Onkogene Effekte von CD19: Vergleichende Untersuchungen an CD19-Verlustvarianten im murinen Burkitt Lymphom - OPUS 4
←
→
Transkription von Seiteninhalten
Wenn Ihr Browser die Seite nicht korrekt rendert, bitte, lesen Sie den Inhalt der Seite unten
Abstract
Onkogene Effekte von CD19:
Vergleichende Untersuchungen an
CD19-Verlustvarianten
im murinen Burkitt Lymphom
Der Medizinischen Fakultät
der Friedrich-Alexander-Universität
Erlangen-Nürnberg
zur
Erlangung des Doktorgrades Dr. med.
vorgelegt von
Kim Veronique Delventhal
aus Hannover
0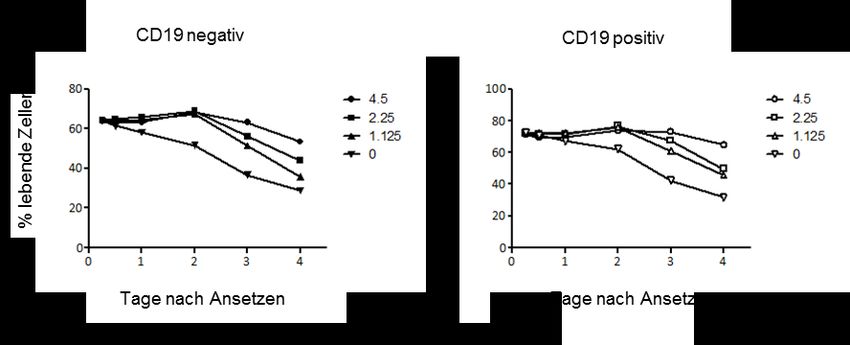
Als Dissertation genehmigt von der
Medizinischen Fakultät der Friedrich-Alexander-Universität
Erlangen-Nürnberg
Vorsitzender des Promotionsorgans: Prof. Dr. Markus F. Neurath
Gutachter: Prof. Dr. Andreas Mackensen
Gutachter: PD Dr. Heiko Bruns
Tag der mündlichen Prüfung: 26. Mai 2020
1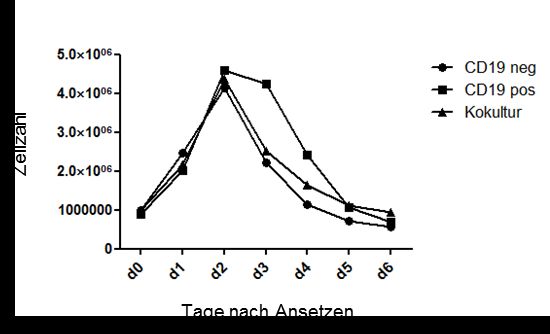
Abstract
Für meine Eltern
IInhaltsverzeichnis
Abkürzungsverzeichnis
Abstract .......................................................................................................................... 1
Zusammenfassung ......................................................................................................... 2
1. Einleitung .................................................................................................................... 3
1.1 Grundlegende Prinzipien des Immunsystems....................................................... 3
1.1.1 Zelluläre Bestandteile des Immunsystems ..................................................... 4
1.2 B-Lymphozyten: Entwicklung und Rezeptorstruktur ............................................. 5
1.2.1 Die Entwicklung von B-Lymphozyten ............................................................. 5
1.2.2 Mechanismen der Antigenerkennung durch B-Lymphozyten ......................... 6
1.2.3 Der B-Zell-Rezeptor-Komplex ........................................................................ 7
1.3 CD19 als spezifischer B-Zell-Marker und tumorassoziiertes Antigen ................... 9
1.3.1 Der Einfluss von CD19 auf die Signalvermittlung in B-Lymphozyten ............. 9
1.4 Lymphatische Neoplasien: Maligne Entartung von B-Lymphozyten ................... 12
1.4.1 Das Burkitt-Lymphom ................................................................................... 12
1.5 Konzept der Immuntherapie ................................................................................ 13
1.5.1 Aktueller Entwicklungsstand der Immuntherapie ......................................... 14
2. Zielsetzung der Arbeit............................................................................................... 16
3. Ergebnisse................................................................................................................ 17
3.1 Autoselektion CD19-positiver B-Lymphomzellen nach retroviraler Transduktion 17
3.2 CD19 als Selektionsvorteil in Ko-Kultur mit primär CD19-defizienten
B-Lymphomzellen ..................................................................................................... 20
3.2.1 Proliferationsvorteil CD19-positiver B-Lymphomzellen in Ko-Kultur mit
CD19-negativen Geschwisterzellen ...................................................................... 20
3.2.2 Untersuchungen der Proliferationsgeschwindigkeit in Ko-Kultur .................. 21
3.2.2.1 Zellteilungshäufigkeit ................................................................................. 21
3.2.2.2 Zellzyklusanalysen .................................................................................... 23
3.2.3 Einfluss von interzellulären Kontakten auf das Proliferationsverhalten in
Ko-Kultur ............................................................................................................... 25
3.2.4 Apoptose als Wirkfaktor in der Entstehung des Proliferationsunterschiedes in
Ko-Kultur ............................................................................................................... 27
3.3 Vergleichende Stoffwechselanalysen an CD19-negativen und CD19-positiven
B-Lymphomzelllinien in Einzelkultur ......................................................................... 29
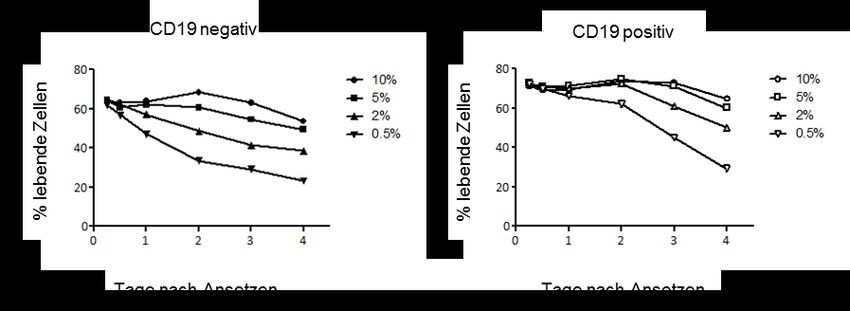
3.3.1 Verbessertes Überleben CD19-positiver B-Lymphomzellen unter
Hungerbedingungen .............................................................................................. 29
II3.3.2 Folgen des kontrollierten Entzugs einzelner Ernährungsfaktoren in
Einzelkultur ............................................................................................................ 30
4. Diskussion ................................................................................................................ 33
4.1 Unklare Rolle von CD19 als Überlebensvorteil für Lymphomzellen ................... 34
4.2 CD19 schützt nicht immer vor dem Tod, hilft aber bei Stress ............................. 37
4.3 Transwell: Alternative Übertragungswege von CD19 durch Membranbrücken .. 39
4.4 Ausblick............................................................................................................... 44
5. Material ..................................................................................................................... 45
5.1 Apparative Technik ............................................................................................. 45
5.2 Reagenzien und Materialien ............................................................................... 45
5.3 Kommerzielle Kits ............................................................................................... 46
5.4 Lösungen und Puffer........................................................................................... 46
5.5 Medien für Zellkultur und Zusätze....................................................................... 46
5.6 Zellkultur Medium................................................................................................ 47
5.7 Antikörper............................................................................................................ 47
5.8 Vitalitätsfarbstoffe ............................................................................................... 48
5.9 Vektoren.............................................................................................................. 49
5.10 Murine Zelllinien ................................................................................................ 49
5.11 Humane Zelllinien ............................................................................................. 49
6. Methoden.................................................................................................................. 50
6.1 Allgemeine Zellkulturtechnik ............................................................................... 50
6.1.1 Bestimmung der Zelldichte und Vitalität ....................................................... 50
6.1.2 Kryokonservierung von Zellen ...................................................................... 50
6.2 Durchflusszytometrische Analysen ..................................................................... 50
6.2.1 Oberflächenfärbungen.................................................................................. 50
6.2.2 Färbung mit Annexin V und 7AAD ............................................................... 51
6.2.3 VPD-Färbung ............................................................................................... 51
6.2.4 Färbung mit BrdU zur Zellzyklusanalyse ...................................................... 51
