3.2.16 Gnitzen (Ceratopogonidae) als Überträger von sich ausbreitenden Infektionserkrankungen bei Tieren
←
→
Transkription von Seiteninhalten
Wenn Ihr Browser die Seite nicht korrekt rendert, bitte, lesen Sie den Inhalt der Seite unten
3. Aktuelle und potenzielle Gefahren für die Gesundheit
3.2.16 Gnitzen (Ceratopogonidae) als Überträger von sich
ausbreitenden Infektionserkrankungen bei Tieren
Helge Kampen & Doreen Werner
Gnitzen (Ceratopogonidae) als Überträger von sich ausbreitenden Infektionserkrankungen bei Tieren:
Hämatophage Arten der Ceratopogoniden-Gattung Culicoides sind Vektoren zahlreicher Krankheitserreger,
insbesondere von Vertebraten-pathogenen Viren. Die Erreger der zwei wichtigsten Infektionen, Blauzungen-
krankheit und Afrikanische Pferdesterbe, werden, global betrachtet, im Wesentlichen von Culicoides imicola
übertragen. Diese Gnitzen-Art afroasiatischer Herkunft hat sich erst in den 1980er Jahren im Mittelmeerraum
etabliert; aperiodische Ausbrüche der Blauzungenkrankheit folgten dort seit 1998. Sowohl der Vektor als auch
die Krankheit scheinen sich nach Norden auszubreiten, vermutlich als Konsequenz der Klimaerwärmung.
Culicoides imicola war jedoch nicht am Blauzungenausbruch 2006 in Mitteleuropa beteiligt, bei dem ein-
heimische Gnitzen für die Virusübertragung verantwortlich waren. Dieselben Culicoides-Arten waren wahr-
scheinlich in die Epidemiologie der Schmallenberg-Krankheit eingebunden, die durch ein 2011 neu entdecktes
Virus verursacht wird. Informationen über die Biologie und Ökologie der einheimischen Gnitzen-Arten sind
spärlich, so dass eine Abschätzung ihrer Vektorrollen oder sogar Vorschläge von Maßnahmen zum Seuchen-
management, die mit der Vektoraktivität und biologie interferieren, problematisch sind. Während eine inten-
sivere Erforschung der europäischen Gnitzen-Fauna gerade erst begonnen hat, ist zu befürchten, dass die
Klimaerwärmung zu einer weiteren Ausbreitung von C. imicola Richtung Nordeuropa führt und weitere Ce-
ratopogoniden-übertragene Krankheitserreger, wie etwa das Afrikanische Pferdesterbe-Virus, mit sich bringt.
Biting midges (Ceratopogonidae) as vectors of spreading infectious diseases of animals: Haematopha-
gous species of the ceratopogonid genus Culicoides are vectors of numerous disease agents, predominantly
of viruses pathogenic to vertebrate animals. From a global point of view, the agents of the two most im-
portant infections, bluetongue disease and African horse sickness, are mainly transmitted by Culicoides
imicola. This ceratopogonid species of Afroasian origin has established in the Mediterranean only in the
1980s, followed by aperiodic bluetongue epidemics since 1998. Both the vector and the disease appear
to be spreading further northwards into Europe, possibly as a consequence of climate change. Culicoides
imicola, however, has not been implicated in the 2006 outbreak of bluetongue disease in central Europe,
where indigenous biting midges were responsible for virus transmission. The same culicoid species were
probably involved in the epidemiology of Schmallenberg disease which is caused by a virus newly dis-
covered in 2011. Information on the biology and ecology of the indigenous biting midge species is scar-
ce, so it is difficult to assess their various vector roles or to even suggest outbreak management measures
interfering with vector activity and biology. While more in-depth research on the European biting midge
fauna has just started, it is to be feared that climate change will allow C. imicola to spread towards nor-
thern Europe and bring along further ceratopogonid-borne pathogens such as African horse sickness virus.
A uf das weit verbreitete Vorkommen der Gnitzen,
ihr häufiges Auftreten als Lästlinge von Mensch
und Tier sowie ihr besonderes Potenzial als Überträ
möglich oder zumindest schwierig ist und dass somit
über die Verbreitung, Biologie und Ökologie der ein
zelnen Arten nur wenig bekannt ist. Weiterhin fehlen
ger von Krankheitserregern auch für den europäischen entscheidende Informationen, die für eine Bemessung
Bereich wurde schon vor Jahrzehnten hingewiesen. Im der Effekte abiotischer und biotischer Faktoren auf die
Jahr 2003 verdeutlichte eine vom Umweltbundesamt Entwicklung des Vektors und eine auf den speziellen
herausgegebene Studie, dass für Deutschland derzeit Fall bezogene Analyse allgemeiner infektionsepidemio
generell keine gesicherte Bewertung der Ausbreitungs logischer Vorgänge erforderlich sind.
