Wachstumsvergleich zwischen gentechnisch modifizierten und natürlich mutierten Bakterien - Impuls Mittelschule
←
→
Transkription von Seiteninhalten
Wenn Ihr Browser die Seite nicht korrekt rendert, bitte, lesen Sie den Inhalt der Seite unten
Wachstumsvergleich zwischen gentechnisch
modifizierten und natürlich mutierten Bakterien
am Beispiel der Streptomycin-Resistenz
von Escherichia coli
Maturitätsarbeit von Miles Burri
Kantonsschule Hohe Promenade,
Gymnasium
Klasse 6d
Fachbereich Biologie
Zürich, Schuljahr 2017/2018
Betreuende Lehrperson: Dominique Grüter
Korreferent: Christian Bürkli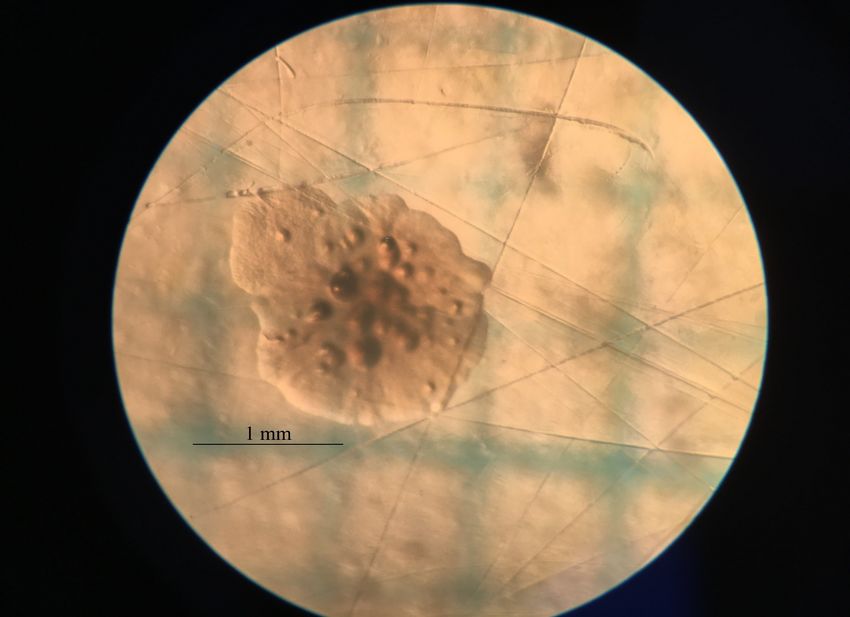
Inhaltsverzeichnis
1 Einleitung ..................................................................................................... 3
1.1 Persönliche Motivation ............................................................................................ 3
1.2 Fragestellung und Hypothese ................................................................................... 3
2 Material und Methode ................................................................................. 3
2.1 Escherichia coli ......................................................................................................... 3
2.2 Streptomycin............................................................................................................ 3
2.2.1 Wirkungsmechanismus ............................................................................................... 3
2.2.2 Streptomycin Stammlösung ........................................................................................ 4
2.3 CRISPR/Cas-Methode ............................................................................................... 4
2.3.1 Vorteile von CRISPR/Cas .............................................................................................. 5
2.3.2 Funktionsweise ........................................................................................................... 5
2.3.3 Template DNA, RNA und Cas9 im konkreten Versuch .................................................. 7
2.4 LB-Medium .............................................................................................................. 7
2.5 Utensilien ................................................................................................................. 7
2.6 Sicherheit und Sterilität............................................................................................ 7
2.7 Versuche .................................................................................................................. 8
2.7.1 Vorbereitung............................................................................................................... 8
2.7.2 Mutationsversuch ....................................................................................................... 8
2.7.3 CRISPR/Cas Versuch .................................................................................................... 8
2.7.3.1 Kompetente Bakterien herstellen ............................................................................ 8
2.7.3.2 Transformation ....................................................................................................... 9
2.7.4 Kontrollgruppe ............................................................................................................ 9
2.8 Messungen............................................................................................................... 9
3 Ergebnisse .................................................................................................. 11
3.1 Wachstumsvergleich .............................................................................................. 11
3.2 Mutationsversuch .................................................................................................. 12
3.3 CRISPR/Cas Versuch ............................................................................................... 13
3.4 Kontrollgruppe ....................................................................................................... 14
4 Diskussion .................................................................................................. 16
4.1 Grösse der Kulturen ............................................................................................... 16
4.2 Morphologie der Kolonien...................................................................................... 16
4.3 Verteilung .............................................................................................................. 17
4.4 Kontamination ....................................................................................................... 17
4.5 Wichtigkeit von CRISPR/Cas ................................................................................... 17
4.6 Signifikanz der Daten.............................................................................................. 17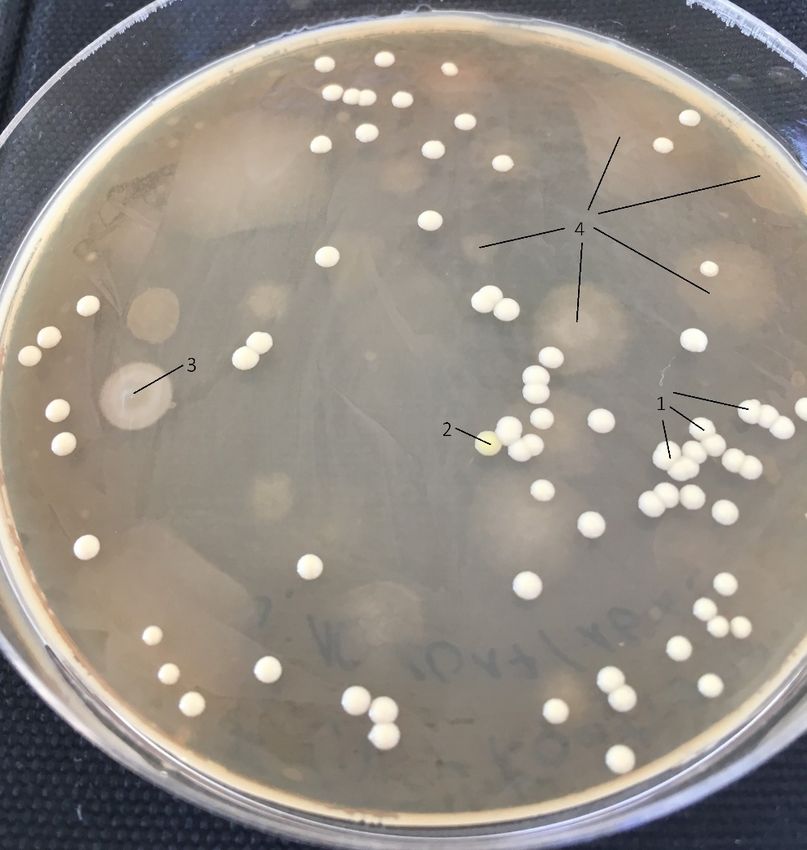
5 Zusammenfassung ..................................................................................... 18
6 Danksagung................................................................................................ 18
7 Quellenverzeichnis ..................................................................................... 19
7.1 Zeitschriften und Bücher ........................................................................................ 19
7.2 Internetseiten ........................................................................................................ 20
7.3 Abbildungen ........................................................................................................... 20
8 Glossar ....................................................................................................... 21
9 Anhang....................................................................................................... 22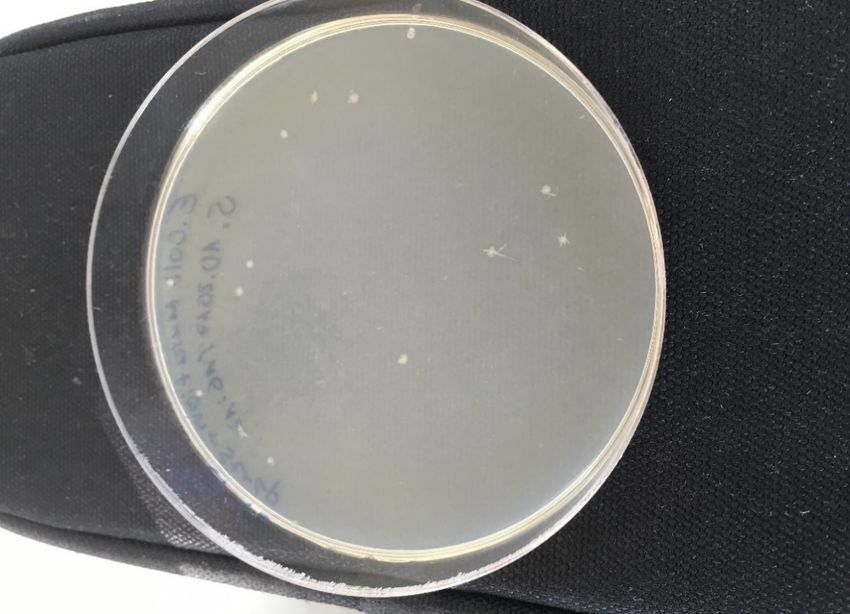
Maturitätsarbeit Miles Burri
1 Einleitung
1.1 Persönliche Motivation
Genmanipulierte Organismen geniessen in unserer Gesellschaft keineswegs einen guten Ruf.
