Beitrag der beiden Kalttransduktionskanäle TRPM8 und TRPA1 zur Kodierung kalter Temperaturen in mono- und polymodalen C-Fasern der Maus - Opus4
←
→
Transkription von Seiteninhalten
Wenn Ihr Browser die Seite nicht korrekt rendert, bitte, lesen Sie den Inhalt der Seite unten
Beitrag der beiden Kalttransduktionskanäle
TRPM8 und TRPA1 zur Kodierung kalter Temperaturen
in mono- und polymodalen C-Fasern der Maus
Anästhesiologische Klinik
der Medizinischen Fakultät
der Friedrich-Alexander-Universität
Erlangen-Nürnberg
zur
Erlangung des Doktorgrades
Dr. med.
vorgelegt von
Philipp Gruschwitz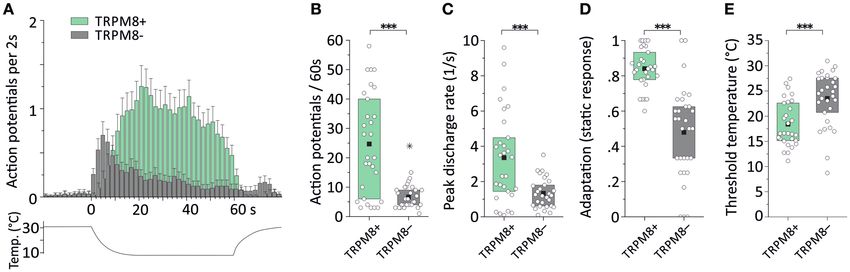
Als Dissertation genehmigt von der Medizinischen Fakultät der Friedrich-Alexander-Universität Erlangen-Nürnberg Vorsitzender des Promotionsorgans: Prof. Dr. med. Markus F. Neurath Gutachter/in: Prof. Dr. Katharina Zimmermann Prof. Dr. Alexey Ponomarenko Tag der mündlichen Prüfung: 01. Dezember 2020
meinen Eltern in Dankbarkeit gewidmet
Inhaltsverzeichnis
1A Zusammenfassung ................................................................................. 1
1B Abstract .................................................................................................. 3
2 Einleitung ............................................................................................... 5
3 Material und Methoden........................................................................... 8
3.1 Versuchstiere ......................................................................................... 8
3.2 Haut-Saphenus-Nerv-Präparation .......................................................... 9
3.3 Einzelfasermessung ............................................................................. 10
3.3.1 Kurzfassung des Versuchsaufbaus ...................................................... 11
3.3.2 Datenakquisition ................................................................................... 13
3.4 Synthetic Interstitial Fluid (SIF) ............................................................ 16
3.5 Statistische Analyse / Datenanalyse .................................................... 17
3.6 Temperatur-Gradienten-Messgerät ...................................................... 18
4. Physiologischer Hintergrund ................................................................ 19
4.1 Transient-Rezeptor-Potential Ionenkanäle (TRP-Ionenkanäle) ............ 19
4.1.1 TRPA1.................................................................................................. 21
4.1.2 TRPM8 ................................................................................................. 22
4.2 Nervus saphenus ................................................................................. 24
4.3 Nervenfasern ........................................................................................ 24
4.3.1 Kategorie C-Fasern .............................................................................. 24
4.5 Somatosensorik ................................................................................... 26
4.5.1 Nozizeption und Schmerz .................................................................... 26
4.5.3 Weiterleitung ins Zentrale Nervensystem (ZNS) .................................. 28
4.5.4 Kaltsensibilität, Temperaturwahrnehmung und Verhaltenstests ........... 29
5 Ergebnisse ........................................................................................... 34
5.1 Expression von TRPM8-Kanälen steigert die Kaltantwort von
Kaltnozizeptoren .................................................................................. 35
5.2 Das Fehlen von TRPM8-Kanälen führt zu deutlichen Kodierungsdefiziten
in Kaltnozizeptoren ............................................................................... 39
5.3 Ohne TRPM8- und TRPA1-Kanäle gibt es nur noch wenige polymodale
kaltsensitive C-Fasern .......................................................................... 39
5.4 Rein kaltsensitive C-Fasern (CC) sind maßgeblich auf TRPM8-Kanäle
zur Kalttransduktion angewiesen ......................................................... 42
6 Diskussion ............................................................................................ 46
7 Tabellenverzeichnis.............................................................................. 54
8 Literaturverzeichnis .............................................................................. 55
9 Abkürzungsverzeichnis ........................................................................ 67
10 Verzeichnis der eigenen Veröffentlichungen ........................................ 68
11 Anhang ................................................................................................. 69
12 Danksagung ......................................................................................... 70
13 Lebenslauf............................................................................................ 71
1A Zusammenfassung
Hintergrund:
In den letzten Jahren publizierte Arbeiten beschreiben TRPM8- und TRPA1-
Rezeptoren als wichtig für die Detektion von Temperaturen im kühlen und kalten
Bereich sowie für die Übermittlung von gewebeschädigender Kälte und damit
Nozizeption. Während der TRPM8-Kanal bereits in der Breite der Wissenschaft
als der essenzielle Transduktionskanal für Kaltsignale anerkannt ist, wurde dies
für den TRPA1-Kanal bis vor kurzem noch kontrovers diskutiert. Aufgrund
dessen, dass in der Vergangenheit eine Reihe widersprüchlicher Ergebnisse
publiziert wurden, die nicht reproduziert werden konnten. Einerseits wird der
Kanal in Einzeluntersuchungen als entscheidend für die Kaltwahrnehmung
genannt. Andererseits konnten aber in mehreren Studien verschiedener
Forschungsgruppen, die Einzel- und Doppel-Knockout (DKO)-Mäuse
einbezogen, keine signifikanten Defizite in der Kaltwahrnehmung im Vergleich zu
einzelnen TRPM8-Knockouts festgestellt werden. Durch einen
Verhaltensversuch unserer Forschungsgruppe zeigte sich allerdings unter
Zuhilfenahme eines neu konzipierten Temperaturgradienten-Tests, dass die
Doppel-Knockout-Tiere in diesem Verhaltenstest ein deutlich vermindertes
Kaltvermeidungsverhalten aufweisen als Tiere, denen nur einer der beiden
funktionsfähigen Kanäle fehlt.
Ziele:
Ziel dieser Forschungsarbeit ist es daher, dieses Defizit in der Kaltwahrnehmung
auf neurophysiologischer Ebene mittels Messung an kutanen C-Fasern am
Nervus-saphenus-Hautpräparat dieser Mäuse zu überprüfen.
Methoden:
Dafür wurden die weitergeleiteten Aktionspotentiale auf direkte Kaltreize im
rezeptiven Feld verschiedener C-Faser-Typen von Doppel-Knockout-Mäusen
(TRPA1 und TRPM8) mit denen von reinen TRPM8-Knockout-Mäusen sowie
einer Kontrollgruppe von C57BL/6J-Mäusen mit Wildtyp-Status sowohl
quantitativ als auch qualitativ verglichen.
[1]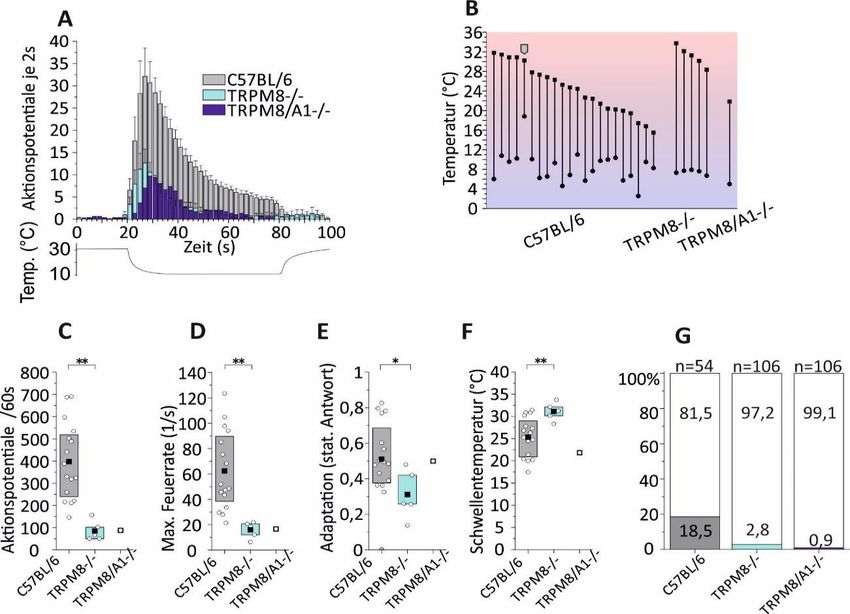
Ergebnisse/Beobachtungen:
Dabei zeigte sich, dass eine C-Faser gleich welchen Typs (mono- oder
polymodal) prinzipiell durch das Vorhandensein eines funktionsfähigen TRPM8-
Kanals befähigt wird, größere Kaltantworten mit höherer
Aktionspotenzialfrequenz bei dynamischer Temperaturänderung und höherer
Momentanfrequenz bei konstanter Temperatur zu generieren. Darüber hinaus
können in TRPM8-Knockout-Mäusen deutlich weniger kaltsensitive C-Fasern
nachgewiesen werden als in Wildtyp-Mäusen. Die Doppel-Knockout-Mäuse
zeigten zwar eine deutlich ausgeprägtere Reduktion der Kaltfaseranzahl
gegenüber den TRPM8-Knockouts, sodass monomodale C-Faser mittels
Einzelfasermessung nicht mehr detektiert werden konnten. Allerdings zeigte sich,
dass die verbleibenden Antworten der Nozizeptoren in entscheidenden
Charakteristika wie Aktivierungsschwelle sowie maximaler und mittlerer
Aktionspotentialfrequenz auf Kaltreize nur marginal zu den Fasern der TRPM8-
negativen C57BL/6J-Mäuse verändert sind.