6.3 Transfektion und Transduktion ........................................................................... 52
6.3.1 Transfektion der Phoenix Zellen................................................................... 52
6.3.2 Transduktion der Lymphomzellen ................................................................ 52
6.4 Ko-Kulturansatz .................................................................................................. 53
6.4.1 Ko-Kultur mit Transwell ................................................................................ 53
6.5 Stoffwechselanalysen ......................................................................................... 54
6.5.1 Proliferationsverhalten.................................................................................. 54
6.5.2 Kontrollierter Entzug von Serum und Glucose ............................................. 54
IIILiteraturverzeichnis ...................................................................................................... 55
Abbildungsverzeichnis .................................................................................................. 60
Tabellenverzeichnis ...................................................................................................... 61
Danksagung ................................................................................................................. 62
Lebenslauf .................................................................................................................... 64
IVAbkürzungsverzeichnis
7AAD 7-Aminoactinomycin
Abl Abelson murine leukemia viral oncogene homolog 1
ALL Akute lymphatische Leukämie
APC Antigen-presenting-cell
B-Zelle Bursa derived cell
BCG Bacillus-Calmette-Guerin
BITE Bispezifische T-Zell-Engager
BrdU 5’-bromo-2’-deoxyuridine
Btk Bruton’s tyrosine kinase
BZR B-Zell-Rezeptor
CaCl2 Calciumchlorid
CART Chimeric Antigen Receptor T-Cells
CCL2 CC-chemokine ligand 2
CD Cluster of differentiation
CD19 KO Homozygot CD19 defiziente (-/-) Mäuse, sowohl für
murines als auch humanes CD19
CD19 Tg Humanes CD19 als transgen (+/+) exprimierende Mäuse
CSF-1 R Colony stimulating factor 1 receptor
CTLA4 Cytotoxic T-lymphocyte-associated Protein 4
DAPI 4’,6-diamidino-2-phenylindole dihydrochloride
DLBCL Diffus großzelliges B-Zell-Lymphom
DMSO Dimethylsulfoxid
DNA Desoxyribonukleinsäure
EBV Epstein-Barr-Virus
EGFR Epidermal Growth Factor Receptor
EV Extrazelluläre Vesikel
Fc Fragment crystallisable
Fab Fragment antigen binding
FCS Fetal Calf Serum
G0/G1/G2-Phase Gap 0/1/2 - phase
GC Germinal center
GITR Glucocorticoid-induced TNFR-related gene
HER2 Human epidermal growth factor receptor 2
HIV Humanes Immundefizienz Virus
VHLA Human leukocyte antigen
huCD19 humanes CD19
ICOS Inducible T-cell Co-Stimulator
IDO Indolamin-2,3-Dioxygenase
Ig Immunglobulin
IL Interleukin
IFN Interferon
IPH-2010 Monoklonaler Antikörper gegen „human anti-NK-cell
inhibitory receptor“
IRES-GFP Internal Ribosomal Entry Site-Green Fluorescent Protein
kDa Kilodalton
KIR Killer cell immunoglobulin-like receptors
KO Knockout
LTR Long Terminal Repeat
M-Phase Mitosis-phase
mAb Monoclonal Antibody
MAP-Kinase Mitogen-activated protein kinase
MHC Major histocompatibility complex
MSCV Murine Stem Cell Virus
muCD19 murines CD19
NK-Zellen Natürliche Killerzellen
OVA Hühnerovalbumin
PAP Prostataspezifische saure Phosphatase
PI3K Phosphoinositid-3-Kinase
PBS Phophate Buffered Saline
PD-L1 Programmed cell death 1 ligand 1
PD1 Programmed cell death protein 1
RAS Rat sarcoma; Proto-Onkogen
S-Phase Synthesis-phase
SH-Domäne Src-homology-Domäne
Src-Familie Akronym aus cellular und sarcoma
T-Zelle Thymus derived cell
TG Transgen
TGF Transforming growth factor
TIL Tumor infiltrierende Lymphozyten
TNT Tunneling nanotubbe
VITVEC Talimogene laherparepvec
UV Ultraviollettes Licht
VEGF Vascular Endothelial Growth Factor
VPD Violet Proliferation Dye
Wnt-Signalweg „Wingless Int-1“ als Mediator des Signalweges
WT Wildtyp
VIIAbstract
Abstract
Background: CD19 is constitutively expressed on B-Lymphocytes and is a central
target antigen in modern immunotherapies for B-cell-neoplasia (1). These therapies
with CD19 specific antibodies or effector cells can be limited by the loss of CD19
rendering target cells invisible. This mechanism is highly relevant: the rate of antigen-
negative relapses after immunotherapy can be as high as 30% (2). This
comprehension addresses the consequences of the loss of CD19 as an important
regulator for B-cell-receptor-signals on the growth of lymphoma cells.
Methods: To compare the effect of CD19, CD19-negative and CD19-positive Burkitt-
like cell lines were established: we cultivated CD19-deficient, λ-myc-transgenic, murine
cell lines. These cell lines were then reconstituted with human CD19 by retroviral
transduction, which led to stable CD19-positive cell lines. These cells were placed into
co-cultivation with the CD19-negative cells. Furthermore, the survival of the CD19-
positive and CD19-negative cell lines was tested by withdrawal of serum and glucose.
Results: After transduction, we observed an autoselection of CD19-positive lymphoma
cells. In co-culture, this selection of CD19-positive lymphoma cells was observed in an
even faster fashion. We analyzed cell proliferation (VPD, BrdU) as well as the
apoptotic-activity (7AAD/Annexin) in co-culture. The results showed no difference
between the CD19-positive and CD19-negative lymphoma cells neither in proliferation,
nor in apoptotic-activity. Thus, our results disprove enhanced proliferation of CD19-
positive lymphoma cells in co-culture. The possibility of intercellular transmission of
CD19 was considered and co-culture experiments with transwell were conducted. The
transwell assay showed no difference in comparison to control co-culture. The
metabolic analysis showed a significantly higher survival rate of CD19-positive
lymphoma cells under deprivation of serum in vitro.
Discussion: The results of the transwell-assay do not disprove the hypothesis of a
transmission of CD19 between lymphoma cells. This could explain the observation in
co-culture and opens an interesting outlook: The transmission of CD19 via membrane-
connections has not yet been described and would be of high relevance for modern
immunotherapies.
1Zusammenfassung
Zusammenfassung
Hintergrund und Ziele: CD19 wird über alle Reifungsstufen der B-Lymphozyten
konstant exprimiert und ist ein zentrales Antigen für Immuntherapien gegen B-Zell-
Neoplasien (1). Diese Therapien mit Antikörpern oder spezifischen Effektorzellen
werden unter anderem durch Resistenzen limitiert: Die Zellen verlieren das Ziel-
Antigen und entziehen sich damit der spezifischen Therapie. Dies ist von hoher
Relevanz, da die Rate an Antigen-negativen Rezidiven nach Immuntherapie bis zu
30% beträgt (2). Welche Auswirkung dieser Verlust von CD19 als Regulator von B-Zell-
Rezeptor-Signalen auf das Wachstumsverhalten von Lymphomzellen hat, ist
Gegenstand dieser Arbeit.
Methoden: Um die Effekte von humanem CD19 auf Lymphomzellen vergleichend zu
beobachten, wurden CD19-negative und CD19-positive Burkitt-like Lymphomzelllinien
erzeugt. Dafür erfolgte die Kultivierung CD19-defizienter, λ-myc-getriebener, muriner
Zellinien. Diese wurden anschließend retroviral mit humanem CD19 transduziert. Die
daraus hervorgegangenen CD19-positiven Zelllinien wurden mit den primären, CD19-
defizienten Zelllinien in Ko-Kultur gebracht. Weiterhin wurden die unterschiedlichen
Zelllinien auf Überleben unter Serum- und Glukoseentzug untersucht.
