Von spontanen Mutanten zu CRISPR/Cas9 - Genveränderung als Treiber der immunologischen Forschung - UZH
←
→
Transkription von Seiteninhalten
Wenn Ihr Browser die Seite nicht korrekt rendert, bitte, lesen Sie den Inhalt der Seite unten
Aus der Forschung
Genveränderung als Treiber der immunologischen Forschung
Von spontanen Mutanten
zu CRISPR/Cas9
Johannes vom Berg, Thorsten Buch
Die Analyse von modifizierten Genen ist zu einem wesentlichen Bestandteil im Methodenrepertoire der Immunolo-
gie geworden. Musste anfangs auf natürliche Mutanten zurückgegriffen werden, so wurden später gentechnische
Methoden entwickelt, die gezieltere Ergebnisse lieferten. Künstliche Genveränderung begann in den 1980ern mit
der Überexpression von Genen durch Transgenese mittels Vorkerninjektion, gefolgt kurze Zeit später von der geziel-
ten Genveränderung in embryonalen Stammzellen. Der nächste Entwicklungsschub waren dann die Zink-Finger-
und Tal-Effektor-Nukleasen, die erstmals gezielte Genveränderung nun im einzelligen Embryo ermöglichten. Der
anschließende Siegeszug der CRISPR/Cas9-Technologie wurde im Prinzp maßgeblich durch diese frühen Designer-
Nukleasen erleichtert. Heute werden nicht nur in der Maus, sondern in vielen anderen Spezies gezielte Genom-
modifikationen mittels CRISPR/Cas9 durchgeführt. Auch für genetische Screens, seien es nun Aktivierungs- oder
Inaktivierungsscreens, hat sich CRISPR/Cas9 als geeignetes Tool etabliert. Cas9-Orthologe von unterschiedlichen
Bakterienspezies tragen zudem zu einer sehr guten Abdeckung über vollständige Genome bei.
Schlüsselwörter: CRISPR/Cas9, Transgenese, gezielte Genveränderung
Immunologische Forschung mittels damit wichtige Elemente in hypothesen- schrieb Ogden Bruton schon 1952 die
Genmutationen getriebener biomedizinischer Forschung Agammaglobulinämie in einem 8-jähri-
In der Immunologie hat das Verständ- sind. Speziellere Genomveränderungen gen Kind [2]. Mit der Einführung von
nis von genbasierten Mechanismen unser sind dann noch solche, in denen Punkt- Modellorganismen wie Maus und Ratte in
Wissen maßgeblich geformt. Um die mutationen eingebracht, oder mit denen die immunologische Forschung und der
Rolle von einzelnen Genen für das Gene aus anderen Tierarten oder dem damit einhergehenden Etablierung von
Immunsystem zu untersuchen, haben sich Menschen ausgeprägt werden. Die Unter- Inzuchtstämmen, in denen alle Individu-
genveränderte Lebewesen als wichtiges suchung von unterschiedlichen Varianten en identische Gene tragen, wurden bald
Hilfsmittel herausgestellt. Im einfachsten eines Gens (Allelen) geht letztendlich auf auch die ersten vererblichen Veränderun-
Fall wird das zu untersuchte Gen inakti- Gregor Mendel zurück – wenngleich er gen am Immunsystem dieser Nager be-
viert. Komplementär ist die Überexpres- sich für Merkmale der Erbse und nicht für schrieben.