Schlussfolgerungen:
Zusammenfassend ergibt sich das eindeutige Bild, dass der TRPM8-Kanal im
Vergleich zum TRPA1-Kanal einen größeren Einfluss sowohl auf die Quantität
kaltsensitiver monomodaler und polymodaler C-Fasern als auch auf die
Antwortgröße der Kaltantwort besitzt. Diese neurophysiologischen Ergebnisse
scheinen im Widerspruch zu dem relativ milden Defizit der TRPM8-defizienten
Mäuse zu stehen, welches sich im Temperaturgradienten-Test auf den
Temperaturbereich um 27 - 32 °C beschränkt und sich erst im Doppel-Knockout
auf dem gesamten Temperaturmessbereich zwischen 5 - 30 °C
beziehungsweise 15 - 40 °C offenbart. Deshalb ist davon auszugehen, dass eine
kooperative Funktion der Kanäle bei der Signaltransduktion vorliegt, bei der das
Fehlen des einen Kanals durch den jeweils anderen Kanal kompensiert werden
kann. Diese Kompensation ist beispielsweise durch verstärkte Expression,
Sensibilisierung oder andere Anpassungsvorgänge vorstellbar. Hieraus ergibt
sich weiterer Forschungsbedarf, um diese Kompensations- und
Kooperationsmechanismen zu untersuchen, die auch bei der Anpassung an
wechselnde Umgebungstemperaturen von Bedeutung sein könnten.
[2]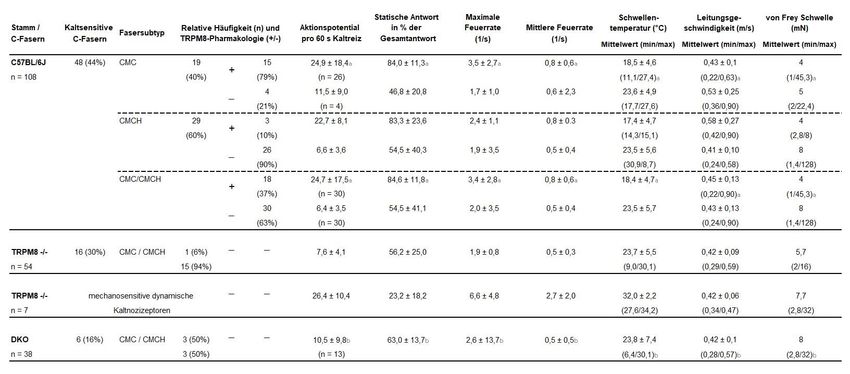
Ergebnisse der vorliegenden Arbeit wurden veröffentlicht in:
Winter, Gruschwitz et al. 2017; Cold Temperature Encoding by Cutaneous
TRPA1 and TRPM8-Carrying Fibers in the Mouse; Frontiers in molecular
neuroscience 10, S. 209. DOI:10.3389/fnmol.2017.00209.
1B Abstract
Background:
Previous research provided sufficient evidence that both transient receptor
potential channel family members TRPM8 and TRPA1 play a crucial role in the
detection of cool as well as cold and noxious cold environmental temperatures.
Whilst there is sufficient evidence for the TRPM8 channel being essential in the
transduction of cool temperatures, the involvement of TRPA1 in the transduction
of noxious cold was under debate for a long time. Some researchers identified
overt deficits in TRPA1-deficient mice in specific behavioral assays, but on the
other hand, double-knockout mice showed no differences compared to single
TRPM8-deficient mice when being confronted with cool or even noxious cold
temperatures.
Aims:
The study was designed to phenotype the precise deficits in cold avoidance and
nocifensive behaviour of TRPM8-/-, and double-knockout mice for further
correlation with the electrophysiological characteristics of the primary afferent
cold responses in monomodal and polymodal cold-sensitive C-fibers. We
compared the frequency and the characteristics of the cold responses of C-fiber
nociceptors in C57BL/6J wild-type mice to nociceptors of TRPM8-deficient and
DKO mice.
Methods:
The presented study used a novel thermal gradient assay to allow more precise
phenotyping of cold avoidance deficits, in combination with ex vivo single fiber
recordings of saphenous nerve skin preparations.
[3]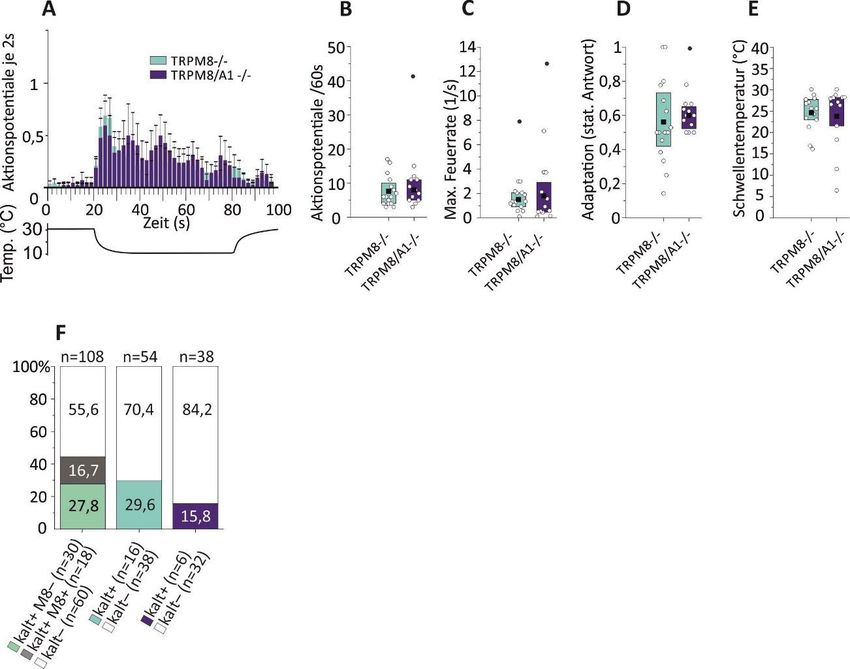
Results:
We found that the number of cold-sensitive C-fibers depends on the expression
of functional TRPM8. Most cold-sensitive monomodal C-fibers were identified in
C57BL/6J mice followed by TRPM8-/- and lastly DKO specimen with fewest C-
fibers. Additionally, the expression of TRPM8 allows nociceptors responding to
cold to generate higher firing frequencies and larger response with a reinforced
static component. Nevertheless, the pattern of cold responses of the remaining
cold nociceptors in DKO mice did not differ significantly from those of TRPM8-
negative C-fibers or the C57BL/6J strain.
Conclusions:
In summary, the TRPM8 channel plays an essential role in the transduction of
cold temperatures in primary afferent C-fibers. A lack of this channel attenuates
the physiological characteristics of the cold response in both monomodal
thermoreceptors and nociceptors. The quantity of functional cold-sensitive
thermoreceptors decreases even further when the TRPA1 channel is also
removed (DKO). Nevertheless, DKO mice show a disproportionately larger lack
in cold avoidance behavior than TRPM8-/- and specimen of the wildtype strain.
These results suggest the assumption TRPA1 and TRPM8 possessing a
cooperative function to cover the transduction of the entire temperature range
from cool, to cold and noxious cold.
Results have been published:
Winter, Gruschwitz et al. 2017; Cold Temperature Encoding by Cutaneous
TRPA1 and TRPM8-Carrying Fibers in the Mouse; Frontiers in molecular
neuroscience 10, S. 209. DOI:10.3389/fnmol.2017.00209.
[4]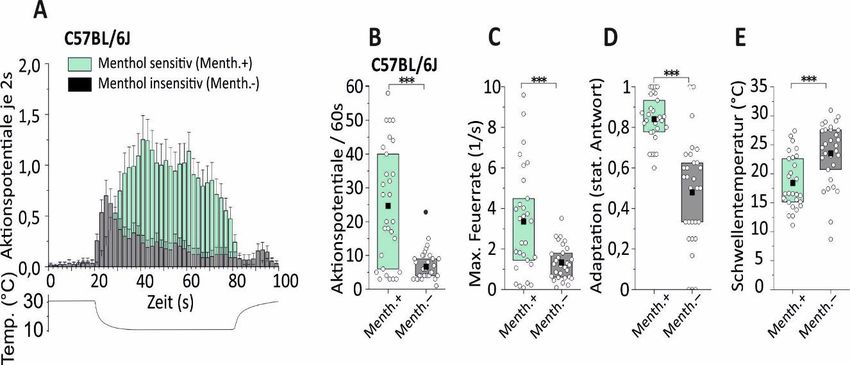
2 Einleitung
Wie für gleichwarme Tiere üblich, zeigen auch Mäuse eine Präferenz für
Temperaturen, die es ihnen ermöglichen, ihre Körpertemperatur energieeffizient
aufrechtzuerhalten. Dafür ist es allerdings notwendig, die
Umgebungstemperaturen präzise wahrzunehmen, um zu kühle oder zu warme,
aber auch schädlich kalte oder heiße Temperaturen möglichst genau und rasch
detektieren zu können, um diese zu meiden. Dieses Vermeidungsverhalten
wurde in verschiedenen Untersuchungen mit unterschiedlichen
verhaltensphysiologischen Messmethoden für die C57BL/6J-Maus aufgezeigt.