Ergebnisse und Beobachtungen: Nach Transduktion selektierten sich die CD19-
positiven Lymphomzellen selbst. In Ko-Kultur ließ sich diese Autoselektion in noch
kürzerer Zeit beobachten. Proliferationsanalysen (VPD, BrdU), sowie
Apoptoseanalysen (7AAD/Annexin) erbrachten keinen Unterschied zwischen den
CD19-positiven und CD19-negativen Zelllinien in Ko-Kultur. Folglich wurde ein
Wachstumsvorteil für CD19-positive Lymphomzellen in Ko-Kultur ausgeschlossen. Es
wurde die Möglichkeit der interzellulären Übertragung von CD19 erwogen und Ko-
Kultur-Versuche mit Transwell durchgeführt. Diese zeigten keinen Unterschied zu
Kontroll Ko-Kulturen. Die Stoffwechselanalysen zeigten ein signifikant besseres
Überleben der CD19-positiven Lymphomzellen unter Serumentzug.
Schlussfolgerungen: Die Transwell-Versuche schlossen die Möglichkeit des
interzellulären Austausches von CD19 in Ko-Kultur nicht aus. Dies könnte eine
Erklärung für die Beobachtungen in Ko-Kultur sein und eröffnet damit einen
interessanten Ausblick: Die Übertragung von CD19 über Membranbrücken wurde bis
heute nicht beschrieben und wäre von großer Bedeutung für aktuelle
immuntherapeutische Therapieansätze.
21. Einleitung
1. Einleitung
Diese Einleitung soll als Einführung und Interpretationsstütze für die im Rahmen dieser
Arbeit erhobenen Ergebnisse dienen. Dabei werden die einzelnen Abschnitte vom
Grundlegenden zum Detaillierten führen: Während das erste Kapitel auf zentrale
Prinzipien des Immunsystems eingeht, widmet sich das zweite Kapitel den B-
Lymphozyten, ihrem Reifungsprozess und ihrer Rezeptorstruktur. Das dritte Kapitel soll
die Rolle von CD19 als B-Zell-Marker und Teil des B-Zell-Rezeptor-(BZR)-Komplexes
als Kernstück dieser Einleitung beleuchten. Das vierte Kapitel umreißt die maligne
Entartung von B-Zellen, mit Schwerpunkt auf das Burkitt-Lymphom, welchem die
Versuchszellreihen dieser Arbeit zugerechnet werden können. Schließlich befasst sich
das fünfte Kapitel mit einer kurzen Skizzierung der aktuell angewandten
Immuntherapien, um die Relevanz CD19-abhängiger Prozesse zu unterstreichen. Die
Fragestellung und Zielsetzung der vorliegenden Promotionsarbeit bilden den
Abschluss.
1.1 Grundlegende Prinzipien des Immunsystems
Das Immunsystem hat ein zentrales Ziel: Die kontrollierte und nachhaltige Elimination
von Pathogenen, die in den Körper gelangen und ihn bedrohen. Dies setzt die
Fähigkeit voraus, zwischen körpereigenen und körperfremden Strukturen zu
unterscheiden. Gleichzeitig ist die Auseinandersetzung mit maligne entarteten Zellen
ein relevanter Teilaspekt immunologischer Prozesse. Diese „Immune Surveillance“
wird jedoch seit Jahren kontrovers diskutiert (3).
Insgesamt können drei Kernprinzipien des Immunsystems zusammengefasst werden:
(i) das spezifische Erkennen von Pathogenen, (ii) deren effektive Elimination, sowie (iii)
die Regulations- und Gedächtnis-Funktion für eine ausgewogene und nachhaltige
Immunantwort (4). Diesen Aufgaben wird über humorale und zelluläre Mechanismen
begegnet. Der Fokus soll im Folgenden auf dem zellulären Anteil liegen.
31. Einleitung
1.1.1 Zelluläre Bestandteile des Immunsystems
Alle Zellen des Immunsystems gehen auf pluripotente hämatopoetische Stammzellen
im Knochenmark zurück (5). Hier erfolgt neben der Erythro- und Thrombozytopoese
die Ausreifung der Leukozyten. Mit Einteilung dieses Reifungsprozesses in
lymphatische und myeloische Zellreihen lässt sich folgende weitere Unterscheidung
machen: Auch wenn eng verzahnt, agiert die lymphatische Zellreihe mit Schwerpunkt
(ausgenommen der natürlichen Killerzellen [NK-Zellen]) als spezifische und die
myeloische Zellreihe als unspezifische Immunabwehr. Der spezifischen Abwehr sind
entsprechend die B- und T-Lymphozyten zuzuordnen, der unspezifischen unter
anderem Phagozyten (Makrophagen, neutrophile Granulozyten) und natürliche
Killerzellen (6).
Während die unspezifischen Mechanismen körperfremde Pathogene (in diesem
Kontext als Antigen zu bezeichnen und im Folgenden synonym verwendet) schnell
erkennen, stellt die spezifische Abwehr eine bei Primärkontakt verzögerte, dafür aber
umso effektivere Immunantwort dar. Diese „spezifische Abwehr“ kann zudem als
„adaptiv“ bezeichnet werden: Die Bildung und Persistenz von Gedächtniszellen nach
Antigen-Kontakt ermöglicht eine Anpassung des Organismus an die ihn umgebenden
Pathogene (7). Die nachfolgende Abbildung soll die B-Lymphozyten in den Kontext der
spezifischen Immunabwehr einbetten. Anschließend legt sich der Fokus in Kapitel 1.2
auf die B-Lymphozyten, um auf das zentrale Thema dieser Arbeit, dem CD19-Molekül,
hinzuführen.
Abbildung 1: Aktivierung von T- und B-Lymphozyten. Während die Initiierung der T-
Lymphozyten-Aktivierung abhängig ist von antigen-präsentierenden Zellen, können B-
Lymphozyten ein Antigen direkt binden und damit den Aktivierungsprozess einleiten. Die
darauffolgenden Prozesse der Reifung beider Zellreihen sind über die T-Helferzelle vom Typ 2,
welche die Differenzierung des B-Lymphozyten zur Plasmazelle unterstützt, verbunden (8).
41. Einleitung
1.2 B-Lymphozyten: Entwicklung und Rezeptorstruktur
Das Wissen um die Existenz der B-Lymphozyten wurde nicht auf direktem Wege
erlangt: Es war die Entdeckung von Immunglobulinen im Serum mittels Elektrophorese
durch Tiselius, welche 1939 den ersten Anhaltspunkt erbrachte (9). Etwa zehn Jahre
später konkretisierte sich die Vorstellung einer antikörperproduzierenden Zelle durch
die Beobachtung der Korrelation von Plasmazell-Entwicklung und Antikörperproduktion
nach Immunisierung (10). Der direkte Nachweis dieses Zusammenhangs erfolgte
wenig später unter dem Immunfluoreszenz-Mikroskop (11). 1965 gelang schließlich die
Differenzierung und damit die Benennung der Lymphozyten in B- (bursa-derived) und
T- (thymus-derived) Lymphozyten (12). Im Folgenden soll der Entwicklungsweg der
B-Lymphozyten zusammenfassend dargestellt werden.
1.2.1 Die Entwicklung von B-Lymphozyten
Über die gesamte Lebensspanne eines Menschen stellt das Knochenmark den
Ursprungsort für B-Lymphozyten dar (13). Ausgehend von Progenitorzellen
durchlaufen sie einen komplexen Entwicklungsweg, welcher von frühen lymphatischen
Pro-B-Lymphozyten bis zur reifen B-Lymphozyten im peripheren lymphatischen
Gewebe führt. In jedem Schritt dieses durch verschiedene reifungsinduzierende
Zytokine und wechselnde Oberflächenmarker getriebenen Prozesses, ist eine maligne
Entartung möglich. Auf diese Weise können B-Zell-Neoplasien mit sehr
unterschiedlichem Differenzierungsgrad und damit klinischer Präsentation entstehen
(14). Abbildung 2 widmet sich der Übersichtlichkeit der Reifungsschritte der
B-Lymphozyten.
