Genetische Untersuchungen der Nasen in der Zürichseeregion
←
→
Transkription von Seiteninhalten
Wenn Ihr Browser die Seite nicht korrekt rendert, bitte, lesen Sie den Inhalt der Seite unten

Genetische
Entwurf :01.11.2021 Untersuchungen der Nasen in der
Zürichseeregion
Schlussbericht
1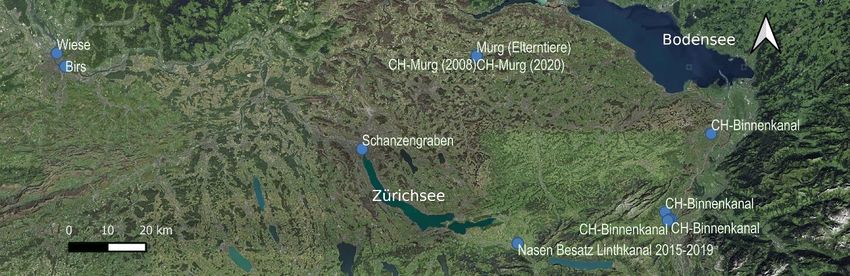
Genetische Untersuchungen der Nasen in der Zürichseeregion
Impressum
Auftraggeber
Die Fischereikommission für den Zürichsee,
Linthkanal und Walensee
Auftragnehmer
Aquabios GmbH
Les Fermes 57
1792 Cordast
info@aquabios.ch
Autoren
Axel Zander
Pascal Vonlanthen
Begleitung:
Michael Kugler
Lukas Bammatter
Kuno von Wattenwyl
Titelbild
Nasen mit Laichausschlag während der Fortpflanzung (Foto © Michel Roggo).
Zitiervorschlag
Zander, Vonlanthen (2021). Genetische Untersuchungen der Nasen der Zürichseeregion. Aquabios GmbH.
Auftraggeber: Die Fischereikommission für den Zürichsee, Linthkanal und Walensee.
Verdankungen
Wir bedanken uns bei Michael Kugler, Lukas Bammatter und Kuno von Wattenwyl für den Auftrag und die
interessanten Diskussionen. Thomas Kreienbühl danken wir für die Hilfe beim Scoren der Daten.
2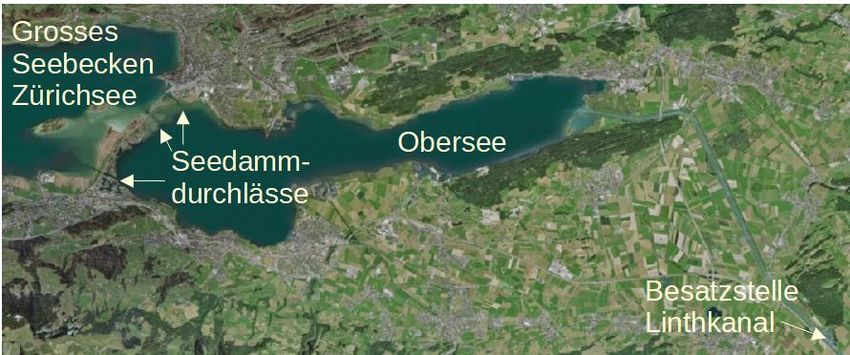
Genetische Untersuchungen der Nasen in der Zürichseeregion
Inhaltsverzeichnis
ZUSAMMENFASSUNG ..................................................................................................................................................4
AUFTRAG .....................................................................................................................................................................5
AUSGANGSLAGE .................................................................................................................................................................. 5
BIOLOGIE DER NASEN ........................................................................................................................................................... 5
GENETIK ............................................................................................................................................................................ 6
HISTORISCHE VORKOMMEN DER NASE IM ZÜRCHSEEGEBIET UND BESATZHISTORIE. .......................................................................... 6
AKTUELLE DURCHGÄNGIGKEITSSITUATION DER LIMMAT VON DER AARE BIS IN DEN ZÜRICHSEE ........................................................... 7
FÖRDERMASSNAHMEN IM ZÜRICHSEE-EINZUGSGEBIET ............................................................................................................... 8
FRAGESTELLUNGEN .............................................................................................................................................................. 9
MATERIAL UND METHODEN ...................................................................................................................................... 10
NASENFÄNGE IM ZÜRICHSEE 2021 ....................................................................................................................................... 10
VORGEHENSWEISE ZUM VERGLEICH DER POPULATIONEN .......................................................................................................... 11