Ebenso wurde solches Vermeidungsverhalten quantifiziert, um den Beitrag des
TRPM8-Kanals auf die Kalttransduktion in den Nervenendigungen der Tiere zu
prüfen. So zeigten z.B. Versuchsaufbauten wie der 2-Platten-Wahltest, bei dem
die Maus zwischen zwei Platten unterschiedlicher Temperatur wählen kann
(Bautista et al. 2007; Dhaka et al. 2007; Colburn et al. 2007), der
Thermogradienten-Test, bei dem die Maus in einem linearen
Temperaturgradienten zwischen 15 °C und 50 °C ihre Präferenztemperatur frei
wählt (Dhaka et al. 2007), oder auch das Beobachten von Abwehrreaktionen
(zum Beispiel Pfotenlecken) auf provozierte Verdunstungskälte mittels Aceton
(Bautista et al. 2007) übereinstimmend, dass TRPM8-Knockout-Mäuse ein
deutlich herabgesetztes Kaltvermeidungsverhalten zeigen als die
entsprechenden Kontrollgruppen. Im Gegensatz zur Bedeutung des TRPM8-
Rezeptors wurde der Beitrag des TRPA1-Kanals zur Kalttransduktion und
Kaltwahrnehmung lange kontrovers diskutiert. TRPA1-Knockout-Tiere zeigten im
oben genannten 2-Platten-Temperaturwahltest in ihrem Verhalten keinen
signifikanten Unterschied zu TRPM8-Einzelknockout-Tieren (Knowlton et al.
2010). Auch bei Vergleichsuntersuchungen von Wildtyp und TRPA1-Knockouts
mittels Aceton-induzierter Verdunstungskälte bzw. Kontakt zu kalten Oberflächen
konnten keine Verhaltensunterschiede festgestellt werden (Bautista et al. 2006).
In der darüber hinaus angewendeten Quantifizierung von c-Fos war die Anzahl
der positiven Nervenzellen nach Kaltinduktion gleich derer von Wildtypmäusen
(Knowlton et al. 2010). Hierbei wurden sensorische Nervenendigungen der Haut
eines der beiden Hinterläufe mittels Kälte (wiederholte Exposition einer
Temperatur von 0 °C über 30 min) und der Chemikalien Menthol bzw. Icilin
gereizt und anschließend die induzierte Expressionsänderung des Gens c-Fos in
[5]Nervenzellen des Rückenmark-Hinterhorns bestimmt. Dies ist allgemein
anerkannt als Indikator für gesteigerte neurale Aktivität (Vetter et al. 2013).
Andere Forschungsgruppen konnten hingegen Kaltsensitivität von TRPA1
nachweisen (Klionsky et al. 2007; Sawada et al. 2007; Story et al. 2003). Im
Einzelnen gelang dies mit Versuchsverfahren wie Calcium-Imaging, bei dem der
intrazelluläre Calcium-Anstieg, der bei Stimulation der Zelle entsteht, über
Chelat-Bildung beziehungsweise Bindung an genetisch modifizierte
fluoreszierende Proteine und deren resultierende bzw. hierdurch modulierte
Fluoreszenz optisch gemessen wird (Grienberger und Konnerth 2012). Ebenso
wurden auch sogenannte Patch-Clamp-Untersuchungen erfolgreich verwendet,
wobei mittels einer Glaselektrode der durch Öffnung oder Verschluss von
Kanälen verursachte elektrische Strom bei ansonsten konstant gehaltenem
elektrischen Membranpotential (clamp, englisch für befestigen) in einem
umschriebenen Areal der Zellmembran (patch, englisch für Flicken) von
unversehrten Zellen oder in Membranfragmenten bestimmt werden (Neher und
Sakmann 1992). Story zeigt bereits 2003 eine Aktivierung von CHO-Zellen
(chinese hamster ovary, Ovarzellen des chinesischen Hamsters), welche murine
TRPA1-Kanäle exprimierten, bei Temperaturen < 20 °C (17,5 ± 3,5 °C) mittels
Calcium-Imaging. Zu diesen Ergebnissen passend, konnten bei diesen Zellen
kaltinduzierte Ströme im Ganzzell-Patch-Clamp mit ähnlicher
Schwellentemperatur abgeleitet werden. Auch die Untersuchung von TRPA1-
exprimierenden XO-Zellen (Xenopus oocyte; Krallenfrosch-Oozyten) im Patch-
Clamp bestätigt diese Ergebnisse (Story et al. 2003). Zusätzlich wurden
kultivierte DRG-Neurone von C57BL/6-Mäusen in Zellkultur untersucht. Dabei
zeigte sich eine kaltsensitive Zellpopulation, die nicht Menthol-sensitiv war und
im Vergleich zu den Methol-sensitiven Zellen eine erniedrige
Schwellentemperatur von ~15 °C (gegenüber ~24 °C bei TRPM8-positiven DRG-
Neuronen) aufwies. Aufgrund der vergleichbaren Aktivierungscharakteristika zu
den TRPA1-tragenden CHO- beziehungsweise XO-Zellen ist davon auszugehen,
dass diese Zellen TRPA1-positiv waren. Eine Bestätigung mittels
Ligandentestung wurde allerdings nicht durchgeführt (Story et al. 2003). Sawada
nutzte kultivierte DRG-Neuronen (dorsal root ganglion; Spinalganglion) von
C57BL/6-Wildtyp-Mäusen sowie humane TRPA1-Känale-exprimierende HEK-
Zellen (humane embryonale Nierenzellen) und zeigte sowohl mittels Calcium-
[6]Imaging eine Kaltaktivierung beider Zelltypen (DRG bei < ~15 °C; HEK bei
< ~18 °C) als auch eine Kaltsensitivität des TRPA1-Kanals (< ~16 °C) im
Einzelkanal-Patch-Clamp (Sawada et al. 2007). Im gleichen Jahr bestätigte eine
weitere Forschungsgruppe die Kaltsensitivität des TRPA1-Kanals im noxischen
Temperaturbereich. Untersucht wurden CHO-Zellen, welche entweder humane
oder murine TRPA1-Kanäle exprimierten, beziehungsweise TRPA1-nativ waren.
Maximale Calciumaufnahme (induzierte Lumineszenz durch radioaktives 45Ca2+)
konnte bei Reizung der TRPA1-positiven Zellen durch noxisch kalte
Badtemperaturen (3 – 7 °C) gemessen werden, während die TRPA1-negativen
Vergleichszellen in diesem Temperaturberiech (Gesamttemperaturreiz
3,5 – 25 °C) die geringste Aktivierung zeigten (Klionsky et al. 2007).
Auch funktionelle cerebrale Magnetresonanztomografien von Wildtyp- und
TRPA1-Knockout-Mäusen bei denen das BOLD-Signal (Blood Oxygenation
Level Dependent, englisch für Blutoxygenierungsgrad-abhängig) nach erfolgten
Kaltreizen am Hinterlauf als Surrogatparameter für neuronale Aktivität bestimmt
wurde, konnten zum einen eine geringe Gesamtaktivierung bei TRPA1 -/- als auch
eine Verschiebung der Aktivierung vom sensorischen Cortex in Areale wie
Thalamus, Striatum und spinoretikulärem Trakt als Zeichen für eine reduzierte
Kaltwahrnehmung nachweisen (Vetter et al. 2012).
Zusätzlich wurde vor kurzem gezeigt, dass der TRPA1-Kanal auf zellulärer
Ebene eine Rolle bei der Detektion von Hitzereizen spielt (Moparthi et al. 2014).
Ein Umstand, der nahe legt, dass hier komplexe Verschaltungsvorgänge bzw.
Synergismen/Antagonismen anderer Kanäle auf nervaler Ebene einen Einfluss
haben könnten (Vriens et al. 2014).