Ein Film zu diesem Thema (Mais im Bundeshuus [1]) im Rahmen des Staatskundeunterrichts
gab mir Einsicht in die hitzige Diskussion einer Nationalratskommission, welche ein inzwischen
bis 2021 verlängertes Moratorium für den Einsatz der Technologie vorschlug. Inspiriert durch
diesen Film, recherchierte ich im Internet. Ich stiess dabei auf die Seite „The Odin“ [2], welche
„DIY Gene Engineering Kits“ anbietet. Fasziniert von der Idee, ganz ohne fortschrittliches Labor
Genversuche durchzuführen, habe ich ein solches Kit in meine Maturaarbeit integriert.
1.2 Fragestellung und Hypothese
In dieser Arbeit wird das Wachstum streptomycin-resistenter Escherichia coli verglichen. Dabei
wird zwischen Organismen verglichen, welche ihre Resistenz auf dem „natürlichen“ Weg der
zufälligen Mutation erlangt haben, und solchen, die gezielt gentechnisch modifiziert wurden.
Es wird erwartet, dass der Weg, auf welchem die Organismen ihre Resistenz erlangt haben,
keinen Einfluss auf ihr Wachstum hat. Ein möglicher Unterschied könnte jedoch darauf
beruhen, dass bei der Modifikation bei allen Organismen die gleiche, spezifische Veränderung
vorgenommen wird, während bei den natürlichen Mutanten verschiedene Veränderungen
auftreten können, welche eine Streptomycin-Resistenz hervorrufen (SPRINGER 2001). Es
existieren bis anhin jedoch keine Beweise dafür, dass diese verschiedenen Mutationen auch
unterschiedlich starke Resistenzen hervorrufen. Falls dies so wäre, müssten bei den Mutanten
Unterschiede bezüglich des Wachstums der einzelnen Kolonien sichtbar werden. In den
Messwerten wäre dies an einer grösseren Verteilung zu erkennen.
3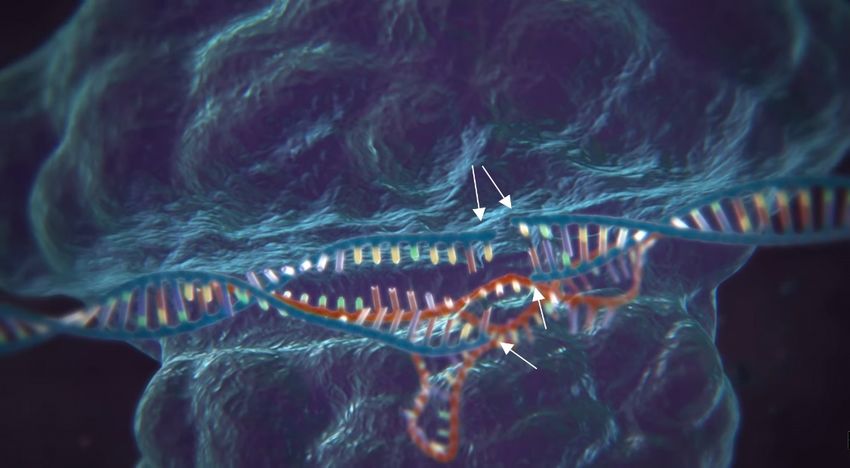
Maturitätsarbeit Miles Burri
2 Material und Methode
2.1 Escherichia coli
Für die Experimente wurde der Stamm
HME63 der Art Escherichia coli verwendet.
Erworben wurden die Bakterien in Form
einer Stichkultur von The Odin [2].
Escherichia coli eignet sich hervorragend
für Experimente im gentechnischen
Bereich, da ihr Genom weitestgehend
bekannt ist. Ein weiterer Vorteil dieses
Bakteriums liegt in seiner einfachen
Kultivierung im Labor, sowie in der kurzen
Generationszeit von 20-30 Minuten bei
optimalen Bedingungen (KNIPPERS 1971),
was das schnelle Beobachten der
Ausprägung neu eingefügter Gene Abb. 1 Escherichia coli Kolonien nach 6 Tagen Wachstum
vereinfacht. unter optimalen Bedingungen.
Wie man in der Abbildung 1 erkennt, bilden die Bakterien weisse, leicht transparente Kolonien,
welche eine kreisrunde Form mit durchgehender Aussengrenze haben. Ein weiteres Merkmal
ist die gleichmässige, konvexe Erhöhung der Kolonien.
2.2 Streptomycin
Streptomycin ist ein bakterizid wirkendes Aminoglykosid-Antibiotikum. In der Schweiz findet es
in der Humanmedizin keine Verwendung mehr. In der Veterinärmedizin wird es jedoch
weiterhin gegen bakterielle Infektionskrankheiten verwendet [3]. Auch in der Landwirtschaft
fand das Antibiotikum Verwendung, und zwar in der Bekämpfung des Feuerbrandes. Weil von
der Anwendung von Streptomycin im grossen Stil jedoch die Gefahr der Resistenzentwicklung
ausgeht, darf es seit 2016 in der Schweizer Landwirtschaft nicht mehr für die Bekämpfung des
Feuerbrandes eingesetzt werden [4]. Ein weiterer Grund für das Verbot liegt darin, dass
Streptomycin zu einer Verunreinigung von Honig führen kann. Dazu kommt es, wenn Bienen
Nektar und Pollen von Pflanzen sammeln, die mit dem Antibiotikum behandelt wurden. 2011
mussten im Thurgau deshalb über sieben Tonnen Honig vernichtet werden [5].
2.2.1 Wirkungsmechanismus
Streptomycin wirkt indem es an ein Protein der 30S-Untereinheit des bakteriellen Ribosoms
bindet. Dort verhindert das Molekül die korrekte Translation, indem es eine Fehlablesung der
mRNA verursacht. Das betroffene Bakterium kann somit keine, beziehungsweise nur noch
funktionsunfähige Proteine synthetisieren und stirbt ab. (HÖCK, FILLE 2012)
3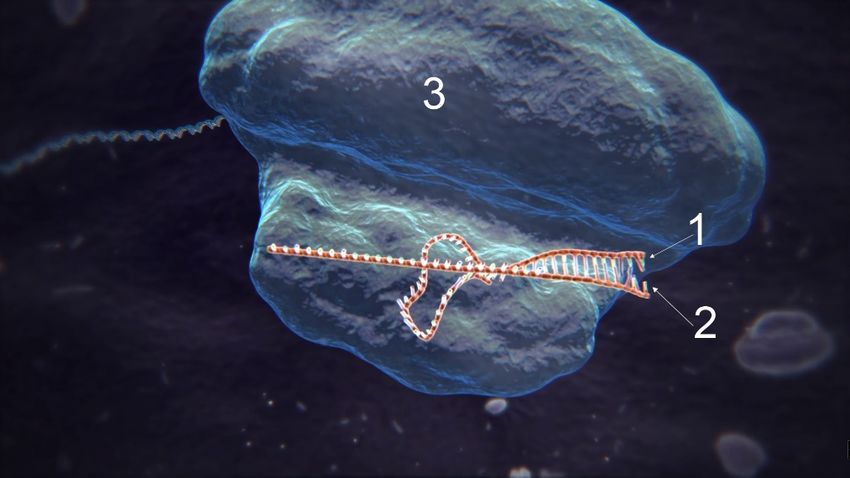
Maturitätsarbeit Miles Burri
Abb. 2 Bändermodell der Streptomycin-Bindungsstelle im Ribosom. Die gelbe Struktur ist das Streptomycin-
Molekül, orange dargestellt ist das S12-Protein.