LABORARBEITEN ................................................................................................................................................................ 12
STATISTISCHE AUSWERTUNGEN UND TESTS ............................................................................................................................ 12
ERGEBNISSE ............................................................................................................................................................... 13
VERGLEICHE ZWISCHEN DEN POPULATIONEN........................................................................................................................... 14
HERKUNFT DER ZÜRICHSEENASEN ......................................................................................................................................... 16
DISKUSSION ............................................................................................................................................................... 17
EMPFEHLUNGEN ........................................................................................................................................................ 18
LITERATURVERZEICHNIS ............................................................................................................................................ 19
3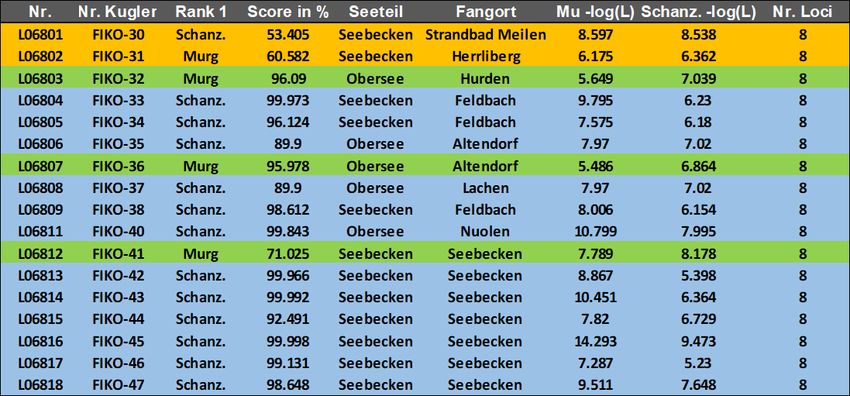
Genetische Untersuchungen der Nasen in der Zürichseeregion
Zusammenfassung
Im Zeitraum von 1995-2004 konnten im Zürichsee keine Nasen mehr nachgewiesen werden. Nur im Schan-
zengraben und der oberen Limmat (sowie der unteren Sihl) konnte sich eine Population halten. Diese war
aber klein und aufgrund der Passage durch einen sogenannten Flaschenhals genetisch verarmt. Nach Fertig-
stellung der Fischtreppe im Schanzengraben, konnte sich diese Population zumindest in der Theorie in den
Zürichsee ausbreiten. Seit 2015 wurde im Linth-Einzugsgebiet mit Besatzmassnahmen versucht die Nase wie-
deranzusiedeln.
2020 wurden im Zürichsee überraschend viele Nasen gefangen. Von einigen wurden Proben gesammelt und
genetisch untersucht. Es stellte sich die Frage, ob die gefangenen Nasen von den Besatzmassnahmen ab-
stammten oder natürlich in den See eingewandert sind. Im Rahmen der vorliegenden Studie sollte überprüft
werden, ob die Nasen des Zürichsees aus dem Schanzengraben oder von der Wiederansiedlung mit Murg-
Nasen stammen. Genetische Populationszuweisungen wurden verwendet, um die im Zürichsee gefangenen
Nasen den beiden möglichen Populationen zuzuweisen.
Von den 17 untersuchten Fischen liessen sich 12 klar zur Gruppe der Schanzengrabennasen zuweisen. Dies
bedeutet, dass ein Grossteil der im Zürichsee gefangenen Nasen natürlicherweise in den See eingewandert
ist. Diese Schanzengrabennasen waren im Untersee häufiger anzutreffen als im Obersee. Insgesamt drei In-
dividuen konnten der Mittellandpopulation zugewiesen werden, die auch bei der Wiederansiedlung im Linth-
gebiet eingesetzt wurden. Im Obersee konnten zwei von fünf untersuchten Proben der Mittellandpopulation
zugwiesen werden. Es ist wahrscheinlich, dass diese Individuen von den Wiederansiedlungsmassnahmen ab-
stammen.