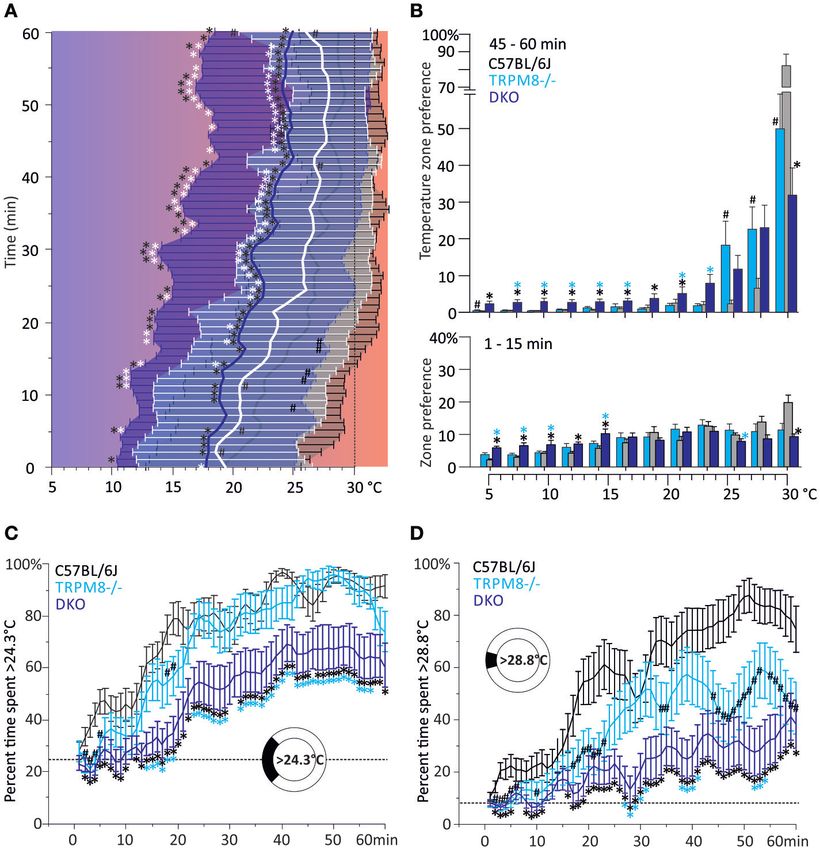
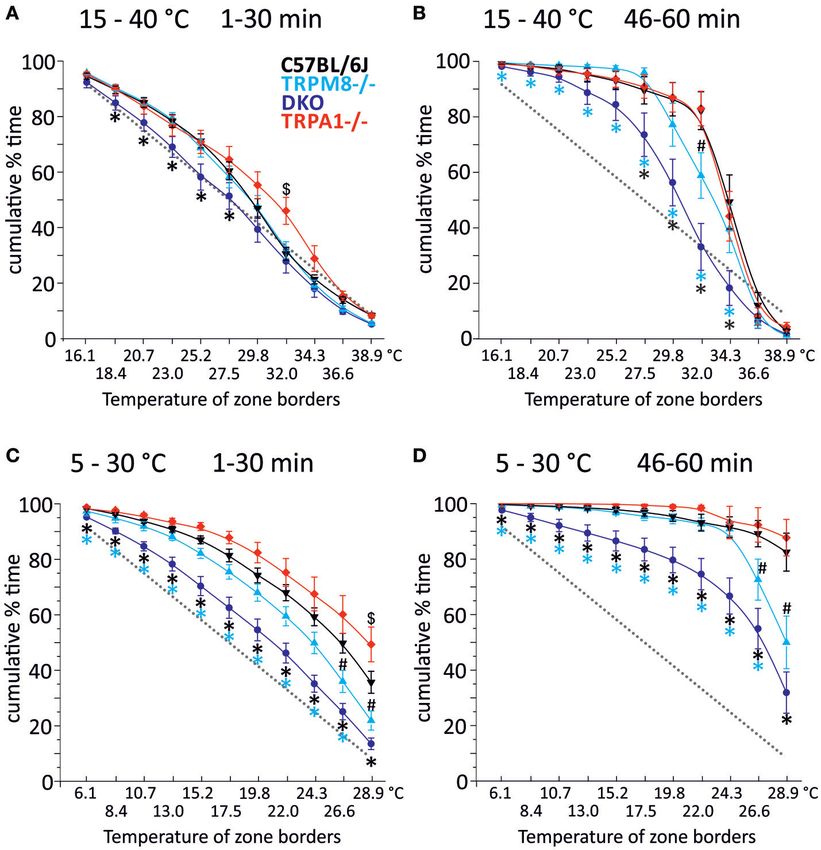
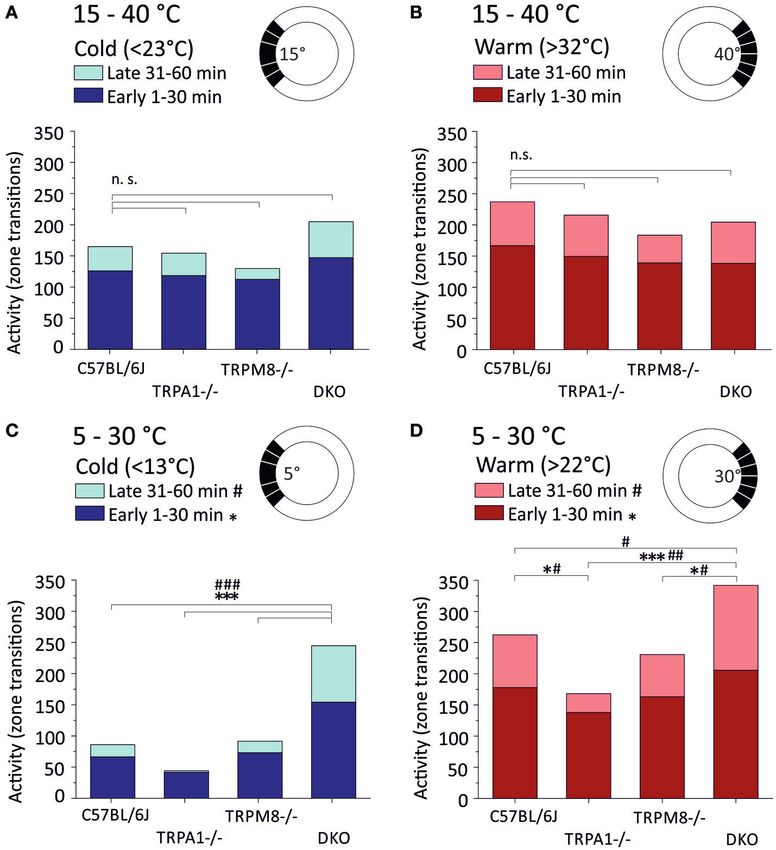
Der von unserer Forschungsgruppe entwickelte runde Temperaturgradienten-
Assay zeigt ein signifikant reduziertes Kaltvermeidungsverhalten der
Doppelknockout-Mäuse sowohl im Vergleich zu den TRPM8-Kockouts, welche
bekannter Weise selbst ein reduziertes Kaltvermeidungsverhalten zeigen, als
auch im Vergleich zu den TRPA1-Knockouts und Wildtyp-Mäusen (Winter et al.
2017; Touska et al. 2016).
Daraus ergibt sich die Frage, inwieweit sich dieser Unterschied im
Kältevermeidungsverhalten zwischen den TRPM8-defizienten und DKO-Mäusen
auf Veränderung der Kaltsensitivität der primären Afferenzen zurückführen lässt.
[7]Um diese Frage zu beantworten, wurden Einzelfasermessungen an C-Fasern der
TRPM8- sowie Doppelknockout-Mäuse durchgeführt und die spezifischen
Charakteristika dieser Fasern mit denen von TRPM8-positiven und -negativen
Fasern, die bereits in einer früheren Studie in Wildtyp-Mäusen gemessen
wurden, verglichen. Dabei zeigten sich Unterschiede sowohl in der Anzahl
kaltsensitiver C-Fasern als auch im dynamischen Antwortverhalten und der
Adaptation auf Kältereize im zeitlichen Verlauf.
3 Material und Methoden
Im Nachfolgenden werden die verwendeten Materialien und
Untersuchungsmethoden genauer beleuchtet. Neben den Kontroll- und
Knockout-Tierstämmen und deren Herkunft soll dabei vor allem auf die
elektrophysiologischen Einzelfaserableitungen am Hautnervenpräparat
eingegangen werden. Der in der Publikation angewandte Temperaturgradienten-
Assay sowie die statistischen Verfahren zur Testung werden ebenfalls
beschrieben.
3.1 Versuchstiere
Die für die Arbeit verwendeten Mäuse besitzen den genetischen Hintergrund des
in den Lebenswissenschaften am häufigsten verwendeten C57BL/6J-
Inzuchtstammes und werden hier als Wildtyptiere bezeichnet (Zucht Charles
River, Sulzfeld, Deutschland). Für die Einzelfasermessung wurden als
Kontrollgruppe Mäuse des Wildtypstammes mit Tieren aus der Linie der TRPA1-
und TRPM8-defizienten Tiere, hier als Doppelknockout (DKO) bezeichnet,
verglichen. Die TRPM8-Knockoutmäuse entstammen der Zucht von Ardem
Patapoutian (The Scripps Research Institute, La Jolla, CA) (Dhaka et al. 2007)
und wurden über sieben bis acht Generationen auf den C57BL/6J-Hintergrund
gekreuzt. Die TRPA1-Knockoutmäuse wurden über 13 Generationen auf
C57BL/6J rückgekreuzt und entstammen dem Labor von David Corey (Harvard
Medical School, Boston, MA) (Kwan et al. 2006). Durch Verpaarung der beiden
Linien entstanden im Institut für Physiologie und Pathophysiologie der FAU
Erlangen-Nürnberg TRPM8/TRPA1-DKO-Mäuse.
Für die Hautnervenpräparation und anschließende Einzelfasermessung wurden
insgesamt 22 erwachsene DKO-Mäuse (17 Männchen und 4 Weibchen, 60 bis
85 Tage alt) verwendet (Abbildungen 5 und 6). Weitere Daten von
[8]Einzelfaserableitungen an Wildtyptieren und TRPM8-/- aus früheren
Publikationen wurden in diesem Zuge neu analysiert (Vetter et al. 2013). Hierzu
wurden Messungen von 53 C57BL/6J-Tieren (37 Männchen, 16 Weibchen)
sowie 16 männlichen TRPM8-/- und 14 männlichen TRPM8+/+ untersucht
(Abbildungen 4 – 6). Die TRPM8-Mäuse waren zu diesem Zeitpunkt für vier bis
fünf Generation auf einen C57BL/6J-Hintergrund rückgekreuzt.
Darüberhinausgehende Daten, die in Abbildung 5 Verwendung fanden, stammen
von 5 männlichen TRPM8-Knockout- und 3 erwachsenen männlichen C57BL/6J-
Mäusen. Zusätzlich sind Ableitungen von aus C57BL/6J-Mäusen stammenden
CC-Fasern integriert, die ebenfalls bereits publiziert worden sind (Toro et al.