Eine Resistenz gegen Streptomycin basiert meist auf einer Veränderung des rpsL-Gens, welches
die Information für die Synthese des S12-Proteins, eines Teils der 30S-Untereinheit, enthält.
Durch zufällige Mutation oder gezielte Manipulation kann dieses Gen so verändert werden,
dass es nun ein leicht anderes S12-Protein kodiert, an welches Streptomycin nicht mehr binden
kann. Somit kann die 30S-Untereinheit des Ribosoms, und damit auch die Proteinbiosynthese
nicht mehr vom Aminoglykosid-Antibiotikum beeinflusst werden. (TRAUB, NOMURA 1968)
Diese Resistenz tritt in der Natur durch zufällige Mutation des rpsL-Gens mit einer
Wahrscheinlichkeit von 0.3 · 10-10 bis 1.1 · 10-10 auf (NEWCOMBE, HAWIRKO 1949). Dieser Wert mag
vorerst gering erscheinen, wenn jedoch die oftmals grosse Anzahl der Bakterien berücksichtigt
wird, ist das absolute Vorkommen der Resistenz nicht sehr selten. Im menschlichen Stuhl
beispielsweise kommen im einem Gramm 108-109 E. coli Bakterien vor (SCHLEGEL 2007).
Statistisch gesehen müsste also pro 10 Gramm Stuhl ein streptomycin-resistentes Bakterium
vorhanden sein. Gemäss STENDERUP (1953) kann eine schrittweise Erhöhung der Streptomycin-
Konzentration die Wahrscheinlichkeit erhöhen, dass in E. coli Resistenzen auftreten.
2.2.2 Streptomycin Stammlösung
Für die Selektion der erfolgreich transformierten und mutierten Bakterien wurden diese dem
Aminoglykosid-Antibiotikum ausgesetzt. Es wurde dazu eine Streptomycin-Stammlösung mit
einer Konzentration von 100 mg/mL hergestellt, welche dann in gewünschter Menge den
Nährmedien hinzugefügt werden konnte.
2.3 CRISPR/Cas-Methode
Die CRISPR/Cas-Methode ist ein Werkzeug zum Verändern der DNA, basierend auf einem
Immunsystem ähnlichen, antiviralen Mechanismus der in Prokaryoten vorkommt. Ein
Bakterium kann nach dem erstmaligen Kontakt mit einer fremden, viralen DNA eine kurze
Sequenz jener in seine eigene DNA einbauen. Diese Sequenz - genannt „Spacer“ - ermöglicht
es dem Bakterium, bei erneutem Kontakt mit derselben viralen DNA diese zu erkennen und zu
4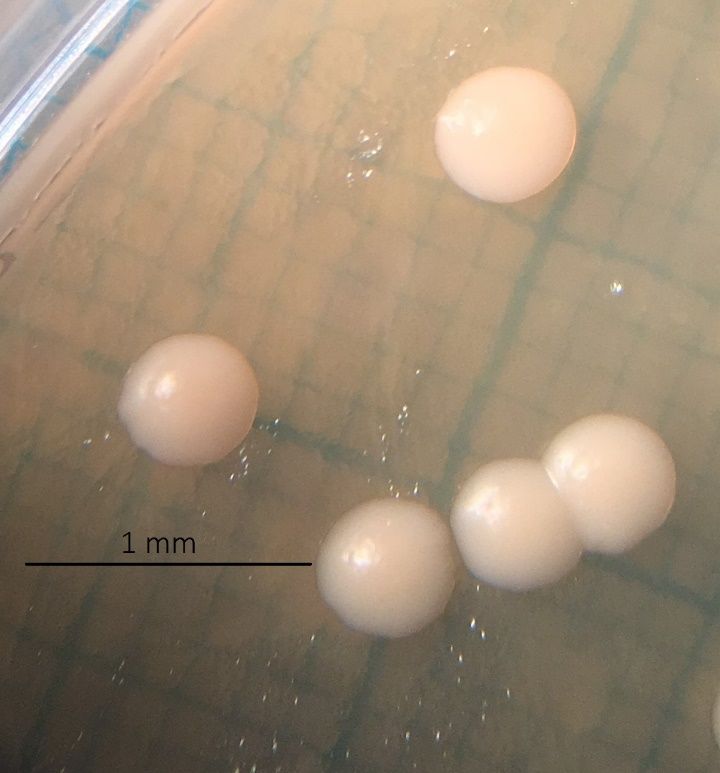
Maturitätsarbeit Miles Burri
zerschneiden. Das Zerschneiden deaktiviert die fremde DNA und macht sie somit ungefährlich
für den Organismus.
Diese Spacer-Sequenzen kann ein Bakterium mehrmals und für verschiedene Viren anfertigen,
wobei die verschiedenen fremden Sequenzen in der eigenen DNA stets von einer sich
wiederholenden Sequenz getrennt werden. Solche Abschnitte von sich abwechselnden
fremden und sich wiederholenden Sequenzen in der prokaryotischen DNA heissen auf Englisch
Clustered Regularly Interspaced Short Palindromic Repeats, kurz CRISPR.
2.3.1 Vorteile von CRISPR/Cas
Die wichtigsten Vorteile dieser Methode gegenüber anderen Möglichkeiten zur
gentechnischen Veränderung sind die Einfachheit der Arbeitsschritte und die hohe Effizienz,
mit welcher Gene eingefügt werden können. Auch die Möglichkeit, mit nur wenigen, kurzen
RNA-Molekülen präzise Schnittstellen zu definieren ist von Vorteil (STERNBERG, DOUDNA 2015),
ebenso wie der grosse Anwendungsbereich der Methode, denn mittels CRISPR/Cas können
sogar Stammzellen sowie befruchtete Embryos gentechnisch modifiziert werden. Ein weiterer
wichtiger Vorteil ist der geringe Preis der Ausgangsmoleküle.
2.3.2 Funktionsweise
Das System besteht im Grundsatz aus vier Komponenten: der Endonuklease Cas9, der CRISPR
RNA (crRNA), der trans-activating CRISPR RNA (tracrRNA) und der Template DNA. Das Cas9-
Protein ist das eigentliche «Werkzeug» dieser Methode. Es ist in der Lage, die DNA an einer
präzisen Stelle doppelsträngig zu zerschneiden und so die Möglichkeit zu schaffen,
Veränderungen vorzunehmen. Die crRNA fungiert als eine Art „GPS“. Sie kann das Cas9-Protein
an eine exakte Stelle der DNA lotsen, die zerschnitten werden soll. Dies geschieht mithilfe eines
RNA-Abschnittes, welcher eine kurze Kopie der Ziel-DNA enthält und somit an jene binden
kann. Damit dies geschehen kann, bricht das Cas9-Protein in der Ziel-DNA die
Wasserstoffbrücken von etwa 20 Basenpaaren auf. Nun kann-die crRNA mit ihrer Kopie der
Ziel-DNA an diese binden. Die tracrRNA hat dabei die Aufgabe, Cas9 und crRNA zu verbinden
und somit einen funktionsfähigen Komplex zu bilden.