Schliesslich konnten zwei Individuen nicht eindeutig einer Population zugewiesen werden. Dies weil sie ge-
netisch intermediär zwischen den beiden Populationen lagen. Diese Individuen stammen also weder aus dem
Schanzengraben noch von den Wiederansiedlungen ab. Wir vermuten, dass es sich um Limmatnasen handelt.
Ein genetischer Vergleich mit Limmatnasen könnte Klarheit schaffen.
4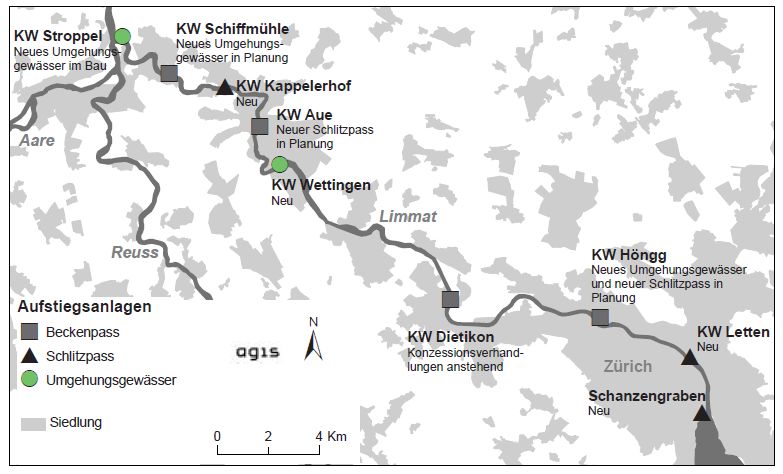
Genetische Untersuchungen der Nasen in der Zürichseeregion
Auftrag
Ausgangslage
Nasen (Chondrostoma nasus) sind Algenfresser und gehören somit zu den wenigen Primärkonsumenten un-
ter den Europäischen Fischen (Pisci Page, abgerufen am 08.07.2021). Adulte Nasen grasen fast ausschliesslich
Kieselalgen vom groben Flusssediment ab (Adamik und Obrdlik, 1977; Dedual, 1990). In den früheren, natur-
nahen Schweizer Gewässern gehörte die Nase zahlenmässig zu der häufigsten Fischart der Barbenregion
(Vonlanthen et al., 2010). Da sie sehr anfällig gegenüber Gewässerverunreinigungen aller Art ist, ist Ihr Be-
stand im 20. Jahrhundert stark zurückgegangen (Maier et al., 1995).
Als weitere Gründe für den starken Rückgang der Nasenpopulationen in den Schweizer Gewässern nennt der
WWF Schweiz (2007) Wanderhindernisse (Wehre), Kolmation (verstopftes Kiesbett) und die morphologische
Degradierung von Flüssen und Bächen z.B. Eindolung und Begradigung. Die Aare und der Rhein, früher die
Hauptlebensräume der Nase in der Schweiz, wurden fast überall in Ketten von Staustufen umgebaut, welche
die Wanderung der Mitteldistanzwanderer wie die Nase behindern und die Ablagerung von Feinsedimenten
erhöhen. Da die Nase steiniges Substrat und starke Strömung für die Fortpflanzung brauchen, fehlen ihr in
den Grossen Flüssen die Laichplätze. Auch Jungfischlebensräume, wie Flachufer und Altarme sind nicht mehr
ausreichend vorhanden. Nicht zu vernachlässigen sind auch zu geringe Restwassermengen und Schwall-Sunk
Betrieb unterhalb von Kraftwerken (WWF Schweiz, 2007).
Aus diesen Gründen waren die Nasenbestände im Zürichseegebiet zeitweise wohl komplett erloschen, bzw.
wurden als erloschen angegeben (Maier et al., 1995; INFO FAUNA http://lepus.unine.ch/carto/70042 abge-
rufen am 01.07.2021; Dönni & Spalinger, 2017; Zbinden et al., 2005). Die Nase ist seit 1. Januar 2007 in der
Schweiz ganzjährig geschützt und darf nicht gefangen werden (VBGF v. 24.11.93, Stand 1.7.08). Dennoch
gehört die Nase heute zu den meistgefährdeten Fischarten in Mitteleuropa, weshalb sie dem höchsten Ge-
fährdungsgrad 1E (1 = vom Aussterben bedroht; E = europäisch geschützt) nach der Berner Konvention un-
terliegt (Rey, 2019).