sion, die künstliche Ausprägung eines das Immunsystem interessierte [1]. Später
(Trans-)Gens generell oder in speziellen wurden dann die ersten immunologi- Spontane Mutanten
Zelltypen. Beide Ansätze werden gerne schen Syndrome im Menschen beschrie- Das Erbgut ist weit weniger stabil als
zusammen verwendet, da sie ein vollstän- ben. Die genetischen Ursachen blieben man zunächst annehmen möchte. Für die
digeres Bild der Genfunktion liefern und zunächst jedoch im Verborgenen. So be- Labor-Maus ist eine Mutationsrate von
96 Trillium Immunologie 2018; 2(2)Aus der Forschung Abb. 1: Eine kleine Geschichte der Genommanipulation in der Maus Natürliche Mutanten werden durch Zufall in größeren Kolonien entdeckt. Bei der konventionellen Transgenese wird das Transgen durch Injektion in den Vorkern eines einzelligen Embryos in das Zielgenom eingebracht. In der ES-Zelltechnologie werden ES-Zellen in vitro mittels "Gene Targeting" modifiziert und die modifizierten Zellen dann wieder in einen Embryonen (Blastozysten) eingebracht. Die daraufhin geborenen Schimären können vollständig manipulierte Nachkommen hervorbringen. Bei der CRISPR/Cas9-Technologie wird wieder meist im Ein-Zellstadium gearbeitet, wo das Cas9 einen durch die gRNA vorgegebenen Schnitt im Genom durchführt. Dieser kann entweder ungezielt oder durch Vorlage von Matrizen repariert werden. 5,4 × 109 für kleine Veränderungen wie dramatische Phänotypen mit in einigen wie die Inaktivierung eines Gens. Für die z. B. Basenaustausche beschrieben worden Fällen dem gleichen modifizierten Gen, Überexpression wurde lange Zeit vor al- [3], andere Mutationen, wie z. B. Verände- Prkdc, sind ebenso in der Humanmedizin lem konventionelle Transgenese verwen- rungen in Small Tandem Repeats, sind bekannt [6]. Auch wenn dort das Wohl der det. Bei dieser wird ein Expressions- sogar noch weitaus häufiger. So ist es kein Patienten immer im Vordergrund steht, konstrukt, im einfachsten Fall bestehend Wunder, dass aufmerksame Wissen- so haben solche Genveränderungen mit aus Promoter, cDNA und Polyadenylie- schaftler schon früh spontane Mutationen immunologischen Konsequenzen essen- rungssignal, in einen Vorkern einer be- von Genen mit elementaren Funktionen ziell zu unserem Verständnis des mensch- fruchteten Eizelle injiziert [12, 13]. Das für das Immunsytem beschrieben haben. lichen Immunsystems beigetragen und Konstrukt inseriert u. a. durch einfache Hier sind als Beispiele die nude-, die scid- helfen die in Tiermodellen gewonnenen Reparatur (non-homologous end joining, und die lpr-Mutationen zu nennen. Mit Detailkenntnisse im Menschen zu validie- NHEJ) oder Mikrohomologien (micro- der nude-Mutation im foxn1 gene konn- ren. Unsere dritte Beispielmutation, lpr, homology-mediated end joining, MMEJ) ten grundlegende Funktionen des Thymus führt zu einem komplexeren Krankheits- an einer zufälligen Stelle im Genom, für T-Zellen herausgefunden werden [4]. bild, einer Autoimmunerkrankung, die meist nicht einzeln sondern mehrfach [14, Durch das Fehlen der thymusabhängigen dem Lupus erythematodes sehr ähnlich 15]. Die Expressionshöhe wie auch die T-Zellen diente und dient dieser Maus- ist [7]. Ursächlich ist eine Punktmutation Expressionskontrolle, also Ausprägung stamm vielfältigen immunologischen im Fas-Gen [8], eines für Zelltod, Teilung im avisierten Zelltyp, war aus diesem Forschungsprojekten einschließlich sol- und Aktivierung wichtigen Gens. Auch Grund wie auch aufgrund der Einflüsse chen mit Bezug zu Organtransplantation für die nude- wie für die lpr-Mutationen der umgebenden, nicht vorhersehbaren oder Krebstherapie. Die scid-Mutation sind die menschlichen Pendants beschrie- Genregionen, recht zufällig; Transgen- greift noch grundlegender in das Immun- ben [9–11], gute Beispiele für die Transla- inaktivierung kann auch noch in späteren system ein und führt zur Abwesenheit von tierbarkeit von im Tier gewonnenen Er- Generationen beobachtet werden. Viele beiden großen Lymphozytenpopulatio- kenntnissen. neu hergestellte transgene Tiere müssen nen, den B- und den T-Zellen, aufgrund bisweilen untersucht werden, bis solche fehlender Erstellung der Antigenrezepto- Transgenese für die Überexpression mit erwünschter Expression gefunden ren beider Zelltypen. Die scid-Mutation Wie schon oben beschrieben, werden werden, ein klarer Nachteil dieser Metho- der Maus wurde von Bosma und Kollegen häufig zwei komplementäre Forschungs- de. Da sie aber die einzige Möglichkeit im Jahr 1983 [5] beschrieben – ähnlich ansätze verfolgt: die Überexpression so- war, das Genom eines Säugers, meist der Trillium Immunologie 2018; 2(2) 97