Sobald dieser Komplex sein Ziel gefunden hat und an die DNA gebunden ist, zerschneidet das
Cas9-Enzym die DNA doppelsträngig. Die Zelle versucht daraufhin sofort ihre DNA wieder zu
reparieren, und zwar mittels homologer Rekombination. Bei dieser Reparation können
Veränderungen an der DNA vorgenommen werden. Denn um ihre DNA zu reparieren, braucht
die Zelle eine Vorlage, vorgesehen dafür wäre das jeweils homologe Chromosom. Wenn die
Zelle jetzt jedoch mit einer DNA-Sequenz geflutet wird, die bis auf die gewünschte Modifikation
genau der zerschnittenen Stelle entspricht (genannt Template DNA, Engl. für Vorlage-DNA),
wird sie in einigen Fällen als Vorlage bei der Reparation genommen. Die Zelle baut die
Modifikation somit selber in ihre eigene DNA ein. (CHARPENTIER, DOUDNA 2014)
5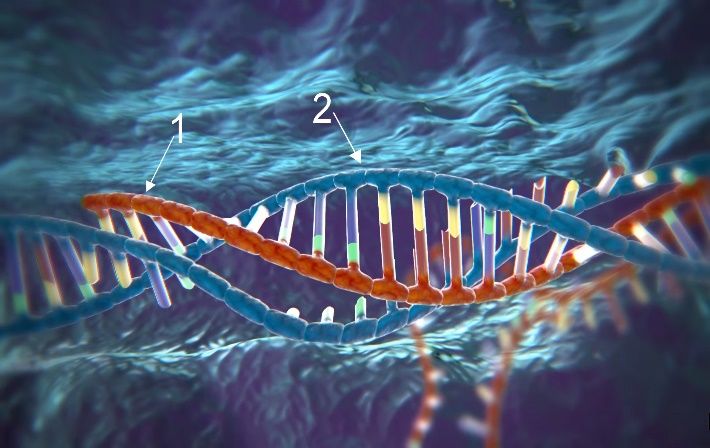
Maturitätsarbeit Miles Burri
1. Funktionsfähiger Komplex bestehend aus tracrRNA (1), 2. Kopie der Ziel-DNA (1, rot) und schematisch dargestellt
crRNA (2) und Cas9 (3). die molekularen Scheren von Cas9 (2).
3. Die crRNA (1) bindet an die aufgebrochene Ziel-DNA (2). 4. Cas9 zerschneidet die Ziel-DNA doppelsträngig.
5. Die Zelle beginnt mit der homologen Rekombination zur 6. In einigen Fällen wird die Modifikation von der Template-
Reparatur der zerschnittenen DNA. DNA auf die Ziel-DNA kopiert.
Abb. 3 Ablauf der verschiedenen Prozesse bei einer gentechnischen Modifikation mit CRISP/Cas.
6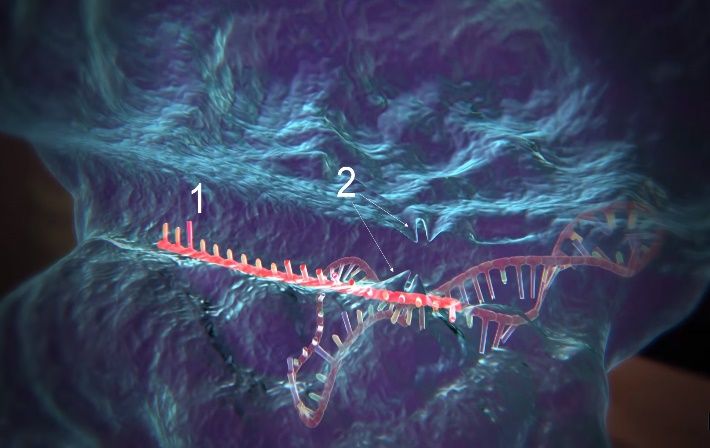
Maturitätsarbeit Miles Burri
2.3.3 Template DNA, RNA und Cas9 im konkreten Versuch
Die Template DNA, die crRNA und tracrRNA
sowie das Cas9-Protein wurden allesamt von
The Odin [2] in Form eines „Bacterial-
Engineering-Kits“ erworben.
Es folgt die in der crRNA enthaltene Sequenz,
welche genau komplementär zur Schnittstelle
im rpsL-Gen ist. Im Folgenden wird jede
Nukleinbase durch einen Buchstaben
dargestellt. (A = Adenin, C = Cytosin, G =
Guanin, T = Thymin) [7]
TACTTTACGCAGCGCGGAGTTCGGTTTTTT
Die folgende Sequenz bildet einen Strang der
Template DNA. Rot markiert ist die einzige
Abb. 4 Darstellung des genetischen Codes. Drei Basen
Base, welche sich vom Wildtyp unterscheidet kodieren jeweils eine Aminosäure. In unserem Fall wird
(anstelle von Cytosin (C) wäre dort ein Adenin anstelle eines Lysins (AAA) ein Threonin (ACA) kodiert.
(A) vorhanden). Wie man in der Abbildung 4 an den roten Pfeilen erkennen kann, führt dieser
Basenwechsel dazu, dass anstelle eines Lysins ein Threonin kodiert wird. [7]
TATGAAATGCGACGCGCCTCAAGCCAAAACATCCTCACCATCATATATGTGCTCATGTA
Diese Mutation des rpsL-Gens wird K42T genannt, da die 42. Aminosäure des kodierten
Proteins von einem Lysin (K) zu einem Threonin (T) mutiert wird. Das Resultat der Mutation ist
eine Resistenz gegen Streptomycin (JIANG 2013).
2.4 LB-Medium
Zur Kultivierung der Bakterien wurde LB-Medium (vom Englischen «Lysogeny Broth») in zwei
Formen verwendet. Zum einen als flüssiges Nährmedium, zum anderen zusammen mit Agar-
Agar als festes Nährmedium in einer Petrischale. Das flüssige LB-Medium wurde bei Faust
Laborbedarf AG erworben, das Agar-Medium bei The Odin.
2.5 Utensilien
Die meisten Instrumente und Behältnisse waren im Kit von The Odin inbegriffen, lediglich die
L-förmigen Ausplattierrechen sowie die 10ml Röhrchen mussten zusätzlich erworben werden.
(KS Hohe Promenade Biologie bzw. Faust Laborbedarf AG)
2.6 Sicherheit und Sterilität
Der verwendete Stamm von E. coli ist nicht pathogen. Trotzdem wurde, weitestgehend mit
Atemschutz und Nitrilhandschuhen gearbeitet. Die Arbeitsflächen wurden vor und nach dem
Arbeiten mit 99.99% Ethanol desinfiziert. Gearbeitet wurde immer neben einem brennenden
Bunsenbrenner, da so ein aufwärtsgerichteter Luftstrom erzeugt wird, welcher die Sterilität
verbessert.
7Maturitätsarbeit Miles Burri
2.7 Versuche
2.7.1 Vorbereitung
Für die folgenden Versuche wurde jeweils 48h vor der Durchführung eine LB-Agarplatte mit
einer Impföse E. coli HME63 aus einer Stichkultur angeimpft. Die Platte wurde danach mit dem
Deckel nach unten bis zu ihrem Gebrauch bei Raumtemperatur und unter Abschluss von Licht
inkubiert. Die Inkubation mit dem Deckel nach unten verhindert bei Agarplatten das
Heruntertropfen von Kondensationswasser auf die Bakterienkultur.
2.7.2 Mutationsversuch
Die empfohlene Endkonzentration für die erfolgreiche Selektion streptomycin-resistenter
Bakterien beträgt 100 µg/mL [6]. Diese wurde durch zweimalige Zugabe von 5 µL Streptomycin-
Stammlösung erreicht. Durch die zweistufige Erhöhung der Konzentration wird die
Wahrscheinlichkeit für Resistenzentwicklung erhöht (siehe Kapitel 2.2).
Es wurden 10 mL flüssiges LB-Medium mit einer Impföse voll E. coli HME63 beimpft. Nach 20
Stunden bei Raumtemperatur wurde ein erstes Mal 5 µL Streptomycin-Stammlösung
hinzugefügt. Dies wurde nach 20 Stunden nochmals wiederholt. Bei einer jetzigen
Konzentration von 100 µg/mL wurden die Bakterien über Nacht bei Raumtemperatur inkubiert.