51. Einleitung
Abbildung 2: B-Lymphozyten-Entwicklung. Stadien der B-Lymphozyten-Reifung in den
jeweiligen Geweben. Während die frühen Stadien im Knochenmark mithilfe der somatischen
Rekombination die Voraussetzung für das Erkennen von Antigenen durch die B-Lymphozyten
schaffen, führt der Kontakt mit einem Antigen zur Differenzierung der reifen B-Lymphozyten in
verschiedene Formen der schnellen und langanhaltenden Immunität (15).
1.2.2 Mechanismen der Antigenerkennung durch B-Lymphozyten
Wie in dem vorangegangenen Kapitel zur Lymphozyten-Aktivierung bereits angedeutet
und in Abbildung 1 dargestellt, steht am Anfang jeder Immunantwort die Erkennung
des Antigens. Ein Antigen kann durch Bindung an ein Immunglobulin (Ig) erkannt
werden. Immunglobuline können in verschiedenen Formen vorkommen: zum Beispiel
membrangebunden als B-Zell-Rezeptor (BZR), oder löslich als Antikörper nach
Sekretion durch eine Plasmazelle (16). Antikörper bestehen dabei aus zwei Teilen:
Eine zur Bindung des Antigens (Fab) und eine zur Rekrutierung von Zellen bzw.
Molekülen der zytotoxischen Immunantwort (Fc) (17). Hiervon abweichend ist der BZR
zusammengesetzt: In seinem Aufbau identisch zu einem Immunglobulin, jedoch über
die konstante Domäne membrangebunden, dient er – zusammen mit weiteren
Molekülen im Komplex – der intrazellulären Signalweiterleitung (18).
61. Einleitung
Abbildung 3: Formen der Antigenerkennung: Der B-Zell-Rezeptor und die Struktur eines
Antikörpers. Die Antikörperstruktur ist in löslicher sowie in membrangebunder Form als B-Zell-
Rezeptor gezeigt. V = Variable Region; C = Konstante Region; H = schwere Kette; L = Leichte
Kette; Fab = Fragment antigen binding; Fc = Fragment crystallisable (18, 19).
1.2.3 Der B-Zell-Rezeptor-Komplex
Die Antigenbindung durch den B-Zell-Rezeptor (BZR) initiiert zusammen mit Zytokinen
und Zell-Zell-Kontakten die Proliferation und Differenzierung der B-Lymphozyten (16).
Dies unterstreicht die Relevanz des BZR für das Überleben von B-Lymphozyten. Wie
in Kapitel 1.2.2 beschrieben, ähnelt der BZR in seinem Aufbau dem eines
Immunglobulins. Damit ist die Fähigkeit zur Bindung eines Antigens an die variable
Domäne der schweren und leichten Ketten gegeben, jedoch keine zur Initiierung von
intrazellulären Signalen. Damit die Erkennung eines Antigens zu einer Immunantwort
führt, ist der BZR auf das Zusammenspiel mit invariablen, akzessorischen Molekülen
angewiesen, welche diese Signalfunktion übernehmen können. Hierbei handelt es sich
um zwei Proteine Ig⍺ (CD79a) und Igβ (CD79b) (20). Zusammen mit dem BZR bilden
sie den B-Zell-Rezeptor-Komplex: Die zwei Proteine sind verantwortlich für den
Transport des Rezeptors an die Zelloberfläche und ermöglichen durch ihre
Signalfunktion die Bildung eines funktionellen Antigen-Rezeptor-Komplexes. Diese
direkte Kopplung von Antigenbindung und Initiierung intrazellulärer Signalwege kann
darüber hinaus im Zusammenspiel mit spezifischen Zelloberflächenproteinen durch
eine zusätzliche Dimension erweitert werden: Der sogenannte Ko-Rezeptor-Komplex
reguliert die Signalstärke sowie die Reizschwelle des BZR-Komplexes (21). Zu diesem
Ko-Rezeptor-Komplex zählen im Wesentlichen drei Oberflächenproteine: CD19, CD21
und CD81, verbildlicht in Abbildung 4 (22).
71. Einleitung
Antigenbindung
Igβ Ig⍺
CD81
CD21
CD19
Signalinitiierung
B-Zell-Rezeptor B-Zell-Rezeptor- B-Zell-Rezeptor-Komplex
Komplex mit Ko-Rezeptor-Komplex
Abbildung 4: B-Zell-Rezeptor im Komplex mit akzessorischen Oberflächenproteinen.
Links: B-Zell-Rezeptor, mit extrazellulär zwei variablen Antigenbindungsstellen. Mittig: der BZR
im Komplex mit den Proteinen der Signalweiterleitung Ig⍺ (CD79a) und Igβ (CD79b). Rechts:
Ko-Rezeptor-Komplex aus CD21, CD19 und CD81 (23, 24).
Im Rahmen des Ko-Rezeptor-Komplexes fungiert CD21 als Rezeptor für den
Komplementfaktor C3d (22). In dem Fall einer Bindung von C3d an CD21, zum
Beispiel durch die Präsenz von C3d auf einem bakteriellen Pathogen, kann der BZR-
Komplex an den Ko-Rezeptor-Komplex gebunden werden. Dies löst die
Phosphorylierung des zytoplasmatischen Anteils von CD19 durch BZR-assoziierte
Tyrosinkinasen aus, welche anschließend unter anderem Kinasen der Src-Familie
rekrutieren. In der Summe führt dies zur Verstärkung der durch den funktionellen
Antigen-Rezeptor-Komplex ausgelösten Signale (25). Die bereits erwähnte Beteiligung
von CD19 an CD21-assoziierten Signalwegen weist auf die signalverstärkende
Wirkung von CD19 hin: Ein Zusammenwirken von CD19 mit dem BZR erhöht unter
anderem die Freisetzung von Calcium, die Aktivität von mitogen-activated protein
kinase (MAP-Kinase) und die Proliferationsgeschwindigkeit der Zelle. Gleichzeitig
zeigen CD19-defiziente Tiere eine immune Hypersensitivität, was auch auf hemmende
Effekte von CD19 hinweist (26). Damit steht CD19 als vielseitiger Regulator des BZR-
Signals im Mittelpunkt des Ko-Rezeptor-Komplexes und wird im anschließenden
Kapitel eingehender beschrieben. Die Rolle des ebenfalls im Ko-Rezeptor-Komplex
befindlichen CD81 ist nicht ausreichend geklärt. Jedoch besteht Grund zur Annahme,
dass die Funktionalität von CD19 in Teilen abhängig vom CD81-Molekül ist (27).
81. Einleitung
1.3 CD19 als spezifischer B-Zell-Marker und tumorassoziiertes Antigen
Das CD19-Protein ist einer der verlässlichsten Biomarker für B-Lymphozyten: Es wird
vom Stadium des Prä-B-Lymphozyten bis zur Plasmazelle exprimiert (24). Auch der
Großteil maligner B-Lymphozyten behält das CD19 auf der Zelloberfläche: 88% der B-
Zell-Lymphome und nahezu 100% der B-Zell-Leukämien sind CD19 positiv (28, 29).
Diese Zahlen unterstreichen die Relevanz von CD19 in der Signaltransduktion des
BZR und als tumorassoziiertes Antigen: Ein Ziel immuntherapeutischer Ansätze zur
Behandlung hämato-onkologischer Erkrankungen.
1.3.1 Der Einfluss von CD19 auf die Signalvermittlung in B-Lymphozyten
CD19 nimmt eine Schlüsselrolle in der Modulation von BZR-abhängigen sowie BZR-
unabhängigen Signalen ein (30, 31). In Versuchen mit transgenen Mäusen (human
CD19Tg) konnte gezeigt werden, dass CD19-Überexpression zu einer verstärkten
Responsivität von B-Lymphozyten gegenüber transmembranen Signalen sowie zu
erhöhten Serum-Immunglobulin-Spiegeln führt (32). Dagegen präsentieren CD19
knock-out (CD19KO) Mäuse signifikant verminderte Immunglobulin-Spiegel im
Serum (33). Folglich ist die Regulation der CD19-Expression von großer Bedeutung für
den Aufbau einer adäquaten B-Zell-Immunität – auch vor dem Hintergrund, dass
gezeigt werden konnte, wie schon geringe Erhöhungen der CD19-Expression von 15-
29% die Bildung von Autoantikörper in Mäusen induzieren kann (34).