2015).
Alle verwendeten Mäuse wurden im Tierstall des Instituts für Physiologie und
Pathophysiologie der FAU Erlangen-Nürnberg gehalten und gezüchtet. Den
Tieren wurde zusätzlich zur Hautpräparat-Entnahme eine Schwanzspitzen-
Biopsie entnommen und ebenfalls im Institut für Physiologie und
Pathophysiologie entsprechend des früher publizierten Vorgehens (Vetter et al.
2012; Vetter et al. 2013) auf den korrekten Genotyp-Status getestet.
Sämtliche Verhaltensversuche an den Versuchstieren wurden von der
Tierschutzkommission der Universität Erlangen-Nürnberg befürwortet und von
der örtlichen Bezirksregierung (Regierung von Mittelfranken) genehmigt.
3.2 Haut-Saphenus-Nerv-Präparation
Die verwendeten Mäuse wurden vor der Präparation nach aktuell geltenden
Tierschutzregeln unter Verwendung von Kohlenstoffdioxid betäubt und getötet.
Anschließend führe eine provozierte Luxation im Atlantoaxialgelenk zum
sicheren Tod der verwendeten Versuchstiere. Das Fell an den Beinen wird
abrasiert, da dieses bei der Präparation hinderlich wäre und dessen hydrophobe
Eigenschaften im weiteren Versuchsaufbau zu Problemen führen würde. Nach
Fixierung der Plantarfläche des Fußes mittels doppelseitigen Klebebands auf
einen Plexiglaskeil wird mit einer kleinen Schere die Haut der Maus direkt
proximal des Kniegelenks zirkulär unter Schonung des Nervus saphenus
eingeschnitten sowie an der medialen und lateralen Oberschenkelseite jeweils
ein weiterer Schnitt bis zum Leistenband durchgeführt. Durch Abziehen des
entstehenden Hautlappens wird der Situs eröffnet und der Nervus saphenus
[9]beziehungsweise Nervus femoralis bis zum Leistenband zugänglich. Alle
weiteren Schritte erfolgen unter mikroskopischer Sicht (SZ-X7, Olympus K. K.,
Tokio, Japan) und ständiger Befeuchtung des Nervenbereichs durch die stets
frisch hergestellte physiologische Interstitialflüssigkeit (SIF-Lösung, Synthetic
Interstitial Fluid-Lösung), um eine Austrocknung und Schädigung des Nervs zu
vermeiden. Unter Zuhilfenahme zweier Uhrmacherpinzetten wird der Nerv
entlang seines Verlaufs am Oberschenkel sorgfältig von den begleitenden
Gefäßen getrennt, ohne dabei das Perineurium zu beschädigen. Am Ende muss
das Nervenende knapp proximal des Leistenbandes dargestellt und mit einem
Faden (chirurgische Seide) umschlungen und verknotet werden. Erst
anschließend wird sowohl der Nerv als auch das Leistenband mit einer
Federschere durchtrennt und der Bereich mit einem befeuchteten, Flusen-freien
Zellstoffstück abgedeckt. Nun muss das durch den Saphenus-Nerv versorgte
Hautareal als Präparat gewonnen werden. Hierfür erfolgt ein Hautschnitt an der
Dorsalkante des Unterschenkels bis zur Ferse an den Übergang von behaarter
zu unbehaarter Haut. Nachfolgend wird durch einen Schnitt am gesamten Fuß
die behaarte von der unbehaarten Haut separiert. Anschließend kann die
behaarte Haut des Fußes vorsichtig mit einer Pinzette bis zum Sprunggelenk
abgehoben und durch sanften Zug bis auf den Hauteintrittspunkt des Nervs von
der Unterschenkel-Muskelfaszie abgelöst werden. Nachdem der Nervus
saphenus in diesem Areal noch behutsam vom umgebenden Bindegewebe
isoliert worden ist, kann das Präparat in ein mit SIF gefülltes kleines Becherglas
überführt und dort bis zur Verwendung (maximal 8 Stunden) gekühlt aufbewahrt
werden. Dieses Vorgehen hat sich in früheren Publikationen als
Versuchsprotokoll bewährt (Zimmermann et al. 2009).
3.3 Einzelfasermessung
Nachfolgend soll zum einen der Versuchsaufbau der Messapparatur näher
vorgestellt und zum anderen auf die eigentliche Einzelfasermessung und
Erhebung der elektrophysiologischen Ableitung eingegangen werden.
[10]3.3.1 Kurzfassung des Versuchsaufbaus
Abbildung 1:
Schemadarstellung des Versuchsaufbaus der Einzelfaserableitung am elektrophysiologischen
Messstand. (I) SIF-Reservoir; (II) hydrostatische Säule mit Flussregelung und Begasungsfritte;
(III) Temperaturregulator zur Applikation von Kaltreizen und Einstellung der Badtemperatur; (IV)
Messkammer mit einem Abteil für das Hautpräparat und einem Abteil für das Splitten des Nerven
auf einem Podest mit Spiegel und die Ableitung von fortgeleiteten Aktionspotenzialen von den
Einzelfasern; (V) Mess- und Neutralelektrode; (VI) Erdung; (VII) Elektro-Stimulationsgerät; (VIII)
Spannungsgerät für Applikation von Wärme-/Hitzereizen; (IX) Überlaufbehälter für SIF.
(Zimmermann et al. 2009)
Zusammenfassend befindet sich innerhalb eines aus Metall bestehenden
Faraday’schen-Käfigs zur Abschirmung von Störsignalen folgender Aufbau: Für
die stetige SIF-Zufuhr existiert ein ca. 1 m oberhalb der Arbeitskammer
gelegenes Reservoir (I). Aus diesem fließt die Flüssigkeit in eine Wassersäule,
in der das SIF kontinuierlich mit Carbogengas begast wird. Die Wassersäule baut
gleichzeitig einen hydrostatischen Druck für das System auf (II). Reguliert über
ein Durchflussventil strömt die SIF von dort durch einen Durchlauferhitzer, der
über eine thermostatische Pumpe betrieben wird und die Badtemperatur von
30 °C aufrechterhält (III), in die Präparatkammer, in der sich das Hautpräparat
befindet. Die Präparatkammer besitzt eine Verbindung mit der Messkammer, die
dem Flüssigkeitsaustausch dient (IV). Unter Zuhilfenahme eines kleinen Wehrs
am Kammerauslass wird der Flüssigkeitsspiegel genauso hoch eingestellt, wie
[11]für eine optimale Messung erforderlich. Das überschüssige SIF fließt über das
Wehr in einen Auslass, der in einem Sammelgefäß endet (IX).
In die Arbeitskammer wird das Hautnerven-Präparat mit der Koriumseite nach
oben eingelegt. Der Nerv des Präparats gelangt durch eine kleine Öffnung in die
Messkammer und wird auf einem dort befestigten Spiegel abgelegt. In der
Messkammer wird auf die vorhandene Flüssigkeit eine Paraffinschicht pipettiert,
sodass der SIF-Spiegel knapp unterhalb der Spiegeloberfläche endet. Dies dient
dazu, eine nicht elektrisch leitfähige und trotzdem durchsichtige
Arbeitsumgebung zu schaffen. In dieser Schicht endet auch die Messelektrode
aus Gold. Eine weitere befindet sich innerhalb der Arbeitskammer als
Neutralelektrode. Diese ermöglichen das Erfassen von elektrischen
Potentialunterschieden zwischen dem Nerven und der Extrazellularlösung mittels
eines Differenzverstärkers (V).
Um die Faserklasse der gemessenen Nerven zu ermitteln, ist die Bestimmung
der Leitungsgeschwindigkeit notwendig, mittels derer auf den Grad der
Myelinisierung zurückgeschlossen werden kann. Hierfür wird eine bewegliche,
an einem Manipulator befestigte, bipolare Mikroelektrode verwendet (VII). Diese
wird von oben möglichst nahe an das rezeptive Feld des zu messenden Nervens
herangeführt und ein elektrischer Impuls mit mindestens doppelter elektrischer
Schwelle abgegeben. Durch wiederholte Messung der Laufzeit des evozierten
propagierten Aktionspotenzials bis zur Nervenantwort am Nervenende (in der
Regel 30 - 100 ms) und der Strecke zwischen Reiz- und Messpunkt kann die
Leitungsgeschwindigkeit berechnet werden.
Eine weitere Vorrichtung dient der Applikation von Kaltreizen mithilfe zweier
Thermostate. Das eine dient der Kühlung der Applikationsflüssigkeit auf 4 - 7 °C,
das andere gewährleistet die Ausgangstemperatur von 30 °C. Durch eine
Rollenpumpe wird eine Flüssigkeit, meist SIF, im Gegenstromprinzip an der
Kühlflüssigkeit vorbeigeführt und auf die gewünschte Temperatur gebracht.
Diese fließt dann über ein kleines Metallrohr in einen zuvor auf das Messareal
(rezeptives Feld und Umgebung) der Haut aufgebrachten Metall-/Teflonring und
wird durch ein zweites Rohr von derselben Pumpe an einem höher gelegenen
Punkt wieder aus dem Ring abgeführt. Ein Temperaturfühler zwischen den
beiden Röhrchen ermöglicht eine zeitgleiche Bestimmung der Ringtemperatur.
[12]Zusätzlich wird eine elektrische Heizvorrichtung verwendet, die das SIF direkt auf
eine bestimmte Temperatur aufheizen kann, um einen Warm- bzw. Hitzereiz zu
applizieren (VIII) (Babes et al. 2010). Die Messapparatur ist detailliert in
(Zimmermann et al. 2009) publiziert.