Danach wurden 200 µL dieser Lösung auf eine Agarplatte mit 100 µg/mL Streptomycin
pipettiert und anschliessend mit einem Ausplattierrechen verteilt. Die Platte wurde danach für
6 Tage bei 37 °C in einem Wärmeschrank an der KS Hohe Promenade inkubiert.
2.7.3 CRISPR/Cas Versuch
Dieser Versuch basiert auf einer Anleitung von The Odin [7].
2.7.3.1 Kompetente Bakterien herstellen
Als kompetent werden in der Mikrobiologie Zellen bezeichnet, die die Fähigkeit besitzen, DNA
aus ihrem Umfeld aufzunehmen. Da E. coli nicht zu den Bakterien gehört, welche von Natur aus
kompetent sind, muss künstlich eine Kompetenz erreicht werden (CHEN, DUBNAU 2004). Dies
geschieht chemisch durch Zugabe einer bakteriellen Transformationsmischung bestehend aus
10% Polyehylenglycol (PEG), 5% Dimethylsulfoxid (DMSO) und 25mM Calciumchlorid (CaCl2).
Die genaue Funktion dieser Stoffe bei der Herstellung kompetenter Zellen ist noch nicht völlig
verstanden. Von PEG und CaCl2 wird im Allgemeinen jedoch angenommen, dass sie die negative
Ladung der DNA neutralisieren und so ein einfacheres Eindringen dieser durch die ebenfalls
negativ geladene Plasmamembran ermöglichen. Es konnten jedoch keine beweisenden Studien
dazu gefunden werden. DMSO erhöht gemäss NOTHAM (2006) die Permeabilität von
Lipiddoppelschichten. Temperaturwechsel erhöhen ebenfalls die Chance, dass DNA passiv in
die Zelle aufgenommen wird. Der Mechanismus dahinter ist jedoch nicht geklärt.
Für die Herstellung der kompetenten Zellen wurden 200 µL Transformationsmischung in ein
Eppendorf-Röhrchen gegeben. Hinzugegeben wurde eine Impföse voll E. coli HME63. Der
8Maturitätsarbeit Miles Burri
Behälter wurde danach so lange von Hand geschüttelt, bis die Bakterien homogen in der Lösung
verteilt waren.
2.7.3.2 Transformation
Für die Transformation der nun kompetenten Bakterien wurden demselben Eppendorf-
Röhrchen 10 µL Cas9+tracrRNA (100 ng/µL), 10 µL crRNA (100 ng/µL) und 10 µL Template DNA
(100 ng/µL) zugegeben. Die Probe wurde dann für 30 Minuten im Kühlschrank bei 4°C inkubiert.
Danach wurde das Röhrchen für 30 Sekunden einem Hitzeschock bei 42°C unterzogen. Zum
Schluss wurden noch 800 µL flüssiges LB-Medium mit Raumtemperatur hinzugegeben,
anschliessend wurde die Probe für 2.5 Stunden bei 37°C inkubiert.
Nach dieser Inkubationszeit wurden 200 µL der Kultur auf eine Agarplatte mit 100 µg/mL
Streptomycin gegeben und mit einem Ausplattierrechen verteilt. Die Platte wurde danach für
6 Tage bei 37 °C im Wärmeschrank inkubiert.
2.7.4 Kontrollgruppe
Die Daten einer unbehandelten Kontrollgruppe wurden ebenfalls erhoben. Dazu wurden 10 mL
flüssiges LB-Medium mit einer Kolonie E. coli HME63 beimpft. Diese Flüssigkultur wurde
anschliessend für 12 Stunden bei Raumtemperatur inkubiert. Danach wurden 200 µL dieser
Kultur auf eine Agarplatte ohne Streptomycin gegeben und für 6 Tage bei 37 °C inkubiert.
2.8 Messungen
Gemessen wurden nach 6 Tagen Inkubationszeit die Durchmesser der einzelnen
koloniebildenden Einheiten (KbE), sowie ihre Anzahl auf jeder Agarplatte. Dazu wurden die
Agarplatten aus dem Wärmeschrank genommen und unter dem Binokular fotografiert.
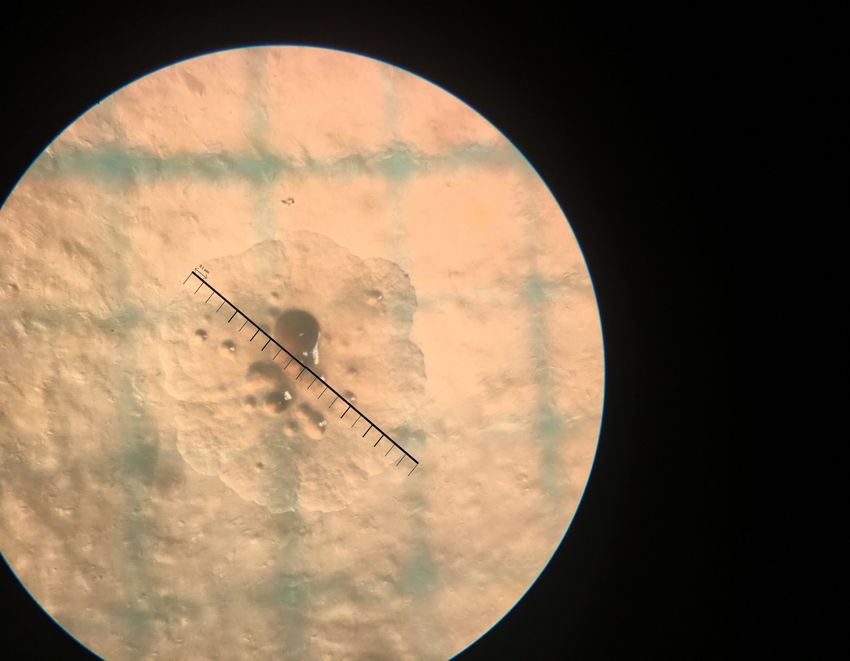
Abb. 5 Beispiel einer Messung. Die abgebildete Kolonie weist einen Durchmesser von 1.9 mm auf.
9Maturitätsarbeit Miles Burri
Es war vorgesehen, die Kolonien direkt unter dem Binokular und einem unter den Agarplatten
liegenden Millimeterpapier zu vermessen. Aufgrund des geringen Wachstums der Kulturen
wurden sie jedoch digital in Photoshop vermessen, indem das Millimeterpapier als
Längenreferenz genommen wurde. Ein digitaler Massstab mit 10 Teilabständen wurde dann
auf genau einen Millimeter Länge gebracht und über das Foto der Kolonie gezogen. So konnte
dessen Durchmesser auf 0.1 mm genau bestimmt werden.
10Maturitätsarbeit Miles Burri
3 Ergebnisse
3.1 Wachstumsvergleich
Vergleich der Mittelwerte
2.50
2.27
2.00
Durchmesser in mm
1.50
1.60
1.00
1.03
0.50
0.00
CRISPR/Cas Mutation Kontrollgruppe
Abb. 6 Vergleich der Durchmesser-Mittelwerte der drei Versuche.
Abbildung 6 zeigt die Mittelwerte der drei Versuchsreihen in Bezug zueinander. Klar zu
erkennen ist, dass die unbehandelten Kolonien der Kontrollgruppe, welche ohne Streptomycin
wachsen konnten mit einem Durchschnitt von 2.27 mm Durchmesser am grössten wurden.
Gefolgt werden sie von den natürlichen Mutanten, dessen mittlerer Durchmesser 1.60 mm
betrug. Am wenigsten gut wuchsen die mittels CRISPR/Cas transformierten Bakterien. Die
Kolonien wurden hier im Mittel lediglich 1.03 mm gross.
11Maturitätsarbeit Miles Burri
3.2 Mutationsversuch
Häufigkeitsverteilung natürliche Mutation
6
5
Häufigkeit
4
3
2
1
0
0.4 0.6 0.8 1 1.2 1.4 1.6 1.8 2 2.2 2.4 2.6 2.8 3
Durchmesser in mm
Abb. 7 Häufigkeitsverteilung des Durchmessers der Kolonien der natürlichen Mutanten.