Aus der Forschung
Maus, zu verändern, fand diese Methode für die Zellkultur zu verändern. Hierzu nur erdenklichen Mutanten verwendet
schnell Anklang bei Immunologen. Sie wurde ein Targeting-Vektor als Plasmid werden. Gene wurden mit Markergenen
war zunächst angewendet worden, um die konstruiert, welcher unter anderem Se- wie dem GFP modifiziert, um ihre Ex-
biologischen Eigenschaften von Antikör- quenzen des Zielgens als Homologiearme pression verfolgen zu können, Punktmu-
pern, bzw. B-Zellrezeptoren aufzudecken. enthielt und so nach Elektroporation in tationen wurden eingeführt, um genaue
Beginnend mit einem Igk-Transgen, ge- die Zellen den Einbau durch den exakten Studien zu Proteinfunktionen aber auch
macht von Ralph Brinster, der auch am Mechanismus der homologen Rekombi- zu regulatorischen Elementen zu ermög-
ersten Expressionstransgen überhaupt nation (HR) ermöglichte. Resistenzgene lichen. Als wahrscheinlich extremstes
beteiligt war [16], wurde in der Folge eine als Selektionsmarker unterstützten die (und aufwendigstes Beispiel) darf sicher
Serie von Immunglobulin-transgenen Reduktion der zu screenenden ES-Zell- die vollständige Humanisierung der Im-
Tieren erstellt, mit denen unter anderem klone. Erst wenn die Veränderung in den munglobulingene gelten, welche es nun
die allele Exklusion in der B-Zellentwick- Zellen eindeutig nachgewiesen war, wur- ermöglicht, menschliche Antikörper
lung entdeckt wurde. Wie schon ange- den diese für die Erstellung von soge- direkt in der Maus zu erstellen [20]. Eine
deutet, hat die konventionelle Transgene- nannten Chimären verwendet, wozu ES- Weiterentwicklung der Methodik stellt
se viele Vorteile (einfach durchzuführen, Zellen wieder durch Mikroinjektion in die konditionale Mutagenese dar, die es
schnelle Erzeugung). Solch eine transge- frühe Embryonalstadien eingebracht wur- erlaubt, Genveränderungen auf Organe
ne Linie ist aber doch auch immer ein den und am Aufbau des sich entwickeln- oder Zelltypen bzw. zeitlich zu beschrän-
Zufallsprodukt und lässt daher für den den Organismus teilnahmen. Wenn nun ken. Hierzu wird das zu untersuchende
Forschenden häufig noch viele Fragen Keimzellen in diesen Schimären gebildet Gen mit Zielsequenzen einer Rekombina-
offen. Eine deutlich zielgerichtetere Me- wurden, die von den manipulierten ES- se ausgestattet und mit der transgenen
thode wurde denn auch bald entwickelt. Zellen abstammten, dann konnten diese (z. B. Zelltyp-spezifischen) Ausprägung
Schimären Nachkommen hervorbringen, einer Rekombinase kombiniert. Die Re-
Gezielte Genveränderung in welche die Genveränderung im gesamten kombinase wird in den Zellen, in denen
embryonalen Stammzellen Körper einschließlich der Keimbahn tru- sie aktiv ist, eine Deletion oder Inversion
Während die Transgenese es zum gen. Eine neue genveränderte Linie war durchführen. Das erste und häufigste
ersten Mal ermöglichte, das Genom eines damit erzeugt [18]. Wie unschwer auf- eingesetzte Rekombinationssystem für
Säugers mit einem vorher definierten Ziel grund der Abfolge der verschiedenen konditionale Mutagenese war die Cre-
zu manipulieren und Gensequenzen von Schritte zu erkennen, ist diese Technik Rekombinase in Kombination mit den
Interesse zu Untersuchungszwecken ein- zwar deutlich präziser aber auch aufwen- loxP-Zielsequenzen [21].
zubringen, wurde sie nach kurzer Zeit diger, riskanter und langsamer als die Die ES-Zelltechnik ermöglichte also
durch einen weiteren methodischen An- konventionelle Transgenese über Vor- eine Vielzahl neuer Forschungsansätze.
satz ergänzt, für den die beteiligten For- kerninjektion. Aufgrund ihrer geradezu Einzig die aufwendige Natur der Metho-
scher, Mario R. Capecchi, Martin J. Evans, unbegrenzten Einsatzmöglichkeiten wird de wie auch die Ungewissheit, ob die
und Oliver Smithies den Nobelpreis in sie aber auch noch heute für komplexe Chimären ihre Mutation weitergeben
Physiologie und Medizin 2007 erhielten. Projekte eingesetzt. Die immunologische würden, ließ die Forscher auf bessere,
Bei diesem neuen Ansatz war es möglich, Forschergemeinde hat auch diese Form einfachere Methoden hoffen.