3.3.2 Datenakquisition
Bevor mit den eigentlichen Ableitungen der Aktionspotentiale (AP) begonnen
werden kann, muss der in der Messkammer einliegende Nervenstumpf von dem
Seidenfaden und eventuell noch vorhanden anderen neuralen Seitenästen
befreit werden. Anschließend wird die Perineuralscheide behutsam am Nerven
zurückgestreift um das nachfolgende Splitten, d.h. das Auffächern des
Nervenbündels in Einzelstränge, zu ermöglichen. Anschließend werden unter
Zuhilfenahme zweier Uhrmacherpinzetten auf dem Spiegel einzelne möglichst
dünne Faserbündel von dem Nerven abgeteilt. Diese werden dann nacheinander
auf die Messelektrode gelegt.
Es folgt die mechanische Reizung der rezeptiven Felder des Hautpräparats mit
einem an der Spitze abgerundeten Glasstab und es werden die propagierten
Aktionspotentiale gemessen. Kommt es zur Identifizierung von mehr als einer C-
Faser im Nervenfaserbündel, so wird das Bündel so lange weiter aufgeteilt, bis
eine einzelne, ein singuläres rezeptives Feld innervierende Nervenfaser
identifiziert werden kann.
Die Bestimmung der mechanischen Reizschwelle erfolgt anschließend durch
Stimulation des rezeptiven Feldes mit von-Frey-Filamenten. Ob ein
Aktionspotential über eine C-Faser geleitet wird, lässt sich über die
Leitungsgeschwindigkeit verifizieren. Entsprechend dem oben genannten
Vorgehen wird der Nerv elektrisch gereizt und die Latenz bis zum
Aktionspotenzial an der Messelektrode ins Verhältnis zur Leitungsstecke gesetzt
(Bessou und Perl 1969). Nachfolgend kann das rezeptive Feld des Nervens
durch einen kleinen Ring (z.B. Teflon; ca. 300 - 400 µl Volumen) von der
restlichen Arbeitskammer abgetrennt werden. Hierfür ist es zwingend notwendig,
diesen koriumseitig durch Auftragen von Vaseline abzudichten, damit keine
Vermischung der in den Ring applizierten und abweichend temperierten
Flüssigkeit mit der Kammerflüssigkeit stattfindet. Dann werden Kaltreize durch
Zuleitung des gekühlten SIF in den Ring appliziert, wie oben beschrieben. Die
[13]Reizdauer beträgt jeweils 60 s, dadurch wird das rezeptive Feld von 30 °C auf
ca. 5 °C abgekühlt. Reagiert eine mechanosensitive Faser während der
Abkühlperiode mit mindestens 3 Aktionspotentialen und zeigt zwischen den
Reizen bei 30 °C Badtemperatur keine spontanen Aktionspotenziale, so wird die
Faser als Kaltnozizeptor (CMC; C-fiber Mechano-Cold) klassifiziert. Um diese
von polymodalen C-Fasern zu unterscheiden, die sowohl mechanosensitiv als
auch thermosensitiv für Kälte- und Hitzereize sind (CMCH; C-fiber Mechano-
Cold-Heat), wird abschließend ein Hitzereiz appliziert, indem das zugeleitete SIF
kontrolliert über den Durchlauferhitzer auf bis zu 50 °C erhitzt wird. Erzeugt die
Faser mehrere Aktionspotenziale auch während der Hitzerampe und steigt die
Momentanfrequenz mit steigender Temperatur oder entsteht zumindest 1 AP am
heißesten Punkt der Rampe, so gilt dies als Hitzeantwort und die C-Faser wird
als CMCH-Subtyp klassifiziert. Entstehen jedoch Aktionspotentiale als einzelner
Burst in der Mitte der Hitzerampe, deutet dies eher auf ein paradoxe, nicht aber
auf eine systematische Hitzeantwort hin, die bei CMC-Fasern vorkommen kann.
Um monomodale Kaltthermozeptoren (CC-Fasern; C-fiber Cold) zu identifizieren,
wird zur Suche anstelle eines mechanischen Reizes (Glasstab) ein thermischer
Reiz verwendet, da diese Fasern keine mechanische Sensibilität besitzen.
Hierfür wird ein kleines Eisstück nahe an der Koriumoberfläche entlang des
Präparates geführt und resultierende AP auf diesen Kaltreiz detektiert. Diese
erfolgen in der Regel mit Spitzenentladungsraten von 50 - 100 AP/s.
Anschließend erfolgt eine Klassifizierung des Nervens. Sollte dieser eine
Leitungsgeschwindigkeit von < 1 m/s haben und nur auf kalte thermische Reize
reagieren, kann er als CC-Faser eingestuft werden. CC-Fasern sind sehr dünne
Nervenfasern, die aus den kleinsten sensiblen Neuronen des Rückenmarks
entspringen (Vriens et al. 2014) und daher eine besondere Herausforderung im
Rahmen der Einzelfasermessung darstellen. Denn je dünner eine Faser ist, desto
besser muss der Kontakt zur Elektrode sein, um das propagierte AP noch mit
einem adäquaten Signal-Rausch-Verhältnis aufzeichnen zu können. Dies ist nur
dann machbar, wenn es gelingt, die Faserbündel sehr fein aufzusplitten. Um die
Inzidenz dieser Fasern zwischen den drei untersuchten Genotypen abschätzen
zu können, wurden daher möglichst dünne Nervenfaserbündel auf rein
kaltsensitive rezeptive Felder getestet.
[14]Die messbaren Aktionspotentiale stellen dabei kleine Ladungsdifferenzen
zwischen der Normal-/Referenzelektrode in der Arbeitskammer und der
Messelektrode in der Messkammer dar. Diese geringen elektrischen Potentiale
werden anschließen verstärkt (Verstärker; Franken Elektronik, Röthenbach) und
akustisch über angeschlossene Lautsprecher beziehungsweise visuell über ein
Oszilloskop (HM507, Hameg, Mainhausen) wahrnehmbar gemacht. Nach
Aufarbeitung der Signale durch ein zwischengeschaltetes Wandler- oder DAP-
Board (DAP5200a, Microstar Laboratories, Redmond, WA) wird das Signal in
einen PC-Slot eingespeist und hier mittels der Software DAPSYS (Data
Aquisition Processor System; Version 8, Brian Turnquist) zur nachfolgenden
Analyse aufgezeichnet. Hierfür werden neben den Aktionspotentialen auch
synchron die durch den Temperaturfühler aufgezeichneten Ring-Temperaturen
erfasst. Letztendlich entsteht ein Datensatz, der die Reizantwort in Abhängigkeit
zur Temperatur im zeitlichen Verlauf enthält.
Ein vereinfachtes Schema des Versuchsaufbaus zur Datenerfassung ist in
Abbildung 2 dargestellt und detailliert in (Zimmermann et al. 2009) dargelegt.
[15]Abbildung 2:
Schemadarstellung des Versuchsaufbaus mit Datenerhebung. STIMULATOR: Möglichkeit zur
elektrischen Reizung der Kutis des Hautpräparats über eine bipolare Elektrode, Signalerfassung
mittels DAP-Board. VERSTÄRKER: Amplifizierung des durch Nervenreizung erzeugten
elektrischen Potentials, Akquise via DAP, Ausgabe als akustischen Signal und Weiterleitung zum
Oszilloskop. OSZILLOSKOP: Darstellung des verstärkten elektrischen Aktionspotentials als
Spannungskurve. COMPUTER: Erfassung und Verarbeitung der Reizantworten,
Temperaturkurven und Signale des elektrischen Stimulators. Thermostatpumpe und
Kühlthermostatpumpe: Erzeugung von kalten/kühlen oder warmen/heißen Temperaturreizen in
Kombination mit dem Netzteil, das den Strom durch den Durchlauferhitzer steuert (PeakTech
Prüf- und Messtechnik, Ahrensburg), und der Rollenpumpe, die die temperierte Flüssigkeit in das
rezeptive Feld führt. Die Erfassung der Temperatur des rezeptiven Felds erfolgt über ein
Thermometer an der Öffnung des Applikationssystems. (Zimmermann et al. 2009)
3.4 Synthetic Interstitial Fluid (SIF)
Für die Nervenpräparation und die Untersuchung im Einzelfaser-Setup wird
künstlich hergestellte und oxygenierte Zwischenzellflüssigkeit benötigt, die den
Nerven und das Hautpräparat während des Experiments am Leben erhält. Für
diese, an jedem Untersuchungstag frisch hergestellte Lösung, werden in
[16]ultrareinem Wasser folgende Chemikalien (Firma Sigma-Aldrich, St. Louis, MO)
gelöst (Zimmermann et al. 2009; Babes et al. 2010).
[mM] Reagenz
9,64 ..... D-Gluconsäure-Natriumsalz (C₆H₁₂O₇ ∙ Na; PNr G9005)
5,55 ..... D-Glucose (C6H12O6; PNr G7021)
3,5 ..... Kaliumchlorid (KCl; PNr P9333)
0,69 ..... Magnesiumsulfat-heptahydrat (MgSO4 ∙ 7H2O; PNr M1880)
26,2 ..... Natriumbicarbonat (NaHCO3; PNr S5761)
1,67 ..... Natriumdihydrogenphosphat-Dihydrat (NaH2PO4 ∙ 2H2O; PNr 71500)
107,8 ..... Natriumchlorid (NaCl; PNr S9888)
7,6 ..... Saccharose (C₁₂H₂₂O₁₁; PNr S7903)
1,53 ..... Calciumchlorid-Dihydrat* (CaCl2 ∙ 2H2O; PNr C5080)
* Die letztgenannte Substanz wird erst nach einer Begasung der Lösung mit Carbogengas
(5 Vol% CO2 und 95 Vol% O2) von mindestens 5 min Dauer nach Erreichen eines pH-Werts von
7,4 hinzugefügt. Diese Carbogenisierung wird auch während des gesamten Experiments
aufrechterhalten.