Abbildung 7 zeigt die Häufigkeitsverteilung der Durchmesser der KbE des natürlichen
Mutationsversuchs. Zu beobachten ist eine eher schmale Verteilung, mit einem Minimalwert
von 1.3 mm und einem Maximalwert bei 1.9 mm. Das Dichtemittel befindet sich bei 1.5 mm,
der Median bei 1.6 mm und der arithmetische Mittelwert beträgt ebenfalls 1.6 mm. Die
Standardabweichung hat einen Wert von ± 0.167 mm. Zur Zeit der Messung befanden sich 15
KbE auf der Agarplatte, was dem geringsten Wert der drei Versuche entspricht.
Abb. 8 Kolonie der natürlichen Mutanten nach 6 Tagen Wachstum auf streptomycinhaltigem Agar.
Von Auge betrachtet erschienen alle Kolonien weiss, während sie durch das Binokular
betrachtet nahezu farblos und durchsichtig erschienen. Zu beobachten war bei allen Kolonien
die Bildung glatter, rundlich aus der Mitte der Kolonie herausragender Verdickungen sowie
unregelmässige Aussengrenzen (siehe Abb. 8).
12Maturitätsarbeit Miles Burri
3.3 CRISPR/Cas Versuch
Häufigkeitsverteilung CRISPR
8
7
6
5
Häufigkeit
4
3
2
1
0
0.4 0.6 0.8 1 1.2 1.4 1.6 1.8 2 2.2 2.4 2.6 2.8 3
Durchmesser in mm
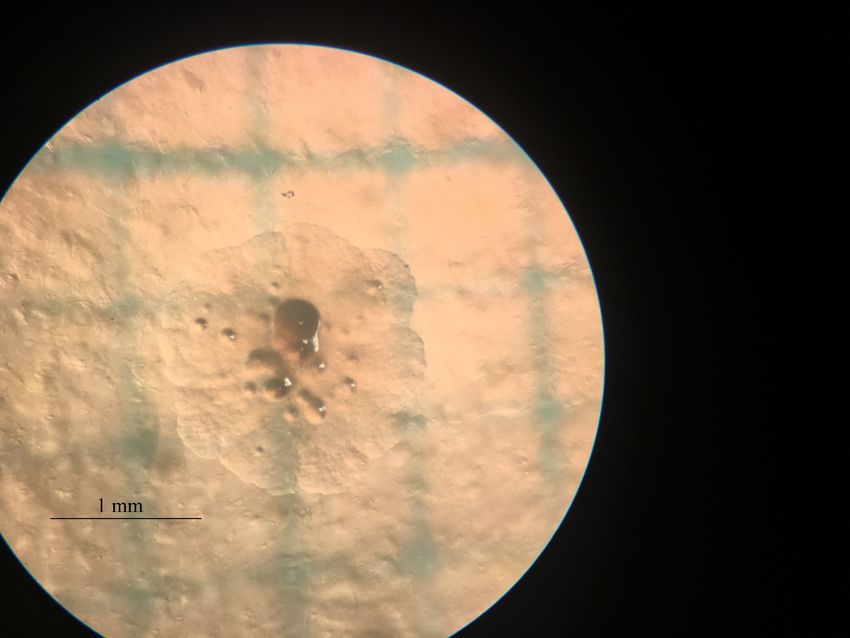
Abb. 9 Häufigkeitsverteilung der Durchmesser des mittels CRISPR/Cas transformierten E. coli.
Auf Abbildung 9 erkennt man die Häufigkeitsverteilung der Durchmesser der KbE des
CRISPR/Cas Versuchs. Mit einem Minimalwert von 0.4 mm und einem Maximalwert von 2.0
mm liegt eine eher breite Verteilung vor. Dies zeigt sich auch an der Standardabweichung von
± 0.343 mm. Der Mittelwert der Datenreihe beträgt 1.03 mm, der Median hat einen Wert von
1.0 mm. Bei diesem Versuch waren nach 6 Stunden Wachstum 39 KbE auf der Agarplatte
vorhanden.
Abb. 10 Kolonie der mittels CRISPR/Cas transformierten E. coli nach 6 Tagen Wachstum.
Vor dem blossen Auge erschienen die Kolonien weiss, unter dem Binokular jedoch hatten sie
eine nahezu farblose Erscheinung. Die Bildung der rundlichen Verdickungen war auch hier bei
allen Kolonien zu beobachten. Unregelmässige Aussengrenzen traten ebenfalls bei allen
Kolonien auf (siehe Abb. 10).
13Maturitätsarbeit Miles Burri
3.4 Kontrollgruppe
Häufigkeitsverteilung
Wachstum ohne Streptomycin
16
14
12
Häufigkeit
10
8
6
4
2
0
0.4 0.6 0.8 1 1.2 1.4 1.6 1.8 2 2.2 2.4 2.6 2.8 3
Durchmesser in mm
Abb. 11 Häufigkeitsverteilung der Durchmesser der Kontroll-Kolonien.
Der Abbildung 11 ist die Häufigkeitsverteilung der Durchmesser der KbE des Kontrollversuchs
zu entnehmen. Mit einem Minimum von 1.7 mm und einem Maximum bei 2.6 mm ist eine
mittelmässige Verteilung zu beobachten. Die Standardabweichung beträgt ± 0.191 mm. Der
Median beträgt 2.3 mm, das arithmetische Mittel 2.27 mm.
Abb. 12 Rechts die Agarplatte ohne Streptomycin, 6 Tage nach der Inokulation mit E. coli. Zum Vergleich links der
gleiche Versuch mit Streptomycin.
Wie auf der Abbildung 12 zu erkennen ist, wurde die Agarplatte ohne Streptomycin
kontaminiert. Die regelmässigen, runden Kolonien mit weisser Farbe (1) sind die E. coli
Kolonien. Sie sind die Einzigen, die für die Datenerhebung berücksichtigt wurden. Zu erkennen
sind aber viele andere Kolonien. Eine (2) teilt bis auf ihre gelbliche Farbe das Erscheinungsbild
mit den E. coli Kolonien. Auf der Oberfläche des Agars wurde eine weitere unbekannte Kolonie
14Maturitätsarbeit Miles Burri
beobachtet (3) sowie mehrere, grössere Kolonien, welche nicht auf der Oberfläche, sondern im
Agar zu wachsen schienen (4).
Links ist die Platte mit Zugabe von Streptomycin abgebildet. Zu erkennen ist nur das Wachstum
einiger weniger, resistenter Kolonien.
15Maturitätsarbeit Miles Burri
4 Diskussion
4.1 Grösse der Kulturen
Der Fakt, dass die streptomycin-resistenten Bakterien deutlich weniger schnell wuchsen als die
unbehandelte Kontrollgruppe legt nahe, dass die K42T-Mutation des rpsL-Gens negative
Auswirkungen auf die Proteinbiosynthese hat. Dies widerspricht einer Studie von
CHUMPOLKULWONG ET AL. (2004), welche für diese Mutation eine höhere Proteinproduktion sowie
eine höhere Genauigkeit bei der Translation gegenüber dem Wildtyp beobachtete. Jene Studie
beruht jedoch auf Beobachtungen bei der zellfreien Genexpression, während in dieser Arbeit
lebende Zellen untersucht wurden.
In einer Studie von PAULANDER ET AL. (2009) wurde festgestellt, dass zwei ähnliche Mutationen
(K42N, K42R) bei Salmonella Typhimurium ebenfalls eine Beeinträchtigung des Wachstums
verursachen. Interessanterweise wurde in jener Studie jedoch herausgefunden, dass diese
Mutationen den Bakterien in einem kohlenstoffarmen Milieu einen Vorteil gegenüber dem
Wildtyp verschaffen, was sich in einem schnelleren Wachstum zeigte. Dies lässt die Frage offen,
ob auch die in dieser Arbeit erzielte K42T-Mutation von E. coli unter suboptimalen
Wachstumsbedingungen einen Vorteil gegenüber dem Wildtyp schaffen würde. Die
Untersuchung dieser Frage konnte in dieser Arbeit jedoch nicht aufgegriffen werden, lässt aber
Raum für weitere Arbeiten in der Zukunft.
