit jeweils 24 Wells (TPP Techno Plastic
Products AG, Trasadingen, Schweiz) vorbereitet, indem sterile Deckgläschen
(Thermo Fisher Scientific Inc., Schwerte, Deutschland) mit einem Durchmesser von
12 mm auf dem Boden der Wells ausgelegt wurden. Diese wurden beschichtet mit
jeweils 100 µl 0,01-prozentigem Poly-L-Lysin (Sigma-Aldrich, München, Deutsch-
land), das mittig auf die Deckgläschen gegeben wurde. Nach 20 Minuten Inkubati-
onszeit wurde das Poly-L-Lysin wieder von den Deckgläschen abpipettiert und es
folgten zwei Waschschritte mit jeweils 500 µl PBS-Puffer. Danach wurden in jedes
Well 250 µl der Neutrophilen-Zellsuspension, entsprechend einer Zellzahl von 5 x
10⁵ Zellen, gegeben. Hinzu kamen in jedes Well je nach Probe verschiedene zuvor
vorbereitete Lösungen, deren Effekt auf die Bildung von NETs getestet werden
sollte (siehe Tabelle 4.7.1). Als Positivkontrolle diente dabei, wie empfohlen von
23Material und Methoden
Brinkmann et al. (Brinkmann et al., 2010), die Inkubation mit 250 µl Phorbol 12-
Myristate 13-Acetate (PMA)-Lösung (Sigma-Aldrich, St. Louis, USA) in RPMI, des-
sen starke NETs-induzierende Wirkung bereits bekannt ist. 250 µl RPMI-Medium
dienten als Negativkontrolle und entsprechen der NETs-Bildung unter basalen Um-
ständen in unserer Versuchsreihe. Als avitale S. aureus verwendeten wir die, wie
zuvor beschrieben, hitzeinaktivierten S. aureus (USA 300 LAC AH 1263 (Wt)). Als
CpG wurden CpG ODN 2006, Type B Cpg oligonucleotide-Human TLR9 ligand (In-
vivogen, San Diego, USA) eingesetzt. Die mtDNA wurde nach zuvor beschriebenen
Protokollen isoliert.
Die Zellkulturplatten wurden für fünf Minuten bei 512 x g und Raumtemperatur zen-
trifugiert, um eine Adhäsion der neutrophilen Granulozyten an den Deckgläschen
zu bewirken. Anschließend wurde im Inkubationsschrank bei 37 °C und 5 % CO₂
eine der Zellkulturplatten zwei Stunden lang, die andere vier Stunden lang inkubiert.
Nach Ablauf dieser Inkubationszeiten wurde der Überstand in den Wells belassen
und in jedes Well 150 µl 16-prozentige Paraformaldehyd (PFA)-Lösung (Electron
Microscopy Sciences, München, Deutschland) zum Fixieren der Zellen hinzugege-
ben. Die 24-Wellplatten wurden sorgfältig mit Parafilm M (Bemis, Braine L'Alleud,
Belgien) umwickelt, um eine Evaporation der Flüssigkeit und Austrocknung der
Neutrophilen zu verhindern. Die Aufbewahrung der Zellkulturplatten erfolgte bis zur
weiteren Bearbeitung im Kühlschrank bei 4 °C.
Tabelle 4.7.1: Vorgelegte Lösungen zur Stimulation der neutrophilen Gra-
nulozyten
Probe Vorgelegte Lösung
Negativkontrolle 250 µl RPMI-Medium
Positivkontrolle (PMA) 250 µl 50 nM PMA-Lösung in RPMI (Endkonzentra-
tion: 25nM)
Hitzeinaktivierter Staphy- 250 µl Staphylokokkus aureus in PBS (4 x 10⁶ Bakte-
lokokkus aureus rien ml-1)
CpG 250 µl CpG-Lösung (20 µg ml-1)
mtDNA 250 µl mtDNA-Lösung in RPMI (20 µg ml-1)
Stimulation mit Phorbol 12-Myristate 13-Acetate (PMA), CpG oligonucleotide-Human TLR9
ligand (CpG) und mitochondrialer DNA (mtDNA).