Für die Einordnung der vorliegenden Promotionsarbeit sind dabei insbesondere zwei
Aspekte von Bedeutung: Zum einen die Struktur von CD19 und seiner
Signalvermittlung für die Grundlage pathophysiologischer Interpretationen. Zum
anderen die Rolle von CD19 in myc-getriebenen B-Lymphozyten, da diese als Vorlage
der durchgeführten Experimente dienten.
91. Einleitung
i) CD19: Struktur und Signalwege: Das CD19-Protein ist ein 95 Kilodalton (kDa)
Transmembran-Glykoprotein, welches der Immunglobulin-Superfamilie
zuzuordnen ist (35). Das kodierende Gen findet sich auf dem kurzen Arm von
Chromosom 16 und ist darüber hinaus speziesübergreifend in all seinen Exons
sowie regulatorischen 3’ und 5’ Einheiten hochgradig konserviert (zwischen
Menschen und Mäusen 66% Übereinstimmung) (36). Dies lässt die Annahme
einer vergleichbaren Regulation des CD19-Gens bei Menschen und Mäusen zu
(37). Die Darstellung der Struktur von CD19 ist in Abbildung 5 gezeigt.
Abbildung 5: Molekulare Struktur von CD19. Die zwei C2-Immunglobulin-like
Domänen werden durch eine helikale non-Ig-like Domäne getrennt. Die hoch
konservierte, aus 242 Aminosäuren bestehende zytoplasmatische Domäne enthält eine
Vielzahl an Tyrosinresten. Beispielhaft sind hier die drei Schlüssel-Tyrosinreste und ihre
assoziierten Signalkinasen /-moleküle aufgeführt (24).
Der zytoplasmatische Anteil des CD19-Proteins trägt neun Tyrosinreste nahe
des C-Terminus (36). Die meisten dieser Tyrosinreste verfügen über aktive src-
homology-2 (SH2)-Domänen, welche regulatorische Signalkinasen rekrutieren
können (38). Hierbei sind die mit CD19 interagierenden Proteinkinasen unter
anderem folgenden Gruppen zuzuordnen: Src-Familie (Lyn, Fyn), Ras-Familie,
Abl-Tyrosinkinasen, Bruton’s tyrosine kinase (Btk), Adaptermoleküle (Vav,
Grb2) sowie Phosphoinositid-3-Kinasen (PI3K) (39). Die Rekrutierung der
genannten Proteinkinasen ist essentiell für die Transduktion BZR-abhängiger
Signale, da diese die Phosphorylierung durch zum Beispiel die Lyn-Proteinase
benötigen (40). Dabei konnte unter anderem gezeigt werden, dass die Aktivität
der Lyn-Proteinase direkt mit der Expressionsstärke von CD19 korreliert (41).
101. Einleitung
Doch nicht nur mit intrazellulären Proteinen, auch mit weiteren
Transmembranproteinen, insbesondere im Rahmen des Ko-Rezeptor-
Komplexes, finden Interaktionen statt, die bereits in Kapitel 1.2.3. angeschnitten
wurden. Der Kontakt mit CD21 als Komplementrezeptor stellt eine Verbindung
zwischen angeborener und erworbener Immunität dar. Zudem bestehen
Interaktionen mit CD81: CD81 unterstützt den Transport von CD19 an die
Zelloberfläche, was unter anderem zu einer Reduktion der CD19-Expression
bei CD81 -/- Mäusen um 30-50% führt (42, 43).
ii) CD19 im Kontext Myc-getriebener Lymphomzellen: In Untersuchungen von Poe
et al. konnte gezeigt werden, dass CD19- und myc-Expression synergistisch
das Entstehen und die Progression von B-Zell-Lymphomen im Sinne eines
amplification-loops positiv beeinflussen. Dabei beobachtete man bei mycTg-
CD19-/-Mäusen ein signifikant verlängertes Überleben mit einer Zunahme des
medianen Überlebens von > 80% sowie einem späteren Auftreten der B-Zell-
Lymphome. CD19 fungiert also als Ko-Faktor bei durch myc-Überexpression
entstehenden Lymphomen. Darüber hinaus zeigte die genannte Arbeit eine
gegenseitige Beeinflussung des Phosphorylierungsgrades von CD19 und myc
und beschrieb zusätzlich eine erhöhte Dichte von CD19 bei konstitutiver myc-
Expression (31).
Dieser Zusammenhang konnte durch weitere Studien untermauert werden, die
unter anderem CD19 als wichtigen BZR-unabhängigen Induktor von myc-
getriebener B-Zell-Lymphomgenesis identifizierten (44). Zusammenfassend
kann CD19 als Progressionsverstärker bei der malignen Entartung von B-
Lymphozyten betrachtet werden, jedoch nicht als ursprünglicher Initiator. Denn
MycTg-CD19-/-Mäuse, wenn auch mit langsamerer Dynamik, entwickeln
ausnahmslos Lymphome (31). Eine Vertiefung bezüglich B-Zell-Neoplasien und
der Rolle von Myc im Burkitt-Lymphom erfolgt im anschließenden Kapitel.
111. Einleitung
1.4 Lymphatische Neoplasien: Maligne Entartung von B-Lymphozyten
B-Zell-Neoplasien machen neben Neoplasien der T- und natürlichen Killerzellen den
Großteil der menschlichen Lymphome aus (45). Die Beschreibung der sehr
heterogenen Gruppe von Lymphomen geht dabei weit in die Geschichte zurück.
Während Thomas Hodgkin schon 1832 das später nach ihm benannte Hodgkin
Lymphom beschrieb, grenzte Rudolf Virchow 1864 eine von ihm als Pseudoleukämie
bezeichnete Erkrankung ab. Der Begriff des malignen Lymphoms wurde erst 1871 von
Theodor Bilroth geprägt (46, 47).
Heute erfolgt die Einteilung der lymphatischen Neoplasien nach der WHO-
Klassifikation – basierend auf pathologischen, genetischen und klinischen Faktoren
(48). Dabei spielen genetische Aspekte eine immer bedeutendere Rolle, da sie zu
einer genaueren Differenzierung zwischen den einzelnen Lymphomformen beitragen.
Der Grundstein für diesen onkogenetischen Ansatz wurde in den 1980ern durch
Mitchell et al. und Dalla Favera et al. gelegt, denen erstmals die Klonierung des Myc-
Gens mit der Translokation t(8;14) im Burkitt-Lymphom gelang (49, 50). Da in den
Versuchsreihen dieser Promotionsarbeit Burkitt-like Lymphomzellen verwendet
wurden, wird der Fokus des anschließenden Kapitels auf dieser Lymphomunterform
liegen.
1.4.1 Das Burkitt-Lymphom
Hochmaligne B-Zell-Lymphome, wie das Burkitt Lymphom, zeigen ein rasches
Wachstum und gehören zu den am schnellsten wachsenden Neoplasien des
Menschen. Unter intensiver Chemo-Immuntherapie kann der Großteil der Patienten
jedoch geheilt werden (51). Es besteht eine Assoziation mit Infektionen durch das
Epstein-Barr-Virus (EBV) sowie Malaria-Erregern. Der irische Chirurg Burkitt beschrieb
die Erkrankung 1958 erstmals bei Kindern im äquatorialen Afrika (Malaria-Assoziation)
(52). Der Verdacht auf einen Umweltfaktor führte schließlich durch Epstein und
Kollegen 1964 zur Entdeckung von EBV in Burkitt-Lymphom-Zelllinien (53). Während
bei der durch Burkitt entdeckten endemischen Form (malaria assoziiert) eine beinahe
hundertprozentige EBV-Positivität vorliegt, wurden später weitere Formen definiert, die
eine geringere EBV-Positivität aufwiesen: Die sporadische Form (außerhalb der
Malaria-Gebiete) mit etwa 15% und eine Immundefizienz-assoziierte Form (z.B.
assoziiert mit dem Humanen Immundefizienz Virus [HIV] mit etwa 40%) (53).