Gene gezielt in exakt vorbestimmter Art der Genveränderung schnell in ihr Tech-
und Weise zu verändern, mithin zu inak- nikportfolio aufgenommen. Als Beispiel Designer-Nukleasen
tivieren. Diese Methode beruhte darauf, mag hier die Inaktivierung des Zytokins Während die ES-zellbasierten Techno-
Zellen eines frühen embryonalen Stadi- Interleukin 4 durch Ralf Kühn im Jahr logien über fast zwei Jahrzehnte die For-
ums im pluripotenten Zustand, als soge- 1991 dienen, welche die Bedeutung von schergemeinde dominierten, wurde im
nannte embryonale Stammzellen (ES- IL-4 für die Antikörperproduktion in Hintergrund an neuen Techniken gearbei-
Zellen) [17], in Kultur zu halten und dort vivo bestätigte [19]. In der Folge konnte tet, die es ermöglichen sollten, Genome
dann mit gängigen Gentechnikmethoden diese Form der Genveränderung für alle effizienter gerichtet zu verändern. Hierzu
98 Trillium Immunologie 2018; 2(2)Aus der Forschung Abb. 2: Genetische Screens gestern und heute Anfangs setzte man auf eine gezielte Mutagenese in der Keimbahn von Tieren. Die resultierenden Mutationen mussten dann umständlich auf Homozygotie gekreuzt werden, bis man auch die rezessiven Merkmale studieren konnte. Die Entwicklung der RNA-Interferenz erlaubte es dann, auch in Zelllinien oder primären Zellen induzierte, nahezu vollständige Funktionsverluste zu untersuchen. Dies konnte sowohl in Zellkultur als auch in vivo durchgeführt werden. Im Gegensatz zu ZFN- und TALEN-Nukleasen eignet sich auch CRISPR/Cas9 hervorragend für genetische Screens; dabei können Forscher inzwischen aus einer großen Anzahl unterschiedlicher Versionen auswählen: Reine Knock-out-Screens mit genetischer Deletion durch unmodifiziertes Cas9-Protein, transkriptionelle Induktion oder Repression von Genakti- vität durch Cas9-Varianten. wurden Designer-Nukleasen entwickelt, HDR. Solche ZFN wurden denn auch er- enten auszuschalten und so die Ausbrei- die ähnlich wie Restriktionsenzyme an folgreich verwendet, um sowohl Gene zu tung des Virus einzudämmen – eine direk- einer vorbestimmten Stelle in einer DNA- inaktiveren wie auch gezieltere Modifika- te klinische Anwendung der Technik [25]. Sequenz schneiden würden. Im Gegensatz tionen durchzuführen – ähnlich derer in Gerade in diesem Kontext darf aber die zu Restriktionsenzymen sollte die Schnitt- embryonalen Stammzellen, aber nun di- Möglichkeit von Designer-Nukleasen, an stelle aber freier wählbar sein und so selten rekt in befruchteten Eizellen wie bei der anderen Stellen im Genom zu schneiden als vorkommen, dass auch ein einzelner konventionellen Transgenese [23]. Einzig geplant, nicht unerwähnt bleiben. Obwohl Schnitt selbst in Säugergenomen möglich die aufwendige Herstellung und der damit diese sogenannten Off-Target-Effekte von sein sollte. Die ersten solcher Enzyme wa- verbundene hohe Preis für ZFN schränkte ZFN zu TALEN weniger wurden, waren sie ren die Zink-Finger-Nukleasen (ZFN). ihren Einsatz stark ein. Sie sollten denn doch gerade im Hinblick auf gentherapeu- Durch Zink-Finger-Domänen wurden spe- auch schnell durch die nächste Generation tische Ansätze eine ernstzunehmende Un- zifische DNA-Zielsequenzen erkannt und an Designer-Nukleasen verdrängt wer- zulänglichkeit. durch die enzymatische Domäne eines den – durch die Tal-Effektor-Nukleasen Restriktionsenzyms nach Dimerisierung (TALEN) [24]. Das einfachere Design, CRISPR/Cas9 als Werkzeug zur der Doppelstrangschnitt durchgeführt [22]. nutzerfreundliche