risiken vieler Infektionskrankheiten möglich ist, deren Ungeachtet der frühen Warnungen und der nach
Erreger durch Vektoren übertragen werden (Maier et drücklich konstatierten Forschungsdefizite wurden bis
al. 2003). Dies betrifft insbesondere Pathogene, für die heute in Deutschland nur vereinzelte Studien zu dieser
Arten aus der Familie Ceratopogonidae (Gnitzen) als Insektenfamilie durchgeführt. So kam der Ausbruch
Überträger in Frage kommen, und resultiert aus den der Blauzungenkrankheit (engl. bluetongue disease,
erheblichen Forschungsdefiziten, die für diese und BTD) in Mitteleuropa im Jahr 2006 für viele Wissen
andere hämatophage Arthropoden bestehen. In vielen schaftler und Behörden völlig überraschend und hat die
Fällen ist deren Vektorpotenzial ebenso wenig unter Brisanz der Situation deutlich gemacht. Während das
sucht wie ihre Wechselwirkungen mit Pathogenen und Auftreten der BTD zuvor in erster Linie im Bereich der
Wirbeltierwirten. Besonders problematisch ist aber, Verbreitungsgrenzen des primären Vektors, Culicoides
dass selbst eine korrekte Artidentifizierung oft nicht imicola, als möglich erachtet worden war, offenbarte
3.2.16 Kampen & Werner
die Analyse des Schadgeschehens 2006, dass in Europa Stunde sogar bis zu 40.000 Gnitzen Versuchspersonen
offenbar noch weitere Ceratopogoniden-Arten Vektor an (Mordue & Mordue 2003). Stiche von C. impunc-
kompetenz für das Blauzungenvirus (engl. bluetongue tatus und anderen Culicoides-Arten können bei Weide
virus, BTV) besitzen (s. Kap. 3.1.18 - Melhhorn - in tieren schwerste allergische Reaktionen (sog. »sweet
diesem Band). Die sich bis ins Jahr 2009 fortsetzende itch« bei Pferden) auslösen.
Epidemie und ihre gravierenden Folgen für die Vieh Weitaus wichtiger sind Ceratopogoniden als Über
wirtschaft wurden u.a. mit klimatischen Veränderungen träger von Krankheitserregern. In der Gattung Culicoi
in Verbindung gebracht. des findet man Vektoren von zahlreichen Viren sowie
vielen Protozoen- und Filarienarten, die allerdings
Allgemeines zu Gnitzen fast ausschließlich veterinärmedizinische Bedeutung
(Fam. Ceratopogonidae) haben (Werner & Kampen 2010). Die einzige bekann
Gnitzen sind 0,5–3 mm kleine Dipteren, die sich über termaßen mit Ceratopogoniden assoziierte Krankheit
vier Larvenstadien und ein Puppenstadium zur flugfä des Menschen ist das in Lateinamerika vorkommende
higen Imago entwickeln. Bei vielen paläarktischen Arten Oropouche-Fieber, eine dem Dengue-Fieber ähnliche
dauert die Entwicklung von der Eiablage bis zum adulten Viruserkrankung, deren Erreger vorwiegend von Culi-
Tier unter durchschnittlichen sommerlichen Tempera coides paraensis übertragen wird. Der Erreger des in
turen (20–25 °C) etwa 4–6 Wochen. Die Überwinterung Afrika vorkommenden Rifttal-Fiebers, einer norma
findet gewöhnlich im vierten Larvenstadium statt, so lerweise durch Stechmücken (Fam. Culicidae) über
dass sich die Präimaginalentwicklung dann auch über tragenen Virusinfektion, wurde aus Ceratopogoniden
mehr als sieben Monate erstrecken kann. Die Lebens isoliert, ohne dass diesen jedoch eine Vektorfunktion
dauer der Weibchen beträgt vermutlich in den meisten nachgewiesen werden konnte. Das Rifttal-Fiebervirus
Fällen maximal einen Monat (Szadziewski et al. 1997). stammte ursprünglich aus Ostafrika, breitete sich in
Das natürliche Vorkommen der Ceratopogoniden den letzten Jahren aber bis nach Nordafrika (Ägypten)
ist fast ausnahmslos an Feuchtgebiete gebunden, da die sowie auf die arabische Halbinsel aus. Eine Einschlep
Präimaginalstadien aquatisch oder semiaquatisch le pung ins südliche Europa wird befürchtet (Martin et al.
ben. Vielen Arten reicht feuchtes Substrat, wie z.B. sich 2008). Gelegentlich übertragen Ceratopogoniden Fila
zersetzendes organisches Material, als Bruthabitat. Von rien (verschiedene Mansonella-Spezies) auf den Men
den weltweit über 5.500 Gnitzen-Arten gehören etwa schen, doch gelten diese i.A. als apathogen und führen
1.400 zur Gattung Culicoides. Die Weibchen von rund nur in seltenen Fällen zu einer Symptomatik.
96% der Arten dieser Gattung sind obligat hämatophag Unter den tierpathogenen Erregern, die von Cera
und nehmen von Vögeln und Säugern, einschließlich topogoniden übertragen werden, fallen v.a. die Viren
des Menschen, Blut auf (Meiswinkel et al. 1994). Da ins Gewicht. Hierunter sind Erreger ggf. tödlich verlau
Gnitzen häufig in großer Zahl auftreten, sind sie ge fender Erkrankungen von Wiederkäuern und Equiden
fürchtete Lästlinge. Darüber hinaus gelten etwa 50 Cu- zu finden (z.B. bovines ephemeres Fieber, epizootisches
licoides-Arten als Überträger von Krankheitserregern. hämorrhagisches Fieber, equine Enzephalose und Aka
Diese haben in der Veterinärmedizin eine weitaus hö bane-Fieber). In Deutschland kam es zuletzt überra
here Relevanz als in der Humanmedizin und verursa schend zu mehrjährigen Epidemien der Blauzungen-
chen alljährlich enorme volkswirtschaftliche Schäden. und der Schmallenberg-Krankheit bei Wiederkäuern.