3.5 Statistische Analyse / Datenanalyse
Zur Analyse der erfassten Daten und anschließenden statistischen Auswertung
wurde das Programm IBM SPSS Statistics Version 21genutzt.
Da von einer Normalverteilung der gewonnenen Daten ausgegangen werden
kann, wird zur Identifizierung von Ausreißern der Test nach Grubbs verwendet.
Diese sind in den Abbildungen 4 bis 6 durch Sterne dargestellt. Die weiteren
Berechnungen zur Erstellung der Abbildungen wie Mittelwert oder Quantile
wurden ohne diese Ausreißer vorgenommen.
Zum Vergleich von Datensätzen mit zwei Gruppen wurde der gepaarte oder
ungepaarte t-Test verwendet. Wurden mehr als zwei Gruppen miteinander
verglichen, erfolgte eine einfaktorielle Varianzanalyse (ANOVA) mittels Fisher-
LSD-post-hoc-Test. Zur Untersuchung der Unabhängigkeit der 3
Vergleichsgruppen (relative Faseranzahl; Abbildung 5) wurde der Chi-Quadrat-
Unabhängigkeitstest angewendet.
Signifikanz wurde ab p < 0,05 angenommen. Entsprechende Datenpunkte
wurden durch ein einzelnes Symbol (*) gekennzeichnet. Zur weiteren Abstufung
[17]erhielten Datenpunkte mit einem P-Wert von p < 0,01 zwei Symbole (**) und mit
p < 0,001 drei Symbole (***) als Markierung.
Die Fehlerbalken stellen entweder Standardabweichungen oder Standardfehler
dar und sind in den Abbildungslegenden entsprechend bezeichnet.
Die durch DAPSYS akquirierten Daten wurden mittels Microsoft Excel (Microsoft
Corporation, Redmond, WA) weiterverarbeitet. Grafiken der Abbildungen 4 - 6
und Diagramme wurden mit OriginPRO (OriginLab Corporation, MA) erstellt und
unter Verwendung von CorelDRAW (Corel Corporation, Kanada) auf den Layout
Stil des Journals „Frontiers in Molekular Neuroscience“ angepasst. Tabelle 1
wurden mittels Microsoft Excel (Microsoft Corporation, Redmond, WA) und die
Abbildungen 1 - 3 wurden mittels Microsoft PowerPoint (Microsoft Corporation,
Redmond, WA) bearbeitet.
3.6 Temperatur-Gradienten-Messgerät
Für das zu dieser Abhandlung gehörende Paper (Winter et al. 2017) wurde die
größere Variante des von unserer Forschungsgruppe neu konzipierten runden
Temperaturgradienten-Assays (Innendurchmesser 45 cm; Außendurchmesser
57 cm; Ringoberfläche 960 cm2) verwendet. Die Gerätschaft modifiziert den in
früheren Publikationen verwendeten linearen Temperatur-Assay durch Kreisform
mit dem Vorteil fehlender Ecken, die meist schlechter ausgeleuchtet sind und die
Mäuse dazu bewegen, sich dort bevorzugt niederzulassen. Durch einseitige
Kühlung und gegenüberliegende Erwärmung bilden sich zwei Ringhalbkreise mit
einem symmetrischen Temperaturgradienten zwischen angelegter Minimal- und
Maximaltemperatur. Die Halbkreise werden in 12 Temperaturfelder unterteilt, die
sich jeweils um wenige Grad Celsius (1/12 der Differenz von Minimal- und
Maximaltemperatur) unterscheiden und so die Wahrnehmung von
Temperaturunterschieden systematischer testen als Zwei-Platten-Wahltests
oder der lineare Temperaturgradient.
Dieses zirkuläre Testgerät zeigte ein signifikant verringertes
Kältevermeidungsverhalten der DKO-Tiere gegenüber den TRPM8-/- und, noch
ausgeprägter, gegenüber dem Wildtyp. Quantitativ zeigte sich dieser Unterschied
in einer reduzierten gemittelten Präferenztemperatur gegenüber dem Wildtyp
beziehungsweise dem TRPM8-/- in den letzten 15 min der Untersuchung (Minuten
[18]46 - 60). Die gewichteten mittleren Vorzugstemperaturen lagen beim DKO,
ebenfalls unterhalb derer der Wildtypmäuse und der TRPM8-/--Tiere. Die
Funktionsweise dieses neuartigen Geräts sowie der Auswertungsprozess der
gewonnenen Daten kann in (Touska et al. 2016) eingesehen werden.
4. Physiologischer Hintergrund
Zum Verständnis der Forschungsarbeit sind einige physiologische Grundlagen
notwendig, um die Prinzipien der Messungen nachvollziehen zu können.
4.1 Transient-Rezeptor-Potential Ionenkanäle (TRP-Ionenkanäle)
Die untersuchten Kanäle sind physiologischer Weise als Transmembranproteine
zum Beispiel an peripheren Endigungen sensorischer Nerven, aber auch vielen
anderen Zellen in die Zellmembran integriert (González-González et al. 2017).
Neben Säugetieren wurden sie auch in Fruchtfliegen (Drosophila melanogaster)
und Würmern (Caenorhabditis elegans) nachgewiesen, was ihre große
Bedeutung unter evolutionärem Druck verdeutlicht (Cosens und Manning 1969;
Montell und Rubin 1989; Baraldi et al. 2010).
Zur Gruppe der TRP-Kanäle zählen insgesamt fast 30 Mitglieder. Diese gliedern
sich in sieben Subgruppen mit meist mehreren Vertretern und verschiedenen
Aminosäuresequenzen. Zu ihren Gemeinsamkeiten zählt unter anderem, dass
sie jeweils sechs Transmembranregionen (S1 - S6) besitzen, als Homo-
/Heterotetramer in die Zellmembran eingebaut sind und bei Aktivierung
unselektiv für Kationen, vor allem Na+, Ca2+ und Mg2+, durchlässig werden
(Montell 2005; Clapham 2003; Baraldi et al. 2010). Die TRP-Kanäle erlauben bei
ihrer Aktivierung durch einen bestimmen physikalischen Reiz oder Botenstoff
(Liganden) einen transienten, also vorübergehenden, Einstrom von
monovalenten (Na+) oder bivalenten Kationen (Ca2+, Mg2+) in die entsprechende
Zelle (Minke et al. 1975). Dieses namensgebende Kanalverhalten wurde
erstmalig bei Drosophila-Mutanten beschrieben, die im Gegensatz zum Wildtyp
eine transientes Rezeptorpotential auf Lichtreize zeigten (Cosens und Manning
1969). Aus dem Kationen-Einstrom ergibt sich eine Depolarisation der (Nerven-)
Zelle und dadurch beispielsweise eine Aktivierung spannungsabhängiger
Natrium- oder Calcium-Kanäle beziehungsweise weiterer intrazellulärer
Signalkaskaden, zum Beispiel G-Protein-gekoppelter Signalwege (Hofmann et
[19]al. 1999). So dienen sie der Wahrnehmung chemischer Substanzen wie
Geschmacksstoffen, noxischen Reizen oder Pheromonen sowie physikalischer
Reize wie Temperaturveränderungen, mechanischem oder osmotischem Druck
oder auch Licht.
TRP-Kanäle sind also multimodal, d.h. sie reagieren auf verschiedene
Reizqualitäten (Clapham 2003) und spielen damit eine wichtige Rolle für die
Sinnes- und Schmerzwahrnehmung (Vriens et al. 2014; Shigetomi et al. 2013;
Pogorzala et al. 2013; Palkar et al. 2015).
Von den bislang entdeckten knapp 30 TRP-Kanälen sind bislang 12 Vertreter aus
vier Subgruppen als thermosensitiv beschrieben. Hierunter zählen die Vanilloid-
Rezeptorsubtypen TRPV1, TRPV2, TRPV3 und TRPV4, die Melastatin-
ähnlichen Rezeptoren der Subtypen TRPM2, TRPM3, TRPM4, TRPM5 und
TRPM8, der einzige bisher bekannte Vertreter der Ankyrin-reichen TRP-Kanäle
TRPA1 und der klassische TRP-Rezeptor (C für classical oder canonical) TRPC5
(Wang und Siemens 2015). Von den zuvor genannten Kanälen wurden TRPM8,
TRPA1 und TRPC5 als Kaltsensoren beschrieben, die übrigen vermitteln
Hitzereize. Der erste als thermosensitiv identifizierte Kanal war TRPV1, welcher
unter anderem auf Hitze von > 42 °C, Capsaicin und saure pH-Werte reagiert,
also ebenfalls polymodal ist (Caterina et al. 1997; Caterina et al. 2000; Davis et
al. 2000). Daneben stellt TRPM8 den am besten erforschten Kaltrezeptor unter
den TRP-Kanälen dar. Der Kanal kommt sowohl als alleiniger TRP-Subtyp in
Nervenfasern, als auch gemeinsam, zum Beispiel mit TRPV1, in nozizeptiven
Fasern vor (Peier et al. 2002; McKemy et al. 2002).