121. Einleitung
Neben diesen Assoziationen mit EBV, Malaria und Immundefizienz sind genetische
Faktoren – wie die oben erwähnte Translokation t(8;14) – ein zentraler
Entstehungsmechanismus. Entsprechend fußt die Diagnostik des Burkitt Lymphoms im
Wesentlichen auf dem Translokationsnachweis (54). Die charakteristischste und
pathognomonische Eigenschaft des Burkitt Lymphoms ist die Überexpression des Myc-
Gens auf Chromosom 8, meist durch eine balancierte Translokation des Myc-lokus an
einem der drei Immunglobulin Gene (schwere Kette t(8;14), Leichtkette lamdba t(8;22)
oder kappa t(2;8)) ausgelöst (55).
Das Onkoprotein Myc ist einer der Hauptschalter in der Regulation des Zell-Zyklus und
wird in allen proliferierenden Zellen exprimiert. In dieser Rolle ist es auch essentiell für
die Initiierung und Erhaltung von Keimzentren als Ausdruck der dort stattfindenden B-
Zell-Proliferation (56). Die meisten wichtigen positiven Zell-Zyklus-Regulatoren (u.a.
Cycline, Cyclin-abhängige Kinasen und Transkriptionsfaktoren) werden durch Gene
kodiert, welche von Myc induziert werden. Zusätzlich kann Myc Zell-Zyklus-Inhibitoren
wie p21 und p27 auf verschiedenen Wegen blockieren (57). Folglich konnte im Model
mit transgenen Mäusen gezeigt werden, dass die Überexpression von Myc in B-
Lymphozyten spontan zur Entstehung von hoch aggressiven B-Zell-Lymphomen
führt (58).
1.5 Konzept der Immuntherapie
Hoch aggressive Lymphome können durch intensive Chemotherapie kurativ therapiert
werden. Jedoch hat der Einzug der Immuntherapien die Erfolge in der
Lymphomtherapie deutlich gesteigert, auf Subgruppen erweitert und nachhaltiger
gestaltet. Die Idee, das in seiner Zytotoxizität enorm effiziente Immunsystem gegen
maligne entartete und damit körpereigene Zellen zu richten, ist die Grundlage für das
Konzept der Immuntherapie. Dieses Vorgehen erfordert die Umgehung der eingangs
besprochenen zentralen Toleranz. Anders gesagt muss der Tumor aus dem “blinden
Bereich” des Immunsystems herausgehoben und sichtbar gemacht werden. Auf
unterschiedlichen Wegen (im Folgenden Tabelle 1) wird das Immunsystem deshalb
gegen körpereigene, aber tumorspezifische Antigene gerichtet. In der Konsequenz wird
deutlich, dass nicht jedes Antigen beliebig als Therapieziel dienen kann: Seine
Spezifität für die jeweilige Tumorentität ist ausschlaggebend für den Therapieerfolg und
die potenziellen Nebenwirkungen.
131. Einleitung
1.5.1 Aktueller Entwicklungsstand der Immuntherapie
Die Entwicklung der Immuntherapie reicht zurück bis in das neunzehnte Jahrhundert,
in dem Berichte über die Behandlung von Sarkomen mit der lokalen Injektion
abgetöteter Bakterien durch den Chirurgen William Coley veröffentlicht wurden (59). In
der Gegenwart ermöglicht das immunologische Verständnis sowie das technische
Know-how eine zielgerichtetere Therapie als die der diffusen Aktivierung des
Immunsystems durch bakterielle Pathogene. Im Folgenden soll ein kurzer,
strukturierter Überblick über die aktuell angewendeten und in Entstehung begriffenen
Formen der Immuntherapie gegeben werden.
Die Bedeutung von CD19 für neue Immuntherapieformen wird dabei insbesondere bei
chimeric antigen receptor (CAR)-T-Zellen und bispezifischen Antikörpern ersichtlich.
Dies unterstreicht die Relevanz des besseren Verständnisses von CD19-abhängigen
Prozessen und bildet damit den Abschluss der Einleitung zu der vorliegenden Arbeit.
141. Einleitung
Tabelle 1: Überblick aktueller Immuntherapien
Therapieform Mechanismus Bekannte Vertreter
Zytokine Multiple Immunmodulation Il-2, Lenalidomid, IFN,
BCG
Monoklonale Antikörper Opsonierung mit in Folge: CD20: Rituximab
-Fc Rezeptor-Antikörper-abhängige Zytotoxizität CD38: Daratumumab
und Phagozytose VEGF: Bevacicumab
-Komplementaktivierung EGFR: Cetuximab
HER2: Trastuzumab
Checkpoint-Inhibition Hemmung der Apoptoseinduktion durch PD- PD1: Nivolumab
L1/PD-L2/CTLA4 PD-L1: Atezolizumab
CTLA4: Ipilimumab
Agonismus Stimulation der T-Zell-Aktivität 4-1BB (CD137)
kostimulatorischer OX-40 (CD134)
Rezeptoren GITR (CD357)
ICOS (CD287)
CD40
CD28
Kombinierte Strategien Effektverstärkung durch doppeltes Checkpoint- Ipilimumab + Nivolumab
Inhibition
T-Zell-Manipulation Chimeric Antigen Receptor T-Cells (CART): CART gegen CD19 bei B-
spezifische T-Zell-Rezeptoren durch genetische Zell Neoplasien
Manipulation autologer T-Lymphozytn.
Ex-vivo Stimulation tumorinfiltrierender TIL’s Therapie bei
Lymphozyten malignem Melanom
Bispezifische T-Zell-Engager Antibodies (BITEs): Blinatomumab
induzierte T-Zell-zytotoxische Tumoreradikation (CD19 +CD3)
Onkolytische Viren Erhöhung der Immunogenität des Talimogene laherparepvec
Tumorenvironments (TVEC)
Zellen des NK-Zellen: Anti-KIR-Antikörper IPH2101+Lenalidomid
Tumorenvironments Lirilumab
Macrophagen: CCL2, CSF-1R Emactuzumab
IDO-Inhibition Imatinib
Impfung Erzeugung von Gedächtniszellen spezifisch für sipuleucel-T (PAP)
tumorassoziierte Antigen
(60)
152. Zielsetzung der Arbeit
2. Zielsetzung der Arbeit
Die in der Einleitung beschriebenen Funktionen von CD19 in der B-Lymphozyten-
Differenzierung und seine Rolle als Antigen im Rahmen von Immuntherapien ergeben
die folgenden Fragen als Grundlage für die hier vorgelegte Arbeit:
1.) Welche Konsequenzen hat der Verlust von CD19 für den Stoffwechsel und die
Proliferation von hochmalignen B-Zell-Lymphomen?
2.) Lassen sich Unterschiede zwischen CD19-negativen und CD19-positiven B-
Lymphozyten herausarbeiten, die das Verständnis für Mechanismen des
Antigen-Verlustes verbessern?
Wie bereits in der Einleitung beschrieben, ist das CD19-Antigen ein wesentlicher
Bestandteil des Ko-Rezeptor-Komplexes des BZR. In diesem Zusammenhang
moduliert es wesentliche BZR-abhängige Signale. Damit stellt es für den einzelnen B-
Lymphozyt eine zentrale Vermittlungsstelle von Wachstumssignalen dar. Es ist folglich
davon auszugehen, dass der Verlust von CD19 einen Nachteil für das Wachstum von
B-Lymphozyten darstellt. Die Tatsache, dass CD19 bis zum Stadium der
Ausdifferenzierung in Form der Plasmazelle exprimiert wird, zeigt indirekt, wie
abhängig der B-Lymphozyt von dem Signal des CD19-Moleküls ist. Entsprechend
eignet es sich in besonderem Maße als Ziel-Antigen für Immuntherapien, da Escape-
Varianten unwahrscheinlich erscheinen.