Biologie der Nasen
Nasen gehören zu den Karpfenfischen (Cypriniden), sie laichen im Frühling (Zbinden S. et al., 2005) zwischen
März und Mai (Bonell, 2009) und legen ihre Eier bevorzugt in 30- 50cm Tiefe auf grobem Kies ab (Dedual,
1990). Vorher kommt es zu Laichwanderungen, die mehrere hundert Kilometer lang sein können (Freyhof
1998) und entsprechend viel Zeit in Anspruch nehmen. Nach dem Ablaichen kehren sie in ihre angestammten
Flussabschnitte zurück und leben dort recht stationär (Fielenbach, 1996, Huber & Kirchhofer, 1997). In diesen
Bereichen halten sich Adulte Nasen bevorzugt an Stellen mit Fliessgeschwindigkeiten zwischen 0.5 und 1 m
pro Sekunde und in Tiefen von 1-2 Meter über grobem Sediment auf (Huber & Kirchhofer, 1997). Die Nase
lebt gesellig, als friedlicher Schwarmfisch, vorzugsweise in grösseren Flüssen (Bonell, 2009). Innerhalb von 4
bis 7 Jahren wird eine Grösse von 30-50cm und die Geschlechtsreife erreicht. Ihr Höchstalter in der Natur
liegt zwischen 15 und 20 Jahren, (Bonell, 2009). In den ersten Lebensmonaten ernähren sich die Nasenlarven
von Rädertierchen und Mückenlarven. Erst danach gehen sie zu einer pflanzlichen Diät über, die überwiegend
aus Diatomeen besteht. (Schneider, 1992, Zbinden S. et al., 2005). Im ersten und zweiten Lebensjahr über-
wintern sie in speziellen Winterhabitaten, die oft sehr weit flussabwärts ihrer Schlupforte liegen (Freyhof,
1997). Sie sind daher darauf angewiesen, dass die Flüsse in denen sie leben flussauf- und flussabwärts durch-
gängig sind.
5Genetische Untersuchungen der Nasen in der Zürichseeregion
Nasen werden nach 4-7 Jahren geschlechtsreif (Zbinden et al., 2005) und benötigen in ihrem Lebenszyklus
einen gut vernetzten und weitverzweigten Lebensraum. Wenn die Gewässerverbauungssituation es zulässt,
können Laichwanderungen von mehreren hundert Kilometern durchgeführt werden. Auf diese Weise kann
die Nase alle Teillebensräume nutzen, die sie im Jahres- und Altersverlauf benötigt. Auch grosse Seen, wie
der Zürichsee können auf diesen Wanderungen durchschwommen werden und sind deshalb Teil des natürli-
chen Lebensraums der Nasen.
Genetik
Bisherige populationsgenetische Untersuchungen von Schweizer Nasenpopulationen haben gezeigt, dass im
natürlichen Verbreitungsgebiet der Nase zwei Metapopulationen existieren. Eine im Oberrhein / Hochrhein
(unterhalb des Rheinfalls) und darin mündenden Nebenflüssen (Aare, Murg, Thur, Wiese, Birs, Saane etc.).
Die andere Metapopulation existiert im Bodensee und Alpenrhein und wurde, da der Rheinfall zu keiner Zeit
von flussaufwärts ziehenden Nasen passierbar war, wahrscheinlich über natürliche Flusslaufdurchbrüche
vom Donau- zum Bodenseeeinzugsgebiet her besiedelt (Vonlanthen et al., 2010). Nasen aus dem Schanzen-
graben wiesen in der Studie von Vonlanthen et al. (2010) gegenüber allen anderen untersuchten Schweizer
Populationen hohe FST-Werte auf , was auf grosse genetische Unterschiede hindeutet.