Verbreitung über Plas- Veränderung von Genomen Die Zelle repariert diesen Doppelstrang- midbanken wie Addgene und effiziente Das CRISPR/Cas-System wurde ur- bruch entweder durch NHEJ oder, bei Klonierungsmethoden wie Golden-Gate- sprünglich als Effektormechanismus an- Anwesenheit von homologer DNA z. B. in Klonierung sorgten für eine schnelle Ver- tiviraler Immunität in Bakterien beschrie- Targeting-Vektoren, durch HR. Im Falle breitung in der Forschungsgemeinde, nicht ben [26], und im August 2012 wurden von NHEJ führt dies häufig zu kleinen zuletzt auch, weil TALEN einzelne Nukleo- dann die einzelnen Komponenten und Insertionen oder Deletionen und damit tide binden konnten und nicht Tripletts wie deren Funktionsweise im Detail aufge- gegebenenfalls zu einer (gewünschten) die ZNF. Die Designer-Nukleasen eröffne- klärt, vor allem durch eine bahnbrechen- Inaktivierung des Zielgens. Bei Anwesen- ten vollkommen neue Anwendungsmög- de Studie von Martin Jinek und Kollegen heit von exogen eingebrachter homologer lichkeiten für gezielte Genomveränderun- [27]. Der große Unterschied zu ZFN und DNA als Reparaturmatritze, kann diese gen, Anwendungen, die mit den vorherigen TALEN, die ja auch programmierbare dann im Falle des HDR in das Genom Methoden nicht oder nur schlecht durch- Nukleasen sind, bestand darin, dass Cas9 eingebaut werden – ähnlich der in ES- zuführen waren. So war es mittels ZFN nicht über Protein-DNA-Interaktionen Zellen verwendeten Methode basierend auf möglich, CCR5 in T-Zellen von HIV-Pati- seine Zielsequenz im Genom identifiziert, Trillium Immunologie 2018; 2(2) 99
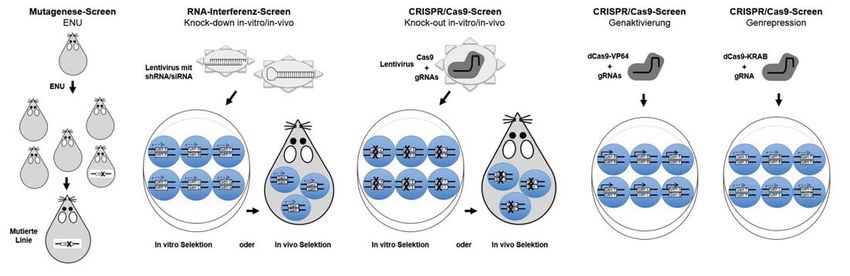
Aus der Forschung sondern über einen RNA-basierten Me- dieses Werkzeugs an ein neues Gen ein- häufig verwendeter Screening-Ansatz be- chanismus mittels Watson-Crick-Basen- fach nur die 20 Basenpaar-lange guide- ruhte auf RNA-interferenz (RNAi), einem paarung. Nur eine kurze Zielsequenz, die Sequenz adaptiert werden muss, war dies natürlichen Mechanismus, mit dem spezi- direkt an die RNA-kodierte Zielsequenz ein enormer Fortschritt, und die einfache fische Proteinexpression auf Ebene der anschließt, ist statisch durch das Cas- und erschwingliche Technologie breitete mRNA unterdrückt bzw. reguliert wird Protein vorgegeben, die sogenannte PAM- sich wie ein Buschfeuer um den Globus [33, 34]. Strenggenommen handelt es sich Sequenz. Für das Auffinden und Binden aus. Es wurde bereits heute in weitaus also nicht um Genveränderungen oder der DNA-Zielsequenz benötigt Cas9 zwei mehr immunologischen Studien verwen- Mutationen, der Effekt kommt einer gene- RNA-Moleküle, von denen eines für die det als ZFNs und TALENs zusammen. Im tischen Ablation aber sehr nahe. Für einen Erkennung der Zielsequenz über einen Folgenden wurden dann auch Versionen Screen wurden siRNA oder shRNA Libra- 20 Basenpaare langen Abschnitt zustän- von Cas9 entwickelt, die, statt zu schnei- ries, die das gesamte Transkriptom zum dig ist (crisprRNA, crRNA), die andere für den, als programmierbares DNA-binden- Ziel hatten, in die zu untersuchenden Zel- die Integration der crRNA in das Cas9- des Molekül agieren und epigenetische len eingebracht. Da die Ploidie keine Rolle Protein (trans-activating crRNA, Veränderungen oder Punktmutationen spielte, konnte direkt auf Funktionsverlust- tracrRNA). Eine Weiterentwicklung des einführen können oder als transkriptio- Phänotypen selektiert werden. Durch die Systems war die Kombination