In dieser Arbeit wurde weiter festgestellt, dass die mittels CRISPR/Cas modifizierten Bakterien
weniger wuchsen als die natürlichen Mutanten. Dies könnte damit begründet werden, dass der
Hitzeschock sowie die Transformationsmischung eine Belastung für die Bakterien darstellten.
Dieser zusätzlichen Belastung sollte mit den 2.5 Stunden Inkubation zwischen dem Beenden
der Transformation und dem Animpfen der Agarplatte entgegengewirkt werden. Die
vorgesehenen 2.5 Stunden könnten jedoch nicht genug Zeit für die Bakterien gewesen sein,
sich vollständig zu erholen.
Ein weiterer Grund für den beobachteten Unterschied könnte darin liegen, dass die DNA der
transformierten Bakterien zuerst repariert werden muss, bevor sich die Organismen teilen
können. Zudem können bei diesen Reparaturen durchaus Fehler entstehen. Genau diese
Anfälligkeit für Fehler bei der Reparatur der DNA wurde bei der Transformation mittels
CRISPR/Cas ausgenutzt. Es kann also nicht ausgeschlossen werden, dass weitere, ungewollte
Fehler in die DNA der Zelle eingebaut werden, welche einen negativen Einfluss auf das
Wachstum hatten.
4.2 Morphologie der Kolonien
Das Aussehen der Kolonien der Mutanten und der modifizierten Bakterien war nahezu
identisch. Der drastische Unterschied dieser gegenüber den Kolonien der Kontrollgruppe ist
jedoch sehr interessant. Farbe und Form der unter Einfluss von Streptomycin gewachsenen
Kolonien unterschieden sich grundlegend von den Attributen einer klassischen E. coli-Kolonie,
wie wir sie in der Kontrollgruppe beobachten konnten. Es kann nicht völlig ausgeschlossen
16Maturitätsarbeit Miles Burri
werden, dass auf den mit Streptomycin angereicherten Agarplatten gar keine E. coli Kolonien,
sondern fremde Bakterien vorhanden waren. Sehr unwahrscheinlich erscheint jedoch, dass bei
beiden Versuchen die gleichen Kontaminanten aufgetreten sind, da die Versuche zeitlich
versetzt und mit unterschiedlichen Instrumenten durchgeführt wurden.
Das auffallend andere Erscheinungsbild der resistenten Kolonien ist jedoch wahrscheinlich -
analog zur geringeren Wachstumsrate - auf die Einflüsse der Mutation auf die
Proteinbiosynthese zurückzuführen.
4.3 Verteilung
Interessant war die Widerlegung der Hypothese, dass die Werte der natürlich mutierten
Bakterien über ein grösseres Spektrum verteilt sein sollten. Genau das Gegenteil war der Fall.
Dies legt nahe, dass die verschiedenen, zu einer Streptomycin-Resistenz führenden Mutationen
ähnlich starke Resistenzen hervorrufen, was bei den Mutanten in einem sehr konstanten
Wachstum der KbE resultierte.
4.4 Kontamination
Wann und wie die Agarplatte kontaminiert wurde, konnte nicht bestimmt werden. Mit diesem
Risiko muss jedoch gerechnet werden, da sich immer Bakterien aus der Luft auf dem Agar
absetzen können, wenn nicht in einem vollkommen sterilen Labor gearbeitet wird.
Dass eine Kontamination nur bei den Agarplatten ohne Streptomycin erfolgte, kann als Hinweis
dafür interpretiert werden, dass das Streptomycin auch tatsächlich wirkte.
4.5 Wichtigkeit von CRISPR/Cas
Es mag auf den ersten Blick unnötig scheinen, den Aufwand und die Kosten in Kauf zu nehmen,
eine Modifikation mittels CRISPR/Cas durchzuführen, zumal die natürliche Mutation ähnlich
erfolgreich war. Es muss an dieser Stelle klargestellt werden, dass dieser Versuch lediglich um
die Untersuchung einer Antibiotikaresistenz bemüht war, welche auch in der Natur relativ
häufig vorkommt. Die enorme Wichtigkeit von CRISPR/Cas in der Forschung liegt jedoch darin,
dass nahezu jede mögliche Sequenz eingefügt werden kann, unabhängig davon ob sie in der
Natur existiert oder nicht.
In der Krebsforschung werden zum Beispiel T-Zellen des Immunsystems von Patienten mittels
CRISPR/Cas so verändert, dass sie besser an Tumorzellen binden und diese somit vernichten
können. Das Verfahren hat in diversen klinischen Studien bei bis zu 90% der Patienten zu einem
völligen Verschwinden der Symptome geführt [8].
4.6 Signifikanz der Daten
Aufgrund der nur einmaligen Durchführung der Versuche und der damit verbundenen geringen
Zahl der erhobenen Rohdaten, kann nicht von repräsentativen Ergebnissen gesprochen
werden. Es war ursprünglich geplant, jeden Versuch drei Mal durchzuführen. Dies war aber
wegen der geringen Menge an Ausgangsmolekülen für den CRISPR/Cas-Versuch schliesslich
nicht möglich.
17Maturitätsarbeit Miles Burri
5 Zusammenfassung
In dieser Arbeit wurde das Wachstum streptomycin-resistenter Escherichia coli verglichen.
Dazu wurden auf zwei Arten resistente Bakterien erhalten. Zum einen wurde die gentechnische
CRISPR/Cas-Methode angewendet, zum anderen wurden durch den Prozess der Selektion
natürliche Mutanten ausfindig gemacht. Das Wachstum dieser resistenten Bakterien wurde
anschliessend untereinander und mit einer unbehandelten Kontrollgruppe verglichen.
Festzustellen war, dass die mutierten Organismen besser wuchsen als die gentechnisch
manipulierten, sowie dass das Wachstum der resistenten Bakterien unter dem der
Kontrollgruppe lag. Interessant war auch die Morphologie der resistenten Kolonien, welche sich
stark von jener des Wildtyps unterschied.
Die Arbeit bietet mit dem natürlichen Mutationsversuch auch ein beängstigendes Beispiel
dafür, wie schnell Bakterien eine Resistenz gegen ein Antibiotikum erlangen können. Nachdem
die Bakterien nur 40 Stunden lang dem Streptomycin ausgesetzt waren, konnten bereits einige
resistente Kolonien ausselektioniert werden. Es wird somit gut verständlich, weshalb
Antibiotika nicht im grossen Stil, sondern nur sehr selektiv angewendet werden sollen.
6 Danksagung
An dieser Stelle möchte ich mich bei allen bedanken, die mich beim Durchführen der Versuche
unterstützt und motiviert haben und beim Verfassen dieser Arbeit begleiteten.
Allen voran ein grosses Merci an Herr Dominique Grüter, der diese Arbeit fachlich betreut und
begutachtet hat. Er hat mich sehr beim Finden einer interessanten Fragestellung und bei der
Planung der Versuche unterstützt. Vor allem als die Materialien für die Versuche nicht eintrafen
und ernsthafte Probleme in Sicht kamen, konnte ich auf seine Hilfe zählen.
Mein Dank gebührt auch Frau Sibylle Schwarz-Scheidegger und Frau Pascale-Anne
Sommerhalder-Molinari, den beiden Biologieassistentinnen. Sie stellten mir jegliche
Instrumente für die Versuche bereit und waren sehr hilfsbereit, wenn ich Probleme oder
Unsicherheiten hatte.
Ebenfalls danken möchte ich meinen Eltern, sowohl für ihre Unterstützung bei der Beschaffung
der Versuchsmaterialien als auch für das mehrmalige Gegenlesen meiner Arbeit.
Diese Arbeit wäre ohne all diese Unterstützung niemals möglich gewesen, vielen Dank!
18Maturitätsarbeit Miles Burri
7 Quellenverzeichnis
7.1 Zeitschriften und Bücher
Charpentier E, Doudna J. (2014): The new frontier of genome engineering with CRISPR-Cas9.
In: Nature, 28:346.
Chen I, Dubnau D. (2004): DNA uptake during bacterial transformation. In: Nature Reviews
Microbiology, 2:241.
Chumpolkulwong N, et al. (2004): Effects of Escherichia coli ribosomal protein S12 mutations
on cell-free protein synthesis. In: European Journal of Biochemistry, 6:271.