Die Nasenpopulation im Züricher Schanzengraben wurde von der übrigen Metapopulation in Aare und Murg
aufgrund von Flussverbauungen in der Limmat abgetrennt und ging dann höchstwahrscheinlich durch einen
genetischen Flaschenhals (Vonlanthen et al., 2010). Es erfolgte also wohl eine Reduktion der Populations-
grösse auf wenige Tiere, die dann wieder zu Gründertieren der neuen Population wurden. Aufgrund der
dadurch verursachten starken Gendrift unterscheiden sich die Schanzengraben-Nasen genetisch von den üb-
rigen Mittellandpopulationen unterhalb des Rheinfalls (Vonlanthen et al., 2010). So auch von der Population
des Thur-/Murg-Systems, aus welchen in den letzten Jahren Nasen im Linthkanal eingesetzt wurden. Es han-
delt sich bei den Schanzengrabennasen natürlicherweise aber vermutlich nicht um eine völlig eigenständige
Population, sondern um eine durch Flaschenhals und Gendrift entstanden lokale Variante, die im Vergleich
zu den Populationen an Thur und Murg genetisch verarmt ist (Vonlanthen et al., 2010), aber ursprünglich
doch Teil der grossen Metapopulation des Mittellandes war.
Andere Studien haben gezeigt, dass der genetische Austausch zwischen nah gelegenen Nasen-Populationen
hoch ist (Hudson et al., 2014, Wetjen et al., 2020) was zu kaum messbaren genetischen Unterschieden in
doch recht weit verbreiteten aber ursprünglich zusammenhängenden Populationen führt. Dies zumindest,
sofern sie nicht durch Wanderhindernisse über viele Generationen hinweg voneinander getrennt waren. Ein
Homing-Verhalten bei den Laichzügen ist für Nasen bisher nicht bekannt (Vonlanthen et al., 2010). Sie laichen
also nicht wie Forellen und Lachse am Ort, an dem sie selbst geschlüpft sind, sondern wahrscheinlich je nach
Jahr an verschiedenen Orten und gegebenenfalls sogar in unterschiedlichen Flüssen.
Historische Vorkommen der Nase im Zürchseegebiet
Die Nase war in Limmat, Sihl, Zürichsee, Linth und Walensee früher verbreitet und teilweise häufig (Maier et
al., 1995). Im Verlauf des 20. Jahrhunderts wurde die Nase im Zürichsee-Einzugsgebiet immer seltener und
verschwand schliesslich fast vollständig. Als Gründe hierfür nennen Maier et al. (1995) den Kraftwerksbau in
Wettingen im Jahr 1932, durch den die Limmat für die Nase und andere Fische unpassierbar wurde. Das letzte
Nasenlaichgebiet der Limmat erlosch daraufhin im Jahr 1950 (Maier et al. 1995). Neben Kraftwerksbauten
und der damit einhergehenden fehlenden Vernetzung der Habitate, sind weitere Ursachen wohl in der star-
ken Verbauung der Ufer und ebenso in der schlechten Wasserqualität von Zürichsee und Limmat zu suchen.
6Genetische Untersuchungen der Nasen in der Zürichseeregion
Pedroli et al (1991) gaben die Häufigkeit der Nase im Zürichsee mit sehr selten an. Im Walensee war sie
damals aber wohl noch etwas häufiger als im Zürichseegebiet (Pedroli et al 1991).
Um das Jahr 2000 wurden einige Fische in der Zürichseeregion gemeldet (vgl. INFO FAUNA http://le-
pus.unine.ch/carto/70042). Es existieren laut INFO FAUNA (2021) Fundmeldungen aus dem Schanzengraben
bei Zürich (2000), dem Zürichseebecken (2000 u. 2001) und dem Walensee (2001). Diese Informationen sind
allerdings nicht verifizierbar und den Fischereibehörden auch nicht als Beobachtungen bekannt.
Bei der Schanzengrabenpopulation handelte es sich wahrscheinlich um eine Reliktpopulation, wie Vonlan-
then et al. (2010) zeigen konnten. Weitere Fundmeldungen vom Schanzengraben, der Sihl und aus dem
Hauserkanal stammen von Hoppler (2014). Der Hauserkanal kann Fischen theoretisch als Umgehungsgerinne
für das KW Höngg dienen, an dem erst seit kurzer Zeit eine Fischaufstiegshilfe existiert (Bammatter persönli-
che Mitteilung 2021). Hoppler (2014) konnte in der Limmat unterhalb des Kraftwerks Höngg und im Hauser-
kanal die Existenz von Nasenlarven und damit Laichtätigkeit bestätigen. Er vermutete eine Zuwanderung von
laichfähigen Nasen aus der Aare, allerdings ist auch ein Abstieg von Nasen aus der Schanzengrabenpopulation
möglich.