24Material und Methoden
4.8 NETs-Visualisierung mittels Immunfluo-
reszenz
Die auf den Deckgläschen fixierten Zellen wurden dreimal mit je 500 µl PBS gewa-
schen. Sie wurden permeabilisiert und durch 45-minütige Inkubation mit 200 µl 2 %
BSA in 0.2 % Triton X-100/PBS bei Raumtemperatur geblockt. Nach Absaugen des
Blockingpuffers erfolgte die Zugabe von 200 µl des Primärantikörpers, ein monoklo-
naler Maus anti-H2A-H2B-DNA Komplex (clone PL2-6, 0.5 µg ml-1 in 2 % BSA in
0.2 % Triton X-100/PBS) und die Inkubation über Nacht bei 4 °C. Nach drei weiteren
Waschschritten mit PBS wurde ein Alexa-488-gekoppelter Sekundärantikörper
(goat anti mouse Alexa 488, Thermo Fisher Scientific Inc., Schwerte, Deutschland)
in Blockingpuffer hinzugegeben und 45 Minuten bei Raumtemperatur ohne Lichtex-
position inkubiert. Vier weitere Waschschritte mit PBS folgten. Auf einen Objektträ-
ger wurden jeweils zwei Tropfen ProlongGold mit 4′,6′-diamidino-2-phenylindole
(DAPI) (Invitrogen, Carlsbad, USA) gegeben und die Deckgläschen umgedreht auf
je einen Tropfen ProlongGold-DAPI gebettet, so dass sich die angehefteten Zellen
auf der Unterseite befanden. Die Objektträger wurden über Nacht vollständig trock-
nen gelassen und am nächsten Morgen mit klarem Nagellack versiegelt.
4.9 Mikroskopische Beurteilung der NETs
Die Beurteilung der Proben wurde in Kooperation mit dem Institut für Physiologische
Chemie der TIHO Hannover unter Leitung von Frau PD Dr. Maren von Köckritz-
Blickwede mittels eines konfokalen Fluoreszenzmikroskops (Leica TCS SP5) sowie
einem nichtkonfokalen Fluoreszenzmikroskop (Zeiss Anxioveit 200M) durchgeführt.
Von jedem Deckgläschen wurden bei 40-facher Vergrößerung drei Aufnahmen von
zufällig ausgewählten Bildausschnitten gemacht und im TIF-Format gespeichert.
Zur Quantifizierung der NETs-bildenden Zellen wurde die Software ImageJ (Version
1.48, National Institutes of Health, USA) verwendet. Der Bewertung der NETs-Bil-
dung lagen folgende Kriterien zugrunde, von denen mindestens eins erfüllt sein
musste:
25Material und Methoden
Es galten alle Zellen laut folgender Primärkriterien als NETs-positiv, die
1a) ihre eigene DNA ausgeschleust haben oder
1b) ausgeschleuste DNA berühren;
oder wenn keines dieser Kriterien zutrifft, aber eines der weiteren Sekundärkriterien:
2a) die Zellen dekondensiertes Kernmaterial aufweisen;
2b) ihr Zellkern im Durchmesser zugenommen hat oder
2c) ihr Zellkern seine runde Form verloren hat
(Brinkmann et al., 2010; Brogden et al., 2017; von Köckritz-Blickwede, 2010).
Die Sekundärkriterien beruhen auf mikroskopischen und lebend-Zell-Imaging Be-
obachtungen, dass es nach Stimulation der Neutrophilen zur Formänderung des
Zellkerns kommt und Eu- und Heterochromatin verschmilzen (Brinkmann et al.,
2010). Im Verlauf kommt es zum Aufbrechen des Zellkerns und der Granuolen und
die Bestandteile der NETs können sich mischen. Erst danach werden die NETs aus
den Zellen ausgeschleust und die Zellmembran bricht.
In den Abbildungen 4.9.1 und 4.9.2 sind die NETs-Kriterien zur Veranschaulichung
anhand einiger Beispiele dargestellt.
26Material und Methoden
Abb. 4.9.1 Beispielbild 1 zu den Kriterien NET-positiver Zellen
Die grün markierten Zellen gelten als NET-negativ. Die gelb markierte Zelle stellt eine akti-
vierte, jedoch ebenfalls NET-negative Form dar. Die rot markierten Zellen sind NET-positiv
a) anhand der Primärkriterien, da sie ausgeschleuste DNA berühren und b) anhand der
Sekundärkriterien, da sie einen vergrößerten oder dekondensierten Zellkern aufweisen.
27Material und Methoden
Abb. 4.9.2 Beispielbild 2 zu den Kriterien NET-positiver Zellen
Die grün markierten Zellen sind NET-negativ. Die gelb markierte Zelle stellt eine aktivierte,
NET-negative Form dar. Die rot markierten Zellen gelten als NET-positiv a) anhand der
Primärkriterien und b) anhand der Sekundärkriterien.