Für Deutschland sind nach Havelka & Aguilar (1999) Während die Blauzungenkrankheit aufgrund der cha
insgesamt 332 Ceratopogoniden-Arten beschrieben, rakteristischen Symptomatik schon mehr als hundert
von denen 56 zur Gattung Culicoides gehören. Jahre bekannt ist, handelt es sich bei der Schmallen
berg-Krankheit um eine erst 2011 aufgetauchte, zuvor
Medizinische Bedeutung unbekannte Entität mit einem völlig neuen Erreger.
der Ceratopogonidae Sie blieb bislang ein rein europäisches Phänomen. Die
Aufgrund von saisonalen und regionalen Massenvor schnelle Ausbreitung des Schmallenberg-Virus in fast
kommen spielen Gnitzen allein schon durch ihre Stech alle Länder Europas muss als Folge hoher Infektiosi
aktivität eine große Rolle als Plageerreger. So führt z.B. tät bei gleichzeitig fehlender Immunität der Wirtstiere
Culicoides impunctatus insbesondere im schottischen sowie der grenzüberschreitenden Verbringung von
Hochland zu ganz erheblichen Belästigungen von virämischen Wiederkäuern, die als Infektionsquelle für
Mensch und Tier. Bei entsprechenden Untersuchungen anderweitig vorkommende Gnitzen dienten, gesehen
in solchen Gebieten wurden pro Nacht z.T. mehr als werden (Conraths et al. 2013). Ein klimatischer Ein
6.000 Gnitzen-Weibchen pro Falle gefangen (Mands fluss auf die Epidemiologie der Schmallenberg-Krank
et al. 2004). Einer anderen Studie zufolge flogen pro heit kann derzeit nicht abgeleitet werden.
3. Aktuelle und potenzielle Gefahren für die Gesundheit
Besonders hervorgehoben werden sollen im Fol und des C. pulicaris-Komplexes als Vektoren in Ver
genden die BTD und die Afrikanische Pferdesterbe dacht (Kampen & Werner 2010).
(engl. African horse sickness, AHS), die aufgrund Trotz der mangelhaften Kenntnisse zur Biologie
ihrer oft dramatischen tiergesundheitlichen und öko und Ökologie der einheimischen Gnitzen-Arten be
nomischen Folgen von der Weltorganisation für Tier stand in Mitteleuropa die Hoffnung, dass die Epide
gesundheit OIE (Office International des Épizooties) mie mit dem saisonalen Verschwinden der Gnitzen im
als meldepflichtig gelistet werden. Beide Krankheiten Winter 2006/2007 von selbst erlöschen würde. Im Juni
haben in der alten Welt in der jüngeren Vergangenheit und Juli 2007 traten in Westdeutschland, Belgien und
signifikante Ausbreitungstendenzen gezeigt, welche Holland allerdings wieder die ersten Fälle der BTD im
vermutlich mit der Ausbreitung ihres wichtigsten natür neuen Jahr auf, so dass die Hoffnung sich nicht bestä
lichen Vektors, C. imicola, in Zusammenhang stehen. tigt sah. Auch in den Jahren 2008 und 2009 wurden
weiterhin zahlreiche BTD-Fälle verzeichnet, die auf
Ausbreitung der Blauzungen- denselben BTV-Serotypen zurückgingen. Die Ursache
krankheit und der Pferdepest für die Persistenz des Virus im Ökosystem ist höchst
Die BTD wurde Ende des 19. Jahrhunderts erstmals wahrscheinlich bei den Gnitzen zu suchen. Ob aller
bei Schafen in Südafrika beschrieben, wo Arten des C. dings in Mitteleuropa Gnitzen-Arten existieren, die die
imicola-Komplexes bis heute Überträger des BTV sind. Winterperiode aktiv im Adultstadium oder durch fort
In Europa trat die Seuche bereits in den 1920er Jahren währende Reproduktion überstehen können, ist unklar.
regelmäßig auf Zypern auf; darüber hinaus wurden in »Wärmeinseln«, wie dungbrütende Arten (z.B. C. de-
Europa jedoch bis Ende des 20. Jahrhunderts nur zwei wulfi) sie evtl. in Tierställen finden, oder milde Winter,
Ausbrüche registriert: ein größerer in den späten 1950er die eine durchgehende Aktivität sonst diapausierender
Jahren auf der iberischen Halbinsel und ein kleinerer Arten bedingen könnten, werden als Ursachen erwogen
Ende der 1970er Jahre auf einigen griechischen Inseln (Wilson et al. 2008).