Der Schanzengraben dient neben der Limmat, mindestens seit 1830 als zweiter Abfluss des Zürichsees. Ab
1864 wurde er in die Sihl abgeleitet, in die er auch heute noch bei der Gessner Allee einmündet (Bauer et al.,
1992). Durch den Einbau einer Fischtreppe ins Wehr beim „Flussbad Schanzengraben“ ergab sich somit ein
direkter Wanderkorridor von Limmat über die Sihl in den See.
Aktuelle Durchgängigkeitssituation der Limmat von der Aare bis in den Zürichsee
Kanton Aargau:
Im Kanton Aargau befindet sich am KW Stroppel sowohl eine Fischaufstiegshilfe (Bölli 2014), als auch eine
Fischabstiegseinrichtung (Bölli 2014). Weitere KW in der Limmat sind Gebenstorf, Turgi, Schiffmühle, Kap-
pelerhof, Oederlin, Aue und Wettingen (Kanton Aarau 2018). In der Literatur finden sich aber nicht für alle
KW gesicherte Informationen zur Durchgängigkeit. Für die KW Kappelerhof und Wettingen wurde der Fisch-
aufstieg bereits untersucht und er ist dort zumindest möglich (Kirchhofer, 2011). Für die Kraftwerke
Schiffmühle und Aue wurden ebenfalls FAH gebaut und der Fischaufstieg als möglich beurteilt. Sogar einige
Nasen konnten in den FAH beobachtet werden (Schölzel et al.2020).
Nasenfänge gab es zwischen 2003 und 2007 im Hard (Kirchhofer, 2011) sowie am Stauwehr Kappelerhof
zwischen 2003 und 2006 (Kirchhofer, 2011) und ebenfalls an der Schiffmühle (INFO FAUNA, http://le-
pus.unine.ch/carto/70042 abgerufen am 01.07.2021).
Kanton Zürich:
Laut Bammatter (persönliche Mitteilung 2021) ist nicht klar, ob die Durchgängigkeit für Fische in der Limmat
gewährleistet ist. Ein Durchwandern der Limmat bis in den Zürichsee kann aber auch nicht ausgeschlossen
werden. Es gibt aktuell neue Fischwanderhilfen beim Kraftwerk Dietikon. Beim KW Höngg ist die Situation
weniger gut, da momentan am Kraftwerk selbst keine Fischaufstiegshilfe (FAH) existiert, sondern lediglich
am dortigen Wehr. Ein Aufstieg über den Hauserkanal ist laut Bammatter zwar theoretisch denkbar, aber
sehr unwahrscheinlich. Hoppler (2014) konnte aber bereits Nasen im Hauserkanal beobachten. Die FAH am
Kraftwerk Letten funktioniert laut Bammatter (persönliche Mitteilung 2021) ebenfalls nicht wunschgemäss
7Genetische Untersuchungen der Nasen in der Zürichseeregion
und wird zurzeit verbessert. Beim Platzspitzwehr ist ein Mäanderfischpass in Planung, da die Sihl aber unter-
halb von diesem einmündet, ist die Durchgängigkeit zum Zürichsee immerhin über den Umweg Sihl und
Schanzengraben gegeben. Im Schanzengraben gibt es ebenfalls noch eine weitere FAH Richtung See, sodass
ein Durchgang von der Kantonsgrenze bis in den See theoretisch möglich ist.
Den Autoren ist keine Studie bekannt, die die Durchgängigkeit über die gesamte Strecke der Limmat (von der
Mündung in die Aare bis zum Zürichsee) untersucht hat. Von den Kantonen Aargau und Zürich sind aber in
den letzten Jahren grosse Anstrengungen unternommen worden, um die Strecke für Fische durchgängig und
damit einen Laichzug der Nasen möglich zu machen.