Umso interessanter ist der Zustand der CD19-negativen B-Lymphomzelle. Trotz der
soeben beschriebenen Abhängigkeit treten Antigen-negative Rezidive nach
Immuntherapien in relevanter Zahl auf. Das bessere Verständnis über den Unterschied
zwischen CD19-negativen und CD19-positiven Lymphomzellen ist deshalb hoch
aktuell. Die oben aufgeführten Fragestellungen der vorliegenden Arbeit sollen zu
diesem Verständnis beitragen.
163. Ergebnisse
3. Ergebnisse
Um den Effekt von CD19 auf das Wachstum und Überleben von B-Zell-Lymphomen
untersuchen zu können, mussten zunächst CD19-negative und CD19-positive
Zelllinien etabliert werden. Die Grundlage bildeten die CD19-defizienten Zelllinien
NZ880, NZ924 und NZ927. Diese Linien wurden durch Rückkreuzung von dem λ-Myc
transgenen CD19 Wildtyp (CD19WT) mit homozygoten CD19-defizienten CD19-/-
Tieren (CD19KO) generiert (Mäuse von der Arbeitsgruppe Prof. Dr. Hans-Martin Jäck).
Diese Tiere entwickelten spontane Lymphome mit ähnlicher Latenz wie die CD19-
positiven Wildtyp-Mäuse. Sie sind vergleichbar mit Mäusen aus Studien von Kovalchuk
et al. Diese zeigten, dass transgen λ-Myc-mutierte Mäuse Lymphome entwickeln, die
in Phänotyp und Histologie stark dem humanen Burkitt-Lymphom ähneln (55). Die
primären B-Zell-Lymphome der oben genannten Kreuzung ließen sich kultivieren.
Anschließend erfolgte die retrovirale Transduktion der genannten murinen CD19
(muCD19)-negativen Zelllinien mit humanem CD19 (huCD19) durch ein retrovirales
Plasmid. Hiermit wurde die Grundlage für alle folgenden vergleichenden
Untersuchungen zwischen CD19-negativen und CD19-positiven Lymphomen gelegt.
Das Modell der retroviralen Transduktion CD19-defizienter, muriner B-Lymphomzellen
mit einem Vektor für humanes CD19 ist dabei als funktionell anzusehen: In der
Literatur konnte gezeigt werden, dass humanes CD19 auf murinen CD19-defizienten
B-Lymphozyten seine physiologische Funktion erfüllen kann (61). Wenn in den
folgenden Kapiteln von CD19 gesprochen wird, ist damit das humane CD19 gemeint.
3.1 Autoselektion CD19-positiver B-Lymphomzellen nach retroviraler
Transduktion
Für die Erstellung des verwendeten retroviralen Vektors wurde die Sequenz der
huCD19-Desoxyribonukleinsäure (DNA) in einen Murine Stem Cell Virus (MSCV)-
Vektor kloniert. Der Vektor E3145 wurde hierfür aus zwei Vektoren (E3150 und E2815)
zusammengeführt. Dabei wurde die CD19-Sequenz (isoliert aus E2815) zwischen
einer 5’ und 3’-Long Terminal Repeat (LTR)-Promotorregionen des MSCV-Vektor
E3150 an Stelle der für Hühnerovalbumin (OVA) und den Internal Ribosomal Entry
Site-Green Fluorescent Protein (IRES-GFP) kodierenden Regionen platziert. Das damit
entstandene Ausgangsplasmid verfügte außerdem über Informationen für eine
Ampicillin-Resistenz. Mit Hilfe des entstandenen Plasmids konnte die retrovirale
Transduktion der CD19-negativen Lymphomzelllinien mit huCD19 durchgeführt
173. Ergebnisse
werden. Hierfür wurde zunächst mit der Verpackungszelllinie Phoenix E ein
Virusüberstand hergestellt (die genaue Beschreibung findet sich im Abschnitt
Methoden). Für die retrovirale Transduktion wurden 125.000 Zellen der jeweiligen
CD19-negativen Zelllinie in frisches Zellmedium aufgenommen und anschließend der
Virusüberstand zusammen mit Polybrene hinzugegeben. Es wurde der 48h
Virusüberstand verwendet, da dieser erfahrungsgemäß die höchsten Virustiter
aufweist. Danach schloss sich eine dreieinhalbstündige Zentrifugation bei 33°C und
Überführung in frisches Zellmedium an. Die Transduktionseffizienz wurde nach 24h mit
einer Färbung auf humanes CD19 durchflusszytometrisch geprüft. Um die effizienteste
Konzentration des 48h Virusüberstandes zu ermitteln, wurde dieser Vorgang mit
unterschiedlichen Konzentrationen des Virusüberstandes wiederholt. Wie in Abbildung
6 zu sehen, zeigte der unverdünnte Virusüberstand die höchste Transduktionseffizienz
mit 8,46%.
Abbildung 6: Retrovirale Transduktion mit verschiedenen Verdünnungsstufen des
Virusüberstandes.
(A) Durchflusszytmetrische Analyse der huCD19-Positivität nach retroviraler Transduktion: Die
y-Achse gibt die Anzahl der erfassten Zellen an, die x-Achse die Fluoreszenz für CD19. Nur bei
Verwendung des unverdünnten und des 1:1 verdünnten Überstandes konnten CD19-positive
Populationen sichtbar gemacht werden. Zum Vergleich ist die CD19-negative Kontrolle in den
Abbildungen grau hinterlegt.
(B) Rechts: Der Graph zeigt auf der y-Achse die durchflusszytometrisch prozentuale Menge
CD19-positiver Zellen. Die x-Achse zeigt in abnehmender Konzentration die verwendete
Verdünnung des Virusüberstandes. Die Transduktionseffizienz nimmt exponentiell mit der
Verdünnung des Virusüberstandes ab. Links: Prozentwerte der CD19-Positivität nach
Transduktion der verwendeten Zelllinie NZ880.
183. Ergebnisse
24h nach Transduktion wurde bei den verschiedenen Zelllinien wiederholt eine
Transduktionseffizienz von ca. 10% erreicht. In den Tagen nach Transduktion zeigte
sich eine prozentuale Zunahme an CD19-positiven Zellen. Dieser sich über etwa
zweieinhalb Monate ziehende Verlauf ist beispielhaft in Abbildung 7 dargestellt. Er ließ
sich mit den verschiedenen, transduzierten CD19-negativen Zelllinien wiederholen.
Abbildung 7: Autoselektion CD19-positiver B-Lymphomzellen nach retroviraler
Transduktion. Der an Tag 10 (d10) nur 10%ige Anteil CD19-positiver Zellen erhöhte sich
innerhalb von etwa 80 Tagen auf 80%. Im weiteren Verlauf etablierte sich unter standardisierten
Kulturbedingungen (siehe Material und Methoden) eine CD19-positive Zelllinie ohne weitere
äußere Einflussnahme. Abgebildet ist ein typischer Verlauf, exemplarisch gewählt aus
insgesamt sechs Experimenten mit jeweils mehreren Zelllinien (NZ880 P3, NZ877 P1, NZ927
P5, NZ 1001 P0).
Zusammengefasst konnte gezeigt werden, dass die retrovirale Transduktion CD19-
negativer Zelllinien mit huCD19 eine gemittelte Transduktionseffizienz von 10%
aufweist. Diese anfänglich unterlegene Population CD19-positiver Zellen reichert sich
über 2-3 Monate an, sodass eine vollständig CD19-positive Zelllinie entsteht.
193. Ergebnisse
3.2 CD19 als Selektionsvorteil in Ko-Kultur mit primär CD19-defizienten
B-Lymphomzellen
Zunächst sollte das Hochwachsen CD19-positiver B-Lymphomzellen, welches nach
Transduktion beobachtet wurde, reproduziert werden.
3.2.1 Proliferationsvorteil CD19-positiver B-Lymphomzellen in Ko-Kultur mit
CD19-negativen Geschwisterzellen
Um die Hypothese des Proliferationsvorteiles CD19-positiver Lymphomzellen weiter zu
untermauern, wurden Mischkulturen aus CD19-negativen und -positiven
Lymphomzellen hergestellt. Für diesen Ansatz wurden CD19-negative mit CD19-
positiven B-Lymphomzellen in einem Verhältnis von 10:1 gemischt. Während der
durchflusszytometrischen Analysen an den Tagen zwei, vier und sechs konnte
beobachtet werden, wie die CD19-positiven Zellen innerhalb von sechs Tagen die
CD19-negativen Zellen überwuchsen. Beispielhaft ist ein Verlauf in Abbildung 8 zu
sehen.