Höck M, Fille M. (2012): Aminoglykoside. In: Medizinische Mikrobiologie und Infektiologie.
Springer Verlag.
Jiang W, et al. (2013): CRISPR-Assisted Editing of Bacterial Genomes. In: Nature biotechnology,
31:233.
Knippers R. (1971): Vermehrung von Bakterien. In: Molekulare Genetik. Georg Thieme Verlag.
Notham R, et al. (2006): Molecular Basis for Dimethylsulfoxide (DMSO) Action on Lipid
Membranes. In: Journal of the American Chemical Society, 128.
Paulander W, et al. (2009): The Fitness Cost of Streptomycin Resistance Depends on rpsL
Mutation, Carbon Source and RpoS (σS). In: Genetics, 2:183.
Schlegel H. (2007): Prokaryoten. In: Allgemeine Mikrobiologie. Georg Thieme Verlag
Springer B, et al. (2001): Mechanisms of Streptomycin Resistance: Selection of Mutations in the
16S rRNA Gene Conferring Resistance. In: Antimicrobial Agents Chemotherapy,
45:10.
Stenderup A. (1953): Development of Streptomycin resistance in E. coli. In: Acta Pathologica
Microbiologica Scandinavica, 33:2.
Sternberg S, Doudna J. (2015): Expanding the Biologist’s Toolkit with CRISPR-Cas9. In: Molecular
Cell, 58:4.
Traub P, Nomura M. (1968): Streptomycin resistance mutation in Escherichia coli: altered
ribosomal protein. In: Science, 12:160.
19Maturitätsarbeit Miles Burri
7.2 Internetseiten
[1] https://www.youtube.com/watch?v=uaz1wx6gt9g Abgerufen am: 22.10.2017
[2] www.the-odin.com Abgerufen am: 20.07.2017
[3] http://www.pharmawiki.ch/wiki/index.php?wiki=Streptomycin
Abgerufen am: 11.12.2017
[4] https://www.bnl.gov/newsroom/news.php?a=24251 Abgerufen am: 12.11.2017
[5] https://www.suedostschweiz.ch/politik/im-thurgau-muss-uber-sieben-tonnen-honig-
vernichtet-werden Abgerufen am: 11.12.20217
[6] https://www.thermofisher.com/fr/fr/home/life-science/cell-
culture/transfection/selection.html Abgerufen am: 19.09.2017
[7] http://www.the-odin.com/crispr-bacterial-guide/ Abgerufen am: 02.10.2017
[8] https://www.nzz.ch/wissenschaft/medizin/gentechnik-crispr-bringt-krebskranken-neue-
hoffnung-ld.103183
7.3 Abbildungen
Abbildung 2 https://www.bnl.gov/newsroom/news.php?a=24251
Abbildung 3 Bildschirmaufnahmen aus einem Film des McGovern Institute for Brain
Research am MIT. https://www.youtube.com/watch?v=2pp17E4E-O8
Abbildung 4 https://de.wikipedia.org/wiki/Genetischer_Code#/media/File:
Aminoacids_table.svg
Die restlichen Abbildungen sind eigene Abbildungen des Autors.
20Maturitätsarbeit Miles Burri
8 Glossar
Fachbegriff Erklärung
30S-Untereinheit Teil des Ribosoms.
Aminoglykosid-Antibiotikum Bakterizid wirkender Stoff, hemmt die Proteinbiosynthese.
Animpfen Auftragen von Zellen auf ein Nährmedium.
Ausplattierrechen L-Förmiges Instrument zum Verteilen von Zellen auf Nährböden.
bakterizid Bakterien abtötend.
Bändermodell Darstellungsvariante für Proteine bei der Sekundärstrukturen
sichtbar sind.
Basenpaar Komplementär liegende Basen im Doppelstrang der DNA, zusam-
mengehalten durch Wasserstoffbrücken.
DNA Desoxyribonukleinsäure, Träger der Erbinformation. Form einer
Doppelhelix.
Eppendorf-Röhrchen Plastikgefäss mit Deckel und Volumen im Mikroliterbereich.
preFeuerbrand Vor allem bei Kernobstbäumen vorkommende Pflanzenkrankheit,
ausgelöst durch das Bakterium Erwinia amylovora.
homologe Rekombination Neuanordung von genetischem Material basierend auf einer ähn-
lichen Nukleotidsequenz.
Impföse Instrument zum Auftragen von Zellen auf Nährmedien.
Inkubation Ausbrühten von Organismen unter guten Bedingungen.
Inokulation Auftragen von Zellen auf ein Nährmedium.
mRNA Von engl. „Messenger-RNA“, beteiligt an der Proteinbiosynthese
im Ribosom.
Nukleinbase Variabler Teil eines Nukleotids, ihre Reihenfolge enthält die ei-
gentliche Information des Erbguts.
Nukleotid Bausteine von Nukleinsäuren, z. B. von DNA und RNA.
Plasmamembran Jede Zelle umgebende Schicht, bildet die Grenze zur Aussenwelt.
Prokaryoten Zelluläre Organismen ohne Zellkern, Vertreter sind Bakterien und
Archaeen.
Proteinbiosynthese Neubildung von Proteinen in Zellen.
Ribosom Ort der Proteinbiosynthese in der Zelle, indem die DNA-Sequenz
in Aminosäuresequenz übersetzt wird.
RNA Einzelsträngige Ribonukleinsäure.
rpsL-Gen Gen, welches die Informationen für die Synthese des S12-Pro-
teins enthält.
S12-Protein Teil der 30S-Untereinheit, wichtig für die Genauigkeit der Transla-
tion.
Stichkultur Nährmedium in länglichem Röhrchen, das mittels Einstechen ei-
nes Drahtes mit Bakterien angeimpft wurde, eignet sich für Lage-
rung von Bakterien.
Transformation (Mikrobiologie) Übertragung von DNA in eine kompetente Zelle
Translation Synthese von Proteinen am Ribosom von lebenden Organismen.
T-Zelle Teilgruppe der weissen Blutzellen, beteiligt an der Immunab-
wehr.
Wildtyp Organismus, dessen Erbgut durch Evolution zustande kam und
das in der Natur überwiegt.
zellfreie Genexpression Proteinbiosynthese ausserhalb einer lebenden Zelle.
21Maturitätsarbeit Miles Burri
9 Anhang
Rohdaten Versuche:
CRISPR Versuch Mutatuionsversuch Kontrollgruppe (Kontrollgruppe,
(Durchmesser in mm) (Durchmesser in mm) (Durchmesser in mm) Fortsetzung)
1,1 1,6 2,00 2,20
1,0 1,5 2,50 2,20
0,9 1,5 2,30 2,20
0,8 1,5 2,10 2,50
0,6 1,6 2,20 2,60
0,6 1,3 1,80 2,20
0,9 1,7 2,00 2,10
0,7 1,8 2,50 2,30
0,4 1,7 2,10 2,40
2,00 2,30
0,7 1,5
2,00 1,70
1,0 1,9
2,40 2,40
0,6 1,4
2,20 2,00
0,5 1,9
2,10 2,00
1,0 1,5
2,30 2,60
0,9 1,6
2,30 2,20
1,1 Ø = 1.6
2,30 2,30
1,1 2,10 2,40
1,0 2,20 2,20
1,8 2,00 2,40
1,2 2,40 2,50
2,0 2,30 2,20
1,3 2,20 2,50
1,5 2,40 2,10
1,4 2,40 2,40
1,0 2,50 2,50
1,2 2,20 2,40
1,1 2,20 2,40
1,0 2,10 2,30
0,9 2,20 2,30
1,0 2,10 2,50
1,4 2,30 2,60
1,4 2,40 2,50
1,3 1,90 2,20
1,5 2,30 2,30
1,1 2,00 2,30
0,9 2,40 2.6
2,50 Ø = 2,27
0,8
2,40
0,5
2,40
0,8
Ø = 1.03
22Sie können auch lesen

















