Es wurde das prozentuale Verhältnis der NET-positiven Zellen zu allen im Bildaus-
schnitt vorhandenen Zellen gebildet. Da es von jedem Versuchsansatz zwei Deck-
gläschen gab und je drei Bilder gemacht wurden, entstanden je Versuchsansatz
sechs Bilder, aus denen der Mittelwert errechnet wurde. Dieser Mittelwert wurde für
die weitere statistische Analyse verwendet.
28Material und Methoden
4.10 Bestimmung der Serumnukleaseaktivität
Die Quantifizierung der Serumnukleaseaktivität erfolgte mittels Gelelektrophorese.
Die Serumproben waren zuvor gesammelt und bei -80 °C gelagert worden. Nach
dem Auftauen der Proben bei Raumtemperatur wurden je 10 µl Serum mit 40 µl
Tris(hydroxymethyl)-aminomethan (TRIS)-Puffer (300 mM TRIS, 50 mM Calcium-
chlorid, 50 mM Magnesiumchlorid) versetzt. Als Negativkontrolle dienten 50 µl des
TRIS-Puffers, als Positivkontrolle eine Verdünnungsreihe des Enzyms Desoxyribo-
nuklease I (DNAse I, Sigma-Aldrich, München, Deutschland) mit einem Aktivitäts-
bereich zwischen 2 und 0,0035 Units ml-1 (U ml-1). Sowohl zu den Serumproben als
auch zu der Negativ- und Positivkontrolle wurden 7,5 µl Kälberthymus-DNA (Sigma-
Aldrich, München, Deutschland) mit einer Konzentration von 1 mg ml-1 gegeben. Es
folgte eine 18-stündige Inkubation bei 37 °C. Anschließend wurde eine Phenol-Chlo-
roform-Extraktion durchgeführt, um die in den Proben enthaltene DNA von den Se-
rumproteinen zu trennen. Dafür wurden die Serumproben mit 50 µl Phenol-Chloro-
form (Carl Roth, Karlsruhe, Deutschland) versetzt und 30 Sekunden bei 15.000 x g
und Raumtemperatur zentrifugiert. Es bildete sich eine wässrige Phase, in der die
DNA enthalten ist. Von dieser wässrigen Phase wurden 16 µl mit 4 µl Ladepuffer
(Thermo Fisher Scientific, Schwerte, Deutschland) vermischt und in die Geltaschen
eines einprozentigen Agarosegels pipettiert. Danach wurde eine 30-minütige Gel-
elektrophorese bei 100 V Spannung durchgeführt.
Von den entstandenen Gelen wurden unter UV-Bestrahlung Bilder aufgenommen,
die zur Auswertung verwendet wurden. Die Bestimmung der DNAse I-Aktivität in
den Serumproben erfolgte semiquantitativ, durch den Vergleich der Gelspuren der
Proben mit den Gelspuren der DNase I-Verdünnungsreihe. Es wurden sechs Kate-
gorien bestimmt (Tabelle 4.10.1), denen die Serumnukleaseaktivität infolgedessen
zugeordnet werden konnte.
29Material und Methoden
Tabelle 4.10.1: Semiquantitative Einteilung der Serumnukleaseaktivität
Aktivitäts- Zuordnung zur DNAse I-Aktivität
kategorie
1 entspricht einer DNase I-Aktivität von 0 U ml-1
2 entspricht einer DNase I-Aktivität < 0,007 U ml-1
3 entspricht einer DNase I-Aktivität von 0,007 U ml-1
4 entspricht einer DNase I-Aktivität zwischen 0,007 und 0,015 U ml-1
5 entspricht einer DNase I-Aktivität zwischen 0,015 und 0,06 U ml-1
6 entspricht einer DNase I-Aktivität ≥ 0,06 U ml-1
Verdünnungsreihe des Enzyms Desoxyribonuklease I (DNase I), Units ml-1 (U ml-1)
Ein exemplarisches Gelbild ist in Abbildung 4.10.1 dargestellt.
Abb. 4.10.1 Serumnukleaseaktivität dargestellt mittels Gelelektrophorese
In diesem exemplarischen Gelbild sind 22 Serumproben aufgetragen. Links im Bild liegen
die Negativkontrolle (NC) und die Positivkontrolle (PC). Rechts befinden sich die DNase-
Verdünnungsreihen. Die Nukleaseaktivitätskategorie ist jeweils unter der zugehörigen Se-
rumprobe eingetragen. Je weiter unten im Bild sich die Serumprobe befindet, desto höher
ist die zugehörige Nukleaseaktivität.
30Sie können auch lesen

















