(Wilson & Mellor 2008). Erst in Verbindung mit der Das Afrikanische Pferdesterbe-Virus (African
Etablierung von C. imicola in Südeuropa kam es ab 1998 horse sickness virus, AHSV) hat seinen Ursprung ver
dort zu zahlreichen aperiodisch auftretenden BTD-Epi mutlich ebenfalls im südlichen Afrika, wo Zebras das
demien. Heute ist fast der gesamte europäische Mittel natürliche Reservoir darstellen. Die Krankheit wurde
meerraum als Endemiegebiet anzusehen, aus dem das aber erst nach der Einführung von Pferden durch Eu
Virus auch in den Wintermonaten nicht verschwindet. ropäer im 17. und 18. Jahrhundert erkannt, da Pferde
Die in Südeuropa vorkommenden Serotypen des BTV im Gegensatz zu Zebras meist schwer an der Infektion
sind vermutlich über importierte infizierte Wiederkäuer erkranken. Man geht davon aus, dass das AHSV heute
aus Nordafrika und dem Nahen Osten nach Südeuropa fast im gesamten tropischen und subtropischen Afrika
gelangt. Wiederkäuer weisen eine begrenzte virämische südlich der Sahara endemisch ist, möglicherweise auch
Phase auf (maximal etwa 100 Tage). Nur in dieser Zeit in Teilen der arabischen Halbinsel, wie serologische
können sich blutsaugende Gnitzen infizieren. Eine Untersuchungen nahe legen. Periodische, z.T. erheb
Epidemie erlischt deshalb dort, wo die Überträger im liche Ausbreitungen des Virus über den afrikanischen
Winter länger inaktiv sind als die virämische Phase an Kontinent hinaus wurden in den letzten Jahrzehnten
dauert. Während das in Jahren mit gewöhnlichen Tem registriert. So kam es zwischen 1959 und 1961 zu
peraturverläufen z.B. in Nordspanien der Fall ist, sind einem Ausbruch, der sich über den Mittleren Osten bis
die Gnitzen in Südspanien ganzjährig aktiv und halten nach Pakistan und Indien hinzog und Hunderttausende
den Infektionszyklus aufrecht. von Pferden das Leben kostete. Derselbe Virus-Sero
Da das prinzipielle Risiko einer Ausbreitung der typ sorgte nur wenige Jahre später für einen weiteren
BTD schon seit längerem auch in manchen mittel- und Ausbruch in Nordwestafrika und Südspanien (Mellor
nordeuropäischen Staaten gesehen worden war, beob 1993). In Ermangelung eines dauerhaften Wirbeltierre
achtete man die Ausbreitung von C. imicola genaues servoirs sowie von langfristig verfügbaren effizienten
tens. Ungeachtet dessen trat die Seuche im Sommer Vektoren konnte sich das Virus aber nie länger als zwei
2006 völlig unerwartet in Holland, Belgien, Frankreich, Jahre in solchen epidemischen Gebieten halten.
Luxemburg und Deutschland auf. Sofort initiierte Stu Verursacht durch einen bis dahin nur im südlichen
dien wiesen allerdings C. imicola in keinem der betrof Afrika beobachteten Serotyp des AHSV kam es 1987
fenen Gebiete nach. Nach bisherigen Erkenntnissen zu einem Ausbruch der Pferdepest in Zentralspanien,
stehen stattdessen Arten des C. obsoletus-Komplexes der auf den Import virämischer Zebras aus Namibia
inkl. C. dewulfi, die von manchen Autoren nicht als in einen Safaripark bei Madrid zurückgeführt werden
dem C. obsoletus-Komplex zugehörig betrachtet wird, konnte (Lubroth 1988). Entgegen der Erwartung der
3.2.16 Kampen & Werner
Fachleute bedeutete der Winter nicht das Ende des gender Temperatur zu. Eine infizierte Gnitze ist also
Seuchengeschehens. Das Virus konnte sich offenbar in bei höheren Temperaturen für einen Blutwirt schneller
wärmeren Regionen Südspaniens halten und breitete infektiös. Des Weiteren findet ein Transmissionszyklus
sich von dort in den nächsten drei Jahren immer wieder umso intensiver statt, je größer die Gnitzen- und Wir
nach Zentralspanien und Portugal aus. Mittlerweile hat beltierwirt-Populationen sind. Umgekehrt nimmt die
sich bestätigt, dass das AHSV durchaus in südlichen Länge der EIP bei sinkenden Temperaturen ab bis sie,
Regionen der iberischen Halbinsel überwintern kann, in Abhängigkeit von der Erregerspezies, zu einem Still
da sich auch der Überträger des Virus, C. imicola, dort stand kommt. Gleichzeitig verlängert sich die Lebens
etabliert hat und das milde Klima eine ganzjährige Ak dauer des Vektors. Diese grundlegenden Erwägungen
tivität des Vektors erlaubt. Die fortgeschrittene Verbrei treffen prinzipiell auf jeden Vektor und die von ihm
tung von C. imicola im Mittelmeerraum lässt zukünftig übertragenen Pathogene zu (Mellor 2000).