von crRNA neller Aktivator oder Repressor Gene Sequenzierung der nach der Selektion und tracrRNA zu einer chimären single an- oder abschalten können [31]. verbleibenden siRNAs konnte dann direkt guide RNA (sgRNA) [27]. Nun überschlu- auf das komplementäre Zielgen geschlos- gen sich die Ereignisse: Nicht einmal ein CRISPR/Cas9 in genetischen Screens sen werden. RNAi-Screens werden in vivo halbes Jahr später wurde das CRISPR/ Während mittels Funktionsverlust vor allem im Bereich der Tumorimmuno- Cas9-System Anfang 2013 als Designer- oder Überexpression eines Genes Hypo- logie eingesetzt [35], wie auch zur Untersu- Nuklease für den Gebrauch in Eukaryo- thesen zur Genfunktion getestet werden chung von viralen Infektionen [36]. ten von mehreren Gruppen gleichzeitig können, dienen Screens dazu, neue Gene Obwohl mit der RNAi-Technik gene- hoff ähig gemacht. Entscheidend war, das oder Wirkmechanismen zu identifizieren. tische Screens einen Schub bekommen Cas9-Protein mit einem nukleären Loka- Dabei werden erst zufällige Mutationen in hatten, stellte diese Methode den Forscher lisationssignal (NLS) auszustatten und die das Genom eingebracht und anschließend immer noch vor eine Reihe von Proble- kodierende Nukleotidsequenz für die über verschiedene Selektionsmethoden men, wie zeitlich begrenzte oder unvoll- Translation in Säugerzellen anzupassen detektiert. Vor CRISPR/Cas9 wurden zu- ständige Knock-downs und das Unterdrü- [28–30]. Wie ZFNs und TALENs führt fällige Mutationen durch Retroviren, cken unspezifischer Transkripte. Die das aktive Cas9-Enzym Doppelstrangbrü- Transposons, UV-Licht oder über chemi- Methode war aufwendig und nur bedingt che an exakt vorherbestimmten Stellen im sche Reagenzien wie N-ethyl-N-nitrosou- für In-vivo-Screens adaptierbar. Designer Genom ein, allerdings mit weitaus höhe- rea (ENU) in das Genom eingebracht. Nukleasen wie TALEN und ZFN konnten rer Effizienz bei minimalem Aufwand Diese Ansätze wurden anfangs vor allem zwar permanente Veränderungen im Ge- und Kosten. Die Doppelstrangbrüche in der Entwicklungsbiologie verfolgt; pro- nom erzeugen, aber aufgrund der aufwen- werden auch bei CRISPR/Cas9 durch minentes Beispiel hierfür sind die Erkennt- digen und teuren Herstellungsweise und NHEJ oder HR repariert, letzteres, wenn nisse über die embryonale Musterbildung ihrer Funktionweise als Heterodimere eine DNA-Matrize mit in die Zelle einge- in der Fruchtfliege, für die Christiane waren sie ungeeignet für genetische schleust wird. Mittels CRISPR/Cas9 las- Nüsslein-Vollhardt und ihre Kollegen 1995 Screens. CRISPR/Cas9 funktioniert hin- sen sich direkt in einzelligen Embryonen mit dem Nobelpreis für Medizin geehrt gegen allein mit einer einzelnen 20-meren aber auch in anderen Zellen an beliebiger wurden. Eine wichtige immunologische Erkennungssequenz, weswegen schnell Stelle im Genom Knock-outs und Knock- Entdeckung aus diesen Screens war die klar war, dass CRISPR/Cas9 sich exzellent ins realisieren. Im Falle von Maustransge- ganze Familie der Toll-like-Rezeptoren für genetische Screens eignete. Hierzu nese entfällt dadurch die aufwendige ES- (TLR) mit wichtiger Rolle in der Erken- werden ähnlich wie bei RNAi-screens Zellkultur. Nachdem für die Anpassung nung von Pathogenen [32]. Ein weiterer mehrere guide RNAs für jedes Gen in ei- 100 Trillium Immunologie 2018; 2(2)
Aus der Forschung
nem Genom erstellt und zusammen mit Danksagung targeting [see comments]. Science 1994. 265: 103-106.