Abbildung 8: Ko-Kultur CD19-positiver B-Lymphomzellen mit CD19-negativen
Geschwisterzellen. Durchflusszytometrische Analyse von Ko-Kulturen, CD19-pos. und -neg.
Lymhphomzellen. Die y-Achse zeigt die Zellzahl, die x-Achse die gemessene Fluoreszenz für
CD19. Im Verlauf der Messzeitpunkte an Tag 2, 4 und 6 sieht man eine deutliche Zunahme
CD19 positiver Zellen. Diese Zunahme führt an Tag 6 zu einem Verhältnis von 20% CD19-
negativen und 80% CD19-positiven Zellen.
Bei einem Anteil von 10% nach dem Ansetzen der Ko-Kultur an Tag 0, verlief die
anschließend zu beobachtende Zunahme des Anteils der CD19-positiven Zellen erst
zögerlich (d2 etwa 10%), ab dem 4. Versuchstag dann stärker (d4 etwa 20%; d6 >
80%). Diese Beobachtung ließ sich mit drei unterschiedlichen Zelllinien mehrfach und
konstant wiederholen. Aus Gründen der besseren Stabilität unter Kulturbedingungen
und der Übersichtlichkeit, wurde für weitere Untersuchungen daher exemplarisch die
203. Ergebnisse
Ursprungszelllinie NZ880 P3 und ihre CD19-positiv transduzierte Geschwisterzelllinie
NZ880+E3154 verwendet.
In Anbetracht der Ergebnisse der Ko-Kultur-Untersuchungen ergab sich die Frage, ob
CD19-positive Zellen stärker proliferieren oder weniger häufig in Apoptose gehen. In
Studien mit CD19-transgenen Mäusen (CD19TG+/+) konnte gezeigt werden, dass eine
CD19-Überexpression zu höheren Proliferationsraten und längerem Überleben führt
(24). Entsprechend ergab sich die Hypothese, dass die CD19-vermittelten
Wachstumssignale, die den CD19-negativen Zellen fehlen, den entscheidenden
Unterschied in Ko-Kultur ausmachen könnten.
3.2.2 Untersuchungen der Proliferationsgeschwindigkeit in Ko-Kultur
Im Weiteren sollte die Proliferationsgeschwindigkeit CD19-negativer und CD19-
positiver Zellen untersucht werden. Hierzu wurden zwei Methoden verwendet:
- Eine Violet Proliferation Dye (VPD) 450-Färbung der Zellen, welche Aufschluss über
die Zellteilungshäufigkeit der einzelnen Zellen geben sollte.
- Eine Zellzyklusanalyse, die bei stärkerer Proliferation der CD19-positiven Zellen eine
vermehrte Repräsentation dieser Zellen in der Synthesis (S)-Phase aufzeigen sollte.
3.2.2.1 Zellteilungshäufigkeit
Mit der Anfärbung der Zellen mit VPD sollte die Hypothese überprüft werden, dass in
Ko-Kultur die CD19-positiven Zellen stärker proliferieren als ihre CD19-negativen
Geschwisterzellen. Bei VPD handelt es sich um eine fluoreszierende Färbung, welche
sich an intrazelluläre Proteine bindet. Mit jeder Zellteilung halbiert sich somit die
Konzentration des Farbstoffes, sodass über die Zeit die Abnahme der Fluoreszenz der
Zellen gemessen werden kann. Die Abnahme der Fluoreszenz ist linear zur
Teilungshäufigkeit. Es wurden sowohl die Ko-Kultur als auch die Ursprungszelllinien in
Einzelkultur mit VPD angefärbt und anschließend in gleichen Zeitabständen
durchflusszytometrisch gemessen - in gleicher Weise wie eine Kontroll-Ko-Kultur.
213. Ergebnisse
Wie in Abbildung 9 zu sehen, nahm innerhalb der Ko-Kultur die VPD-Fluoreszenz von
der CD19-positiven Zellpopulation synchron mit der CD19-negativen Population ab.
Demnach konnte keine höhere Proliferationsgeschwindigkeit der CD19-positiven
Zelllinie in Ko-Kultur mit dieser Methode nachgewiesen werden.
CD19 d6 d4 d2 d0
negativ
B220
CD19 negativ
CD19 positiv
Anzahl der Zellen
huCD19
CD19
positiv
B220
huCD19 VPD
Abbildung 9: Bestimmung der Proliferationsgeschwindigkeit mit VPD in Ko-Kultur.
Durchflusszytometrische Untersuchung der Zellkulturen CD19-positiver und CD19-negativer
Zelllinien in Ko-Kultur. Links: die Durchflusszytometrie der verwendeten Zelllinien. Rechts: die
Abnahme der VPD-Fluoreszenz mit der Zeit im Histogramm (Zellzahl y-Achse, Fluoreszenz von
VPD x-Achse). Es zeigt sich eine Abnahme der relativen Fluoreszenz von 105 auf 0 innerhalb
von sechs Tagen. Diese Abnahme ist für beide Zelllinien vergleichbar.
223. Ergebnisse
3.2.2.2 Zellzyklusanalysen
Als zweite Methode zur Prüfung der Hypothese eines Proliverationsvorteils CD19-
positiver Zellen in Ko-Kultur wurden Zellzyklusanalysen mit Bromdesoxyuridin (BrdU)
durchgeführt. BrdU ist ein Thymidin-Analogon und wird daher während der Phase der
DNA-Replikation (S-Phase) in die DNA der Zellen eingebaut. Folglich zeigen stark
proliferierende Zellen eine höhere Anreicherung von BrdU. Der Nachweis von BrDU in
der Zelle erfolgt über einen spezifischen Antikörper. Die Methode basiert dabei auf der
Kombination aus einem DNA-Farbstoff 4’,6-diamidino-2-phenylindole dihydrochloride
(DAPI) zur stöchiometrischen Erfassung des DNA-Gehaltes einer Zelle (Doppelt mit
Abschluss der S-Phase) und dem BrdU-Farbstoff als Marker der DNA-
Syntheseaktivität (62). Als Messzeitpunkt wurde der Tag 6 in Ko-Kultur gewählt, da in
dieser Phase meist der Umschlag hin zu einer mehrheitlich CD19-positiven Zellkultur
stattfand. Die Zellen wurden an Tag 6 in Ko-Kultur zunächst mit BrdU inkubiert und
danach mit DNAse behandelt, um durch diesen Denaturierungsschritt eine Bindung
durch den anschließend hinzugegebenen fluoresziereneden BrdU-Antikörper zu
ermöglichen. Abschließend konnte die Färbung mit DAPI und mit
Oberflächenantikörpern (in diesem Fall für CD19) erfolgen. Abbildung 10 zeigt
exemplarisch die durchflusszytometrische Analyse (Abb. 10, A). Die aus den fünf
Versuchsreihen hervorgegangenen Werte wurden gemittelt (Abb.10, B). Dabei liegt
das Augenmerk bei der ursprünglichen Fragestellung nach dem Proliferationsvorteil
CD19-positiver Zellen vor allem auf der S-Phase: Der Anteil an BrdU-positiven Zellen
von der gesamten Zellpopulation repräsentiert, wie oben beschrieben, diese Phase der
DNA-Reproduktion als Ausdruck der Proliferation. Es zeigte sich, dass sich CD19-
positive Zellen nicht signifikant (p=0,5476) vermehrt in der S-Phase befinden, als
CD19-negative Zellen. Infolge dessen kann das deutliche Überwachsen innerhalb der
Ko-Kultur nicht durch eine vermehrte Proliferation der CD19-positiven Zellen erklärt
werden.
23Sie können auch lesen

















