Ausbrüche der Pferdepest auch in anderen europäischen Der weltweit wichtigste Überträger des BTV und
Mittelmeerländern befürchten. des AHSV, C. imicola, ist eine wämeliebende Ceratopo
Während des Ausbruchs der Pferdepest in Spani goniden-Art, die in Afrika und Südostasien beheimatet
en und Portugal 1987–1991 konnte das AHSV auch in ist. In Europa wurde sie erstmals 1982 in Südspanien
gepoolten Freilandfängen von C. obsoletus und C. puli- nachgewiesen, war aber vermutlich schon lange vorher
caris nachgewiesen werden (Mellor 1994). Es ist hier eingeschleppt worden (Wilson & Mellor 2008). Bei
also eine ähnliche Situation gegeben wie bei dem BTV: ihrer Einschleppung fand sie also offenbar ein für eine
C. imicola ist in Südeuropa vermutlich der wichtigste, Ansiedlung geeignetes Klima vor. Im Jahr 2000 waren
aber möglicherweise nicht der alleinige Überträger der weite Bereiche des südeuropäischen Mittelmeerraumes
Viren, während in nördlicheren Gebieten, wo C. imi- besiedelt. Darüber hinaus ist eine deutliche Ausbrei
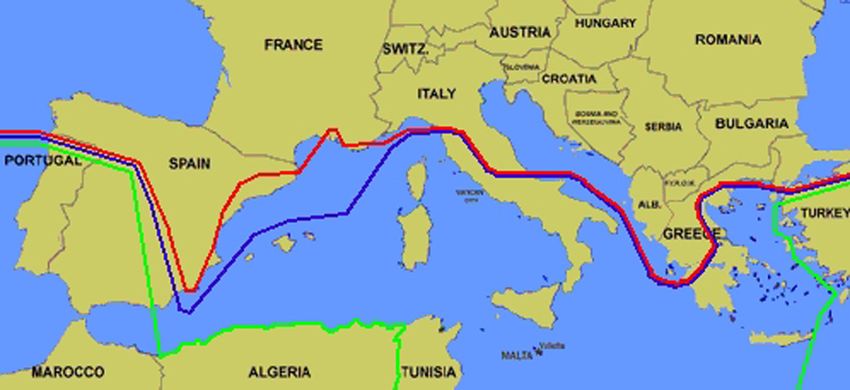
cola (noch) nicht vorkommt, andere Ceratopogoniden- tungstendenz nach Norden erkennbar (Abb. 3.2.16-1),
Arten mit Vektorpotenzial definitiv (BTV) oder mögli die möglicherweise als Konsequenz von klimatischen
cherweise (AHSV) heimisch sind. Veränderungen zu sehen ist (Purse et al. 2005). Wäh
Einfluss des Klimas auf das rend die Imagines in den meisten europäischen Verbrei
Vorkommen und die Verbreitung tungszonen im Winter verschwinden, sind sie in eini
von Ceratopogonidae und gen Regionen Spaniens und Portugals das ganze Jahr
auf deren Vektorpotenzial hindurch aktiv. Geht man bis Mitte des 21. Jahrhundert
Der Einfluss des Klimas auf das geografische und sai von einer Temperaturerhöhung um 1,5 bis 3 °C aus,
sonale Vorkommen und die Entwicklung von Gnitzen wobei ein Anstieg von 1 °C etwa 90 km geographische
sowie auf deren Rolle als Überträger von Krankheits Breite und 150 m Höhe entspricht, so könnte C. imicola
erregern, somit auf die Verbreitung von Gnitzen-asso bald in Mitteleuropa auftauchen (Mellor et al. 2000).
ziierten Krankheiten, ist vielfältig (Wilson & Mellor In warmen Sommermonaten dringt C. imicola bereits
2008). Eine prinzipielle direkte und indirekte Abhän bis nach Südfrankreich vor (Baldet et al. 2005), und
gigkeit von Temperatur, Niederschlag und relativer in der Südschweiz wurde 2003 ein Exemplar gefangen
Luftfeuchtigkeit gilt prinzipiell für alle hämatophagen (Cagienard et al. 2006).
Arthropoden und die von ihnen übertragenen Patho Infektionsversuche mit BTV und AHSV wurden
gene. Wie jedes andere Lebewesen, das äußeren Klima mit verschiedenen Gnitzen-Arten bei 24–25 °C durch
bedingungen ausgesetzt ist, haben sich Gnitzen im Lau geführt (Mellor 2000). Bei einem Teil der infizierten
fe der Evolution an die spezifischen Bedingungen ihres Gnitzen war eine Virus-Übertragung nach ein bis zwei
natürlichen Verbreitungsgebietes optimal angepasst. Wochen nachweisbar. Die Lebensdauer der Gnitzen al
Ausreichende Luftfeuchtigkeit vorausgesetzt, sind sie lerdings war bei solchen konstant hohen Temperaturen
zu bestimmten Jahreszeiten und bestimmten Tempera stark reduziert. Bei niedrigeren Temperaturen nahm
turen aktiv. Werden diese Temperaturen früher im Jahr die Rate infektiöser Gnitzen ab, aber ihre Lebensdauer
oder früher am Tag erreicht, so setzt die Aktivität ent zu. Die Übertragungswahrscheinlichkeit ist daher eine
sprechend früher ein, solange nicht andere Faktoren, Funktion dieser beiden gegensätzlichen Trends. Bei
wie z.B. Tageslänge, interferieren. Mit der Temperatur wechselnden Temperaturen war für das Übertragungs
steigen die Saugfrequenz, die Entwicklungsgeschwin potenzial ausschließlich die kumulative Zeit entschei
digkeit und die Populationsdichte, allerdings nimmt dend, die bei für die Virogenese ausreichenden Tempe
auch die Lebensdauer des Einzelindividuums ab. Auch raturen verbracht wurde.