Wir danken Matthias Heidenreich und Olaf Groß für 22. Kim, Y. G., Cha, J. and Chandrasegaran, S. Hybrid
Cas9 in die zu screenende Zellpopulation ihre hilfreichen Kommentare zum Manuskript. restriction enzymes: zinc finger fusions to Fok I
cleavage domain. Proc Natl Acad Sci U S A 1996. 93:
eingebracht [37, 38]. Auch für Screens hat 1156-1160.
Referenzen 23. Meyer, M. et al. Gene targeting by homologous re-
CRISPR/Cas9 seine Anwendbarkeit in der 1. Mendel, G. Versuche über Pflanzen-Hybriden. Ver- combination in mouse zygotes mediated by zinc-finger
handlungen des Naturforschenden Vereines in Brünn nucleases. Proceedings of the National Academy of
Immunologie bereits bewiesen: Mit einem 1866. 4: 3-47. Sciences of the United States of America 2010. 107:
15022-15026.
CRISPR/Cas9-Screen wurde z. B. schon 2. Bruton, O. C. Agammaglobulinemia. Pediatrics
1952. 9: 722-728. 24. Miller, J. C. et al. A TALE nuclease architecture for
die Lipopolysaccharid-induzierte efficient genome editing. Nature biotechnology 2011.
3. Uchimura, A. et al. Germline mutation rates and
29: 143-148.
Zytokinantwort [38] untersucht und es- the long-term phenotypic effects of mutation accumu-
lation in wild-type laboratory mice and mutator mice. 25. Tebas, P. et al. Gene editing of CCR5 in autologous
senzielle Gene für Anti-Tumor-Immun- Genome Res 2015. 25: 1125-1134. CD4 T cells of persons infected with HIV. N Engl J Med
2014. 370: 901-910.
4. Pantelouris, E. M., Absence of thymus in a mouse
antworten unter Checkpoint-Blockade mutant. Nature 1968. 217: 370-371. 26. Barrangou, R. et al. CRISPR provides acquired
resistance against viruses in prokaryotes. Science
ermittelt [39, 40]. Abgesehen von stabilem 5. Bosma G. C., Custer R. P., Bosma M. J. A severe com-
2007. 315: 1709-1712.
bined immunodeficiency mutation in mouse. Nature
und vollständigem Funktionsverlust mit 1983. 301: 527-530. 27. Jinek, M. et al. A programmable dual-RNA-guided
DNA endonuclease in adaptive bacterial immunity.
6. van der Burg, M. et al. A DNA-PKcs mutation in a
Cas9 lassen sich über modifizierte Versi- Science 2012. 337: 816-821.
radiosensitive T-B- SCID patient inhibits Artemis acti-
vation and nonhomologous end-joining. J Clin Invest 28. Cong, L. et al. Multiplex genome engineering using
onen von Cas9 auch Knock-down 2009. 119: 91-98. CRISPR/Cas systems. Science 2013. 339: 819-823.
(CRISPRi, wie mit siRNA) und Aktivator- 7. Murphy, E. D. and Roths, J. B. A single gene model 29. Mali, P. et al. RNA-guided human genome enginee-
for massive lymphoproliferation with immune ring via Cas9. Science 2013. 339: 823-826.
Screens (CRISPRa) durchführen [41]. Es complex disease in new mouse strain MRL. Excerpta 30. Jinek, M. et al. RNA-programmed genome editing
Medica, Amsterdam 1976. in human cells. Elife 2013. 2: e00471.
ist zu erwarten, dass in Zukunft auch in 8. Matsuzawa, A. et al. A new allele of the lpr locus, 31. Wang, H., La Russa, M. and Qi, L. S. CRISPR/Cas9
der Immunologie bald erste direkte In- lprcg, that complements the gld gene in induction of in Genome Editing and Beyond. Annu Rev Biochem
lymphadenopathy in the mouse. J Exp Med 1990. 171: 2016. 85: 227-264.
vivo-Screens, wie sie bereits im Kontext 519-531.
32. Anderson, K. V. and Nusslein-Volhard, C. Informa-
9. Fisher, G. H. et al. Dominant interfering Fas gene tion for the dorsal--ventral pattern of the Drosophila
von Hirntumoren durchgeführt wurden, mutations impair apoptosis in a human autoim- embryo is stored as maternal mRNA. Nature 1984.
mune lymphoproliferative syndrome. Cell 1995. 81: 311: 223-227.
publiziert werden. CRISPR/Cas9 hat also 935-946.