Darüber hinaus spielen TRP-Kanäle im Zentralen Nervensystem (ZNS) eine
wichtige regulatorische Rolle. Sie vermitteln über Signalkaskaden neuronales
Wachstum, Vesikel-Ausschüttung, Einbau neuer Proteine in die Zellmembran
oder auch programmierten Zelltod (Moran et al. 2004).
In dieser Arbeit wurde speziell die Funktion zweier Vertreter dieser
Membrankanalklasse untersucht, die eine entscheidende Rolle bei der Kühl-
bzw. Kaltwahrnehmung in den Organismen einnehmen.
[20]4.1.1 TRPA1
Der TRPA1-Kanal oder Synonym ANKTM1 (Ankyrin-Transmembranprotein 1) ist
der einzige bislang identifizierte Vertreter der Ankyrin-Unterfamilie der TRP-
Kanäle (Jaquemar et al. 1999; Clapham et al. 2005). Das namensgebende
Charakteristikum ist das gehäufte Auftreten von Ankyrin-Wiederholungen am N-
Terminus des Proteins (García-Añoveros und Nagata 2007; Story et al. 2003). Er
ist ein typischer Nozizeptor-Kanal, der der Schmerzwahrnehmung dient und der
durch verschiedene chemische Liganden aktiviert werden kann (Macpherson et
al. 2007b; Tai et al. 2008; Zhang et al. 2008; Jordt et al. 2004; Andersen et al.
2015). Bei seiner näheren Betrachtung konnte nachgewiesen werden, dass viele
Liganden eine kovalente Bindung mit einer Cystein-Gruppe des Kanals eingehen
und so über Konfigurationsänderung des Proteins zur Aktivierung führen
(Macpherson et al. 2007a; Bautista et al. 2006). Ebenso ist eine Aktivierung durch
mechanische, osmotische oder thermische Reize beschrieben (Moparthi et al.
2014; Kwan et al. 2006; Karashima et al. 2009; Story et al. 2003; Baraldi et al.
2010), wobei gerade die Kalttransduktionsfunktion des Kanals für lange Zeit in
der Literatur als widersprüchlich galt. Einigen dafürsprechenden Studien (Story
et al. 2003; Kwan et al. 2006; Bandell et al. 2004; Karashima et al. 2009; Moparthi
et al. 2014; Sawada et al. 2007; Klionsky et al. 2007) stehen gegenteilige
Forschungsergebnisse gegenüber, die dem Kanal diese Funktion absprechen
(Jordt et al. 2004; Nagata et al. 2005; Bautista et al. 2006). Neuere Ergebnisse
deuten auf eine zusätzliche Funktion von TRPA1 als Hitzetransduktionskanal hin
(Moparthi et al. 2016; Moparthi et al. 2014; Vandewauw et al. 2018).
Im menschlichen Organismus lässt sich der TRPA1-Kanal in sensorischen
Nervenendigungen der Haut finden. Darüber hinaus konnte auch ein Vorkommen
in verschiedenen weiteren Organen gezeigt werden, unter anderem Gefäßen,
Lunge oder viszeralen Organen (Nagata et al. 2005). Er führt bei Aktivierung zu
Vasodilatation, lokalen Inflammation, Ödembildung sowie Hypersensibilisierung
umgebender Nerven (Klionsky et al. 2007; Nagata et al. 2005).
Darüber hinaus scheint der TRPA1-Kanal an den antinozizeptiven Eigenschaften
von Paracetamol durch Bindung eines Metaboliten des Medikaments an den
Kanal mit nachfolgender Desensitivierung beteiligt zu sein (Andersson et al.
2011). Über Bindung von Metaboliten des Oxaliplatin (Oxalate), einem
insbesondere in der Therapie des kolorektalen Karzinoms eingesetzten
[21]Zytostatikum, wird eine häufige Nebenwirkung dieser antineoplastischen
Therapie ausgelöst, die kaltinduzierten akuten peripheren Neuropathie (Miyake
et al. 2016).
In dieser Arbeit wurde der Einfluss dieses Kanals auf die
Temperaturwahrnehmung der Mäuse im Einzelknockout und im Doppelknockout
bei gleichzeitigem Fehlen des Weiteren Kalttransduktionskanals TRPM8
untersucht. In früheren Studien zeigten TRPA1-/- eine herabgesetzte Chemo-
(Bautista et al. 2006; Kwan et al. 2006), Mechano- (Kwan et al. 2006; Petrus et
al. 2007) und, wie zuvor bereits erwähnt, Kaltsensibilität (Karashima et al. 2009;
Vetter et al. 2012). Außerdem konnte ein Ausbleiben der mechanisch-induzierten
Hyperalgesie der Versuchstiere gezeigt werden (Kwan et al. 2006).
Doppelknockouts hingegen ließen im 2-Platten-Temperaturwahltest keinen
Schluss auf einen quantitativ über das Defizit der TRPM8-Knockouts
hinausgehend veränderten Phänotyp in der Kaltwahrnehmung zu (Knowlton et
al. 2010).
4.1.2 TRPM8
Der TRPM8-Kanal, einer von insgesamt acht Mitgliedern der Melastatin-
Unterfamilie der TRP-Kanäle (Kraft und Harteneck 2005; Boesmans et al. 2011),
reagiert auf thermische Reize im kühlen Bereich mit vorrübergehender Öffnung
(Peier et al. 2002) und wird durch Menthol oder Icilin chemisch aktiviert sowie
sensibilisiert (Andersson et al. 2004; Bandell et al. 2006; Brignell et al. 2008;
Behrendt et al. 2004; McKemy et al. 2002). Daher stammt auch der Rezeptor-
Zweitname CMR1 (Englisch „cold and menthol receptor 1“; Kalt- und Menthol-
Rezeptor 1).
Das Kanalprotein besitzt 6 Transmembranproteine (Pedretti et al. 2009) und wird
als Homotetramer (Jiang 2007), also mit vier identischen Untereinheiten, in die
Zellmembran von sensorischen Neuronen eingebaut. Er ließ sich ebenfalls in
Zellen der Prostata (Zhang und Barritt 2004) oder Lunge nachweisen (Sabnis et
al. 2008). Als nicht-selektiver Kationen-Kanal liegt eine Durchlässigkeit für positiv
geladene Ionen vor, vor allem Natrium- und Calcium-Ionen (Pedretti et al. 2009).
Der Knockout des Kanals führt mit etwa Halbierung der Anzahl kaltsensitiven
DRG-Neurone (dorsal root ganglion; Spinalganglion) sowie abgeschwächten
[22]Kaltvermeidungsverhalten für nicht-noxisch kalte Temperaturen von 23 - 30 °C
(Dhaka et al. 2007) beziehungsweise kaltsensibler TG-Neurone
(Trigeminalganglion) auf etwa ein Fünftel (Bautista et al. 2007) zwar zu Defiziten
in der Kaltwahrnehmung, jedoch nicht zum vollkommenen Wegfall der
Wahrnehmung von insbesondere noxischer Kälte.
Die verbliebenen Kaltfasern zeigten Eigenschaften der Menthol-insensitiven
Fasern des Wildtyps mit Schwellentemperaturen für die Kaltaktivierung um 12 °C
(Bautista et al. 2007). Zudem ließ sich für Temperaturen < 20 °C ein
unverändertes Kaltvermeidungsverhalten der TRPM8-Knockouts im Vergleich
zum Wildtyp mittels linearen Temperaturgradienten-Assay nachweisen (Dhaka
et al. 2007).
Bei Einzelfasermessungen an TRPM8-Knockoutmäusen konnte zusätzlich eine
weitere Faserpopulation entdeckt werden, die zwar mechanosensitiv ist,
allerdings eine größere Kaltantwort zeigt als die noch vorhandenen polymodalen
CMCH-Fasern. Diese besitzt eine Schwellentemperatur von circa 30 °C und
wurde wohl aus diesem Grund primär bei normaler Badtemperatur nicht entdeckt,
sondern erst nach Anhebung der Temperatur im Rahmen der Suche nach CC-
Fasern. Sie zeigen eine starke dynamische Kaltreaktion und ihre
Gesamtkaltantwort ist mit der von Menthol-positiven Kaltfasern des Wildtyps
vergleichbar (26,4 ± 10,4 AP/60s gegenüber 24,7 ± 17,5 AP/60s; (Winter et al.
2017)). Während sie eine deutlich höhere Maximalfeuerrate im dynamischen
Antwortanteil aufweisen, ist deren statische Antwort nur etwa ein Viertel (~23 %)
so groß wie die der vergleichbaren Wildtyp-Fasern (Bautista et al. 2007; Toro et
al. 2015).
Die anderen Subtypen der TRPM-Kanäle haben andere Funktionen wie z.B.
Geschmackswahrnehmung (Moran et al. 2004), Neuriten-Wachstum und
axonales Wachstum in Assoziation mit Krankheiten wie Amyotropher
Lateralsklerose oder Parkinsondemenz (Hermosura et al. 2005), Nierenfunktion,
insbesondere Rückresorption von Magnesium und Calcium (Walder et al. 2002),
Mediation der Kontraktion von Herz- und Gefäßmuskulatur, z.B. zur Veränderung
des Gefäßdiameters im Rahmen des perfusionsregulierenden Bayliss-Effekts
oder der Blutdruckregulation (Touyz et al. 2006).
[23]Sie können auch lesen

















