die Entwicklungsgeschwindigkeit des Erregers in der Auch bei eigentlichen Nicht-Vektoren, wie die in
Gnitze (extrinsische Inkubationsperiode, EIP) nimmt Europa weit verbreitete Art C. nubeculosus, können
– bis zu einem gewissen Maximum – i.d.R. mit stei hohe Temperaturen während der Larvalentwicklung
3. Aktuelle und potenzielle Gefahren für die Gesundheit
dazu führen, dass eine Virus-Replikation im adulten pulicaris eher eine stabile hohe relative Luftfeuchtig
Gnitzen-Weibchen und eine Übertragung stattfinden. keit der entscheidende Faktor ist. Auf Korsika wurde
Hierbei wird der übliche Infektionsweg der Gnitze für beide Arten eine bimodale Saisondynamik mit Ma
über die Mitteldarm-Epithelzellen übergangen, und das xima im Frühjahr und im Herbst beobachtet, was für
Virus gelangt aus dem Darm der Gnitze gleich in die einen ungünstigen Einfluss hoher Temperaturen und
Leibeshöhle (»leaky gut«-Phänomen), wo es die inne niedriger relativer Luftfeuchtigkeiten spricht (Mellor
ren Organe infizieren und zur Replikation nutzen kann 2000). Aufgrund der verschiedenen temperaturabhän
(Wittmann 2000). Hohe Temperaturen während der gigen Prozesse im Lebenszyklus von Ceratopogo
Larvalentwicklung bedingen kleinere adulte Gnitzen niden sind – gemessen an den Fangzahlen – höchste
mit einer dünneren Basallamina, was vermutlich die Populationsdichten generell unmittelbar vor und nach
Ursache für die Durchlässigkeit der Darmepithelbarrie den heißen Sommermonaten festzustellen. Hier sind
re ist. Vergleichbare Effekte mit kleinwüchsigen Weib sowohl Temperatur-, als auch Luftfeuchtigkeitswerte
chen und durchlässigem Darmepithel treten auch bei förderlich für die Aktivität der meisten Gnitzen-Arten.
dauerhaft schlechter larvaler Ernährungssituation und Entsprechend waren die in Mitteleuropa aufgetretenen
sehr hohen Larvendichten auf (Tabachnik 1996), wie BTD-Ausbrüche stets durch einen bimodalen Verlauf
sie allgemein bei hohen Populationsdichten aufgrund gekennzeichnet: einem ersten Maximum im Juni/Juli
von idealen Klima- und Entwicklungsbedingungen zu folgte ein zweites Mitte Oktober. Als Ursache für den
beobachten sind. schnellen Rückgang der BTV-Fälle im November und
BTV stellt bei Temperaturen unter 10 °C, AHSV Dezember wird die jahreszeitlich bedingte Temperatur
bei Temperaturen unter 15 °C die Replikation im Vek abnahme und die daraus resultierenden Konsequenzen
tor komplett ein. Gnitzen-Aktivität ist dagegen auch bei für Vektoren und Virogenese betrachtet.
niedrigeren Temperaturen festzustellen, so z.B. bei C. Das Klima ist im Allgemeinen auch mit der Hö
imicola noch bei ca. 7 °C. Steigt die Umgebungstempe henlage korreliert. Eine Studie aus der Schweiz zeigt,
ratur an, so setzt auch die Vermehrung des Virus in der dass Gnitzen dort in allen Höhenlagen und in allen
Gnitze wieder ein (Mellor 2000). Klimazonen vorkommen, wobei aber das Artenspek
Die Anpassung der Gnitzen-Arten an bestimmte trum schwankt und die dominierenden Arten in den
Temperaturbedingungen gilt ebenso für ihre Anpas verschiedenen Klimazonen andere sind (Kaufmann et
sung an die Luftfeuchtigkeit, wenngleich eine Min al. 2009). Die höhenabhängige Artenverteilung und
destluftfeuchtigkeit für das Vorkommen aller Arten die unterschiedlichen Vektorkompetenzen der Gnitzen,
eine notwendige Voraussetzung ist. So scheint z.B. das ebenso wie die Temperaturabhängigkeit der Patho
Entwicklungsoptimum der bis nach Nordeuropa weit genentwicklung in den Überträgern, würden hier wie
verbreiteten Art C. obsoletus durch die Temperaturen anderswo die Epidemiologie einer Gnitzen-assoziier
bestimmt zu werden, während für das Auftreten von C. ten Krankheit bestimmen. In Übereinstimmung mit
Abb. 3.2.16-1: Mutmaßliche nördliche Verbreitungsgrenzen von C. imicola in Europa
(grün: 1999, blau: 2002, rot: 2005) (modifiziert nach Mellor 2004).
3.2.16 Kampen & Werner
prinzipiellen Erwägungen zur Klimaabhängigkeit der Während sich C. imicola nur dort etablieren kann, wo
Gnitzen und des BTV zeigt die Analyse des BTD-Ge der monatliche Durchschnitt der Tageshöchsttempera
schehens von 2006, dass die Anzahl der Ausbrüche mit turen in den sieben kältesten Monaten des Jahres 12,5
zunehmender Höhenlage sank. Bereits ab 400 m traten °C nicht unterschreitet (Mellor & Boorman 1995),
nur noch Einzelfälle auf, in Höhen über 650 m wurden sind auch den von Ceratopogoniden übertragenen Vi
gar keine Fälle mehr verzeichnet. Die absolute Mehr ren Temperaturgrenzen gesetzt. Angesichts unserer
zahl der Ausbruchsherde lag im Tiefland in Höhen bis Wissenslücken bleibt es offen, ob bei entsprechenden
zu 50 m (EFSA 2007). Temperaturbedingungen zukünftig auch mit einer
Das Klima kann gegebenenfalls kleinräumig deut Übertragung des AHSV durch die einheimischen Gnit
lich von der allgemeinen Großwetterlage abweichen. zen-Arten zu rechnen ist oder ob das Virus erst im Ge
So können z.B. Stallungen auch bei widrigen äußeren folge von C. imicola bei uns auftreten wird.