33. Whitehurst, A. W. et al. Synthetic lethal screen
auch funktionelle genetische Screens ver- 10. Rieux-Laucat, F. et al. Mutations in Fas associa- identification of chemosensitizer loci in cancer cells.
ted with human lymphoproliferative syndrome and Nature 2007. 446: 815-819.
bessert und mit vielen neuen Möglichkei- autoimmunity. Science 1995. 268: 1347-1349.
34. Gazin, C. et al. An elaborate pathway required for
11. Frank, J. et al. Exposing the human nude phenoty- Ras-mediated epigenetic silencing. Nature 2007. 449:
ten bereichert. pe. Nature 1999. 398: 473-474. 1073-1077.
12. Gordon, J. W. and Ruddle, F. H. Integration and 35. Zhou, P. et al. In vivo discovery of immunotherapy
stable germ line transmission of genes injected into targets in the tumour microenvironment. Nature
Fazit mouse pronuclei. Science 1981. 214: 1244-1246. 2014. 506: 52-57.
13. Palmiter, R. D. et al. Dramatic growth of mice that 36. Chen, R. et al. In vivo RNA interference screens
Die Immunologie hat viel von der develop from eggs microinjected with metallothion- identify regulators of antiviral CD4(+) and CD8(+) T
Manipulation von Genomen profitiert. ein-growth hormone fusion genes. Nature 1982. 300: cell differentiation. Immunity 2014. 41: 325-338.
611-615.
37. Wang, T. et al. Genetic screens in human cells using
Von spontanen und chemisch induzierten 14. Chiang, C. et al. Complex reorganization and the CRISPR-Cas9 system. Science 2014. 343: 80-84.
predominant non-homologous repair following
Mutanten über Transgene und ES-Zell- chromosomal breakage in karyotypically balanced 38. Parnas, O. et al. A Genome-wide CRISPR Screen
germline rearrangements and transgenic integration. in Primary Immune Cells to Dissect Regulatory Net-
Technologie hin zu CRISPR/Cas9 haben Nat Genet 2012. 44: 390-397, S391. works. Cell 2015. 162: 675-686.
15. Yan, B. W. et al. Mechanism of random integration 39. Patel, S. J. et al. Identification of essential genes for
sich die Möglichkeiten zur Manipulation cancer immunotherapy. Nature 2017. 548: 537-542.
of foreign DNA in transgenic mice. Transgenic Res
von Genomen verändert. Die Geschwin- 2013. 22: 983-992. 40. Manguso, R. T. et al. In vivo CRISPR screening
16. Brinster, R. L. et al. Expression of a microinjected identifies Ptpn2 as a cancer immunotherapy target.
digkeit, mit der immer neue molekulare immunoglobulin gene in the spleen of transgenic Nature 2017. 547: 413-418.
mice. Nature 1983. 306: 332-336. 41. Gilbert, L. A. et al. Genome-Scale CRISPR-Mediated
Werkzeuge, basierend auf CRISPR/Cas- Control of Gene Repression and Activation. Cell 2014.
17. Evans, M. J. and Kaufman, M. H. Establishment in
Technologie, beschrieben werden, setzt culture of pluripotential cells from mouse embryos. 159: 647-661.
Nature 1981. 292: 154-156.
der Forscherphantasie kaum noch Gren- 18. Thomas, K. R. and Capecchi, M. R. Site-directed
zen. War anfangs die Durchführung der mutagenesis by gene targeting in mouse embryo-
derived stem cells. Cell 1987. 51: 503-512.
genetischen Manipulation oft der ge- 19. Kuhn, R., Rajewsky, K. and Muller, W. Generation
and analysis of interleukin-4 deficient mice. Science
schwindigkeitsbestimmende Schritt, ist 1991. 254: 707-710.
20. Murphy, A. J. et al. Mice with megabase huma-
dies heute eher die Phänotypisierung bzw.
nization of their immunoglobulin genes generate
Dr. Johannes vom Berg, Prof. Dr. Thorsten Buch
die funktionelle Aufarbeitung von Befun- antibodies as efficiently as normal mice. Proc Natl Institut für Labortierkunde
Acad Sci U S A 2014. 111: 5153-5158. Universität Zürich, Schweiz
den aus solchen genetischen Eingriffen. 21. Gu, H. et al. Deletion of a DNA polymerase beta
gene segment in T cells using cell type-specific gene
johannes.vomberg@uzh.ch
thorsten.buch@uzh.ch
Trillium Immunologie 2018; 2(2) 101Sie können auch lesen

















