Bedingungen geeignete Klimainseln für Gnitzen sein
und Einfluss auf ein Schadgeschehen nehmen. Bei Literatur
Untersuchungen in Frankreich traten einige Gnitzen- BALDET T., MATHIEU B., DELECOLLE J. C.,
Arten, darunter vermutete BTV-Vektoren, nach tem GERBIER G. & F. ROGER (2005): Emergence de la
fièvre catarrhale ovine dans le Bassin méditerranéen
peraturbedingtem Rückgang der Fänge im Freiland in
et surveillance entomologique en France. Rev. Élev.
Stallungen noch in relativ hoher Zahl auf (EFSA 2007), Méd. Vét. Pays Trop. 58, 125-132.
und in einem Versuchsstall in Belgien konnten im Win CAGIENARD A., GRIOT C., MELLOR P. S., DENI
ter 2006/2007 fast jede Nacht Gnitzen gefangen wer SON E. & K. D. STÄRK (2006): Bluetongue vector
den (Losson et al. 2007). species of Culicoides in Switzerland. Med. Vet. En
tomol. 20, 239-247.
Schlussbetrachtung CONRATHS F, PETERS B. & M. BEER (2013):
Klimaveränderungen beeinflussen zweifellos die Ver Schmallenberg virus, a novel orthobunyavirus infec
breitung Vektor-assoziierter Infektionskrankheiten. So tion in ruminants in Europe: Potential global impact
würden Temperaturerhöhungen vielfach nicht nur die and preventive measures. New Zeal. Vet J. 61, 63-
67.
Entwicklung bekannter tropischer und subtropischer EFSA (European Food Safety Authority) (2007): Epi
Überträger von Krankheitserregern über ihre bisherigen demiological analysis of the 2006 bluetongue virus
Verbreitungsgrenzen hinaus fördern, sondern könnten serotype 8 epidemic in north-western Europe. Parma,
auch die Entwicklung von Krankheitserregern in ein Italien. http://www.efsa.europa.eu/de/efsajournal/
heimischen, unbekannterweise vektorkompetenten Ar doc/34r.pdf
thropoden ermöglichen. HAVELKA P. & M. AGUILAR (1999): Ceratopogoni
Inwieweit klimatische Faktoren zum Ausbruch dae. Studia Dipterol., Suppl. 2, 80-82.
der Blauzungenkrankheit in Mitteleuropa beitrugen KAMPEN H. & D. WERNER (2010): Three years of
bluetongue disease in central Europe with special
oder dessen Ausmaß beeinflussten, ist nicht geklärt. reference to Germany: What lessons can be learned?
Während sich die BTD-Forschung auf die Biologie Wien. Klin. Wochenschr. 122 (Suppl. 3), 31-39.
und klimatisch bedingte Ausbreitung von C. imicola KAUFMANN C., SCHAFFNER F. & A. MATHIS
konzentrierte, wurde in neuen Studien deutlich, dass (2009): Monitoring von Gnitzen (Culicoides spec.),
auch einheimische, bisher nicht als Überträger in Er den potentiellen Vektoren des Blauzungenkrankheits
scheinung getretene Culicoides-Arten ohne präadap virus, in den 12 Klimaregionen der Schweiz. Sch
tive Phase in der Lage sind, das BTV zu übertragen. weiz. Arch. Tierheilk. 151, 205-213.
Der Ausbruch der BTD zeigt daher eindrucksvoll, wie LOSSON B., MIGNON B., PATERNOSTRE J., MAD
DER M., DE DEKEN R., DE DEKEN G., DEB
wichtig die Erhebung von gesicherten Daten zur Biolo
LAUWE I., FASSOTTE C., CORS R., DEFRANCE
gie und Ökologie einheimischer hämatophager Arthro T., DELÉCOLLE J.C., BALDET T., HAUBRUGE
poden sind. Erst diese gestatten abgesicherte Modellie E., FREDERIC F., BORTELS J. & G. SIMONON
rungen und Prognosen zu eventuellen klimabedingten (2007): Biting midges overwintering in Belgium.
Effekten. Die nordwärts gerichtete Verschiebung der Vet. Rec. 160, 451-452.
für die Artverbreitung kritischen Temperaturuntergren LUBROTH J. (1988): African horse sickness and the
ze und die damit verknüpften biologischen Effekte sind epizootic in Spain 1987. Equine Pract. 10, 26-33.
anhand von Daten aus pflanzenphänologischen Mess MAIER W. A., GRUNEWALD J., HABEDANK B.,
netzen geostatistisch valide fassbar und können epide HARTELT K., KAMPEN H., KIMMIG P., NAUCKE
T., OEHME R., VOLLMER A., SCHÖLER A. & C.
miologisch modelliert werden. SCHMITT (2003): Mögliche Auswirkungen von
Unterdessen scheint die klimatisch bedingte Aus Klimaveränderungen auf die Ausbreitung von primär
breitung von C. imicola nach Norden fortzuschreiten. humanmedizinisch relevanten Krankheitserregern
3. Aktuelle und potenzielle Gefahren für die Gesundheit
über tierische Vektoren sowie auf die wichtigen (Diptera: Ceratopogonidae) an Weidetieren in Nord
Humanparasiten in Deutschland. Climate Change deutschland. Ergebnisse aus dem Freiland und dem
05/03, Forschungsbericht 20061218/11 im Auftrag Laboratorium. Dissertation Tierärztliche Hochschule
des Umweltbundesamtes (UBA-FB 000454), erstellt Hannover.
im Rahmen des Umweltforschungsplanes des Bun PURSE B. V., MELLOR P. S., ROGERS D. J., SAM
desministeriums für Umwelt, Naturschutz und Reak UEL A. R., MERTENS P. P. & M. BAYLIS (2005):
torsicherheit. Climate change and the recent emergence of blue
MANDS V., KLINE D. L. & A. BLACKWELL (2004): tongue in Europe. Nat. Rev. Microbiol. 3, 171-181.
Culicoides midge trap enhancement with animal SZADZIEWSKI R., KRZYWIŃSKI J. & W. GIŁKA
odour baits in Scotland. Med. Vet. Entomol. 18, 336- (1997): Diptera Ceratopogonidae, biting midges.
342. In: Nilsson A.N. (Hrsg.), Aquatic Insects of North
MARTIN V., CHEVALIER V., CECCATO P., ANY Europe – A Taxonomic Handbook, Vol. 2. Apollo
AMBA A., DE SIMONE L., LUBROTH J., DE LA Books, Stenstrup, Denmark. 243-263.
ROCQUE S. & J. DOMENECH (2008): The impact TABACHNIK W. J. (1996): Culicoides variipennis and
of climate change on the epidemiology and control bluetongue virus epidemiology in the United States.
of Rift Valley fever. Rev. Sci. Tech. Off. Int. Epiz. Annu. Rev. Entomol. 41, 23-43.
27, 413-426. WERNER D. & H. KAMPEN (2010): Gnitzen (Dip
MEISWINKEL R., NEVILL E. M. & G. J. VENTER tera, Ceratopogonidae) und ihre medizinische Be
(1994): Vectors: Culicoides spp. Aust. J. Zool. 38, deutung. Denisia 30, 245-260.
68-89. WILSON A., DARPEL K. & P. S. MELLOR P. S.
MELLOR P. S. (1993): African horse sickness: trans (2008): Where does bluetongue virus sleep in the
mission and epidemiology. Vet. Res. 24, 199-212. winter? PLoS Biol. 6, e210.
MELLOR P. S. (1994): Epizootiology and vectors of WILSON A. & P. S. MELLOR P. S. (2008): Blue
African horse sickness. Comp. Immun. Microbiol. tongue in Europe: vectors, epidemiology and climate
Infect. Dis. 17, 287-296. change. Parasitol. Res. 103 (Suppl. 1), S69-S77.
MELLOR P. S. (2004): Infection of vectors and blue WITTMANN E. J. (2000): Temperature and the trans
tongue epidemiology in Europe. Vet. Ital. 40, 167- mission of arboviruses by Culicoides. Dissertation
174. Universität Bristol, UK.
MELLOR P. S. (2000): Replication of arboviruses in
insect vectors. J. Comp. Path. 123, 231-247.
MELLOR P. S. & J. BOORMAN (1995): The transmis
sion and geographical spread of African horse sick
ness and bluetongue virus. Ann. Trop. Med. Parasi
Kontakt:
tol. 89, 1-15 PD Dr. Helge Kampen
MELLOR P. S., BOORMAN J. & M. BAYLIS (2000): Friedrich-Loeffler-Institut
Culicoides biting midges: their role as arbovirus vec Bundesforschungsinstitut für Tiergesundheit, Insel Riems
tors. Annu. Rev. Entomol. 45, 307-340. helge.kampen@fli.bund.de
MORDUE J. & W. MORDUE (2003): Biting midge Dr. Doreen Werner
chemical ecology. Biologist 50, 159-162. Institut für Landnutzungssysteme, Leibniz-Zentrum für
OLBRICH S. (1987): Untersuchungen zur Biolo Agrarlandschaftsforschung e.V., Müncheberg
gie von Gnitzen der Gattung Culicoides Latreille Doreen.Werner@zalf.de
Kampen, H. & D. Werner (2014): Gnitzen (Ceratopogonidae) als Überträger von sich ausbreitenden Infektionser-
krankungen bei Tieren. In: Lozán, J. L., Grassl, H., Karbe, L. & G. Jendritzky (Hrsg.). Warnsignal Klima: Gefahren für
Pflanzen, Tiere und Menschen. 2. Auflage. Elektron. Veröffent. (Kap. 3.2.16) - www.klima-warnsignale.uni-hamburg.
de.
Sie können auch lesen

















