Abbildung 1: Fischgängigkeit der Limmatkraftwerke im Jahr 2011. Quelle Kirchhofer 2011
Fördermassnahmen im Zürichsee-Einzugsgebiet
Ab 2015 wurde nach Annahme der Abstimmung in der Fischereikommission das Artenförderprogramm der
Nasen im Zürichseegebiet für den Zürichsee- Linthkanal und Walensee (FIKO) gestartet. Dazu wurde in der
Murg (TG) Laichfischfang durchgeführt. Nach der Verpaarung der gefangenen Tiere (gemäss Besatzrichtlinien
mit möglichst vielen Weibchen und Männchen (N > 50) und Paarungsvarianten zur Erhaltung einer möglichst
grossen genetischen Variabilität) wurden die Eier im Fischereizentrum Steinach erbrütet. Die geschlüpften
Fische wurden mit Natur- und Kunstfutter aufgezogen und als Pilotprojekt in revitalisierte Bereiche des Linth-
kanals sowie dessen Seitenkanäle eingesetzt (Kugler persönliche Mitteilung 2021). Der Linthkanal verbindet
den Walensee mit dem Zürichsee. Sowohl der Rechte als auch der Linke Linthnebengraben wurden revitali-
siert; sie verlaufen parallel zum Linthkanal und münden unmittelbarer neben diesem in den Zürich-Obersee.
8Genetische Untersuchungen der Nasen in der Zürichseeregion
Abbildung 2. Besatzstelle im Linthkanal und Obersee mit Seedamm. Quelle: Swisstopo
Im oben beschriebenen Pilotprojekt wurden in den Jahren 2015, 2016, 2018, 2019 und 2021 insgesamt
153‘000 Nasensömmerlinge (~ 30 – 50 mm, vgl. Tabelle 1), die von Elterntieren aus der Murg stammen im
Linthkanal und geeigneten Seitengewässern eingesetzt. Frühere Besatzmassnahmen mit Nasen hat es laut
Bammatter (persönliche Mitteilung 2021), im Kanton Zürich vorher nicht gegeben.
Tabelle 1: Übersicht zum Besatzmanagement der Nasen im Linthkanal (Quelle: Kugler persönliche Mitteilung 2021)
Besatzgewässer Herkunft Datum Anzahl
Linthkanal / linker Linthhintergraben Murg/Thur 25.09.2015 25'000 Fische
Linthkanal Murg/Thur 26.08.2016 20'000 Fische
Linthkanal+ rechter Linthnebengraben Murg/Thur 29.08.2018 30'000 Fische
Linthkanal+ rechter Linthnebengraben Murg/Thur 06.08.2019 28'000 Fische
Linthkanal +rechter Linthnebengraben Murg/thur 15.07.2021 50'000 Fische
Total Murg/Thur 2015-2021 153'000 Fische
Fragestellungen
Im Frühjahr 2021 wurden im Zürichsee häufiger Nasen von Netzfischern gefangen. Dieses plötzliche und ge-
häufte Auftreten wurde nicht erwartet und hat auch die Fischer und die Fischereibehörde überrascht. Es
wurde sogar ein Fernsehbeitrag zu diesem Thema bei TeleZüri veröffentlich. Die kantonalen Fischereibehör-
den SG und ZH haben deshalb von 19 der 59 durch Netzfischer gefangenen Nasen DNA Proben entnommen.
Mit der vorliegenden Studie soll genetisch überprüft werden, ob die gefangenen Nasen aus dem Besatz der
letzten Jahre im Linthkanal stammen oder ob sie allenfalls natürlicherweise aus dem Schanzengraben oder
von einer weiter entfernten Population eingewandert sind, bzw. einer solchen Population genetisch näher
stehen. Ausserdem soll untersucht werden, ob sich bei den Fängen aus dem Zürichsee Aussagen hinsichtlich
genetischer Vielfalt machen lassen. Dies sind wichtige Daten für die zukünftige Bewirtschaftung der Nasen in
der Region.
9Genetische Untersuchungen der Nasen in der Zürichseeregion
Material und Methoden
Nasenfänge im Zürichsee 2021
Zwischen dem 04.01.2021 und 04.03.2021 wurden im Zürichsee insgesamt 59 Nasen von Berufsfischern ge-
fangen. Von 19 dieser Fische wurden Gewebeproben entnommen und für eine genetische Analyse zur Ver-
fügung gestellt. 17 konnten schliesslich genetisch ausgewertet werden. Die Fangstandorte und Daten sind
nach Angabe der Fischer in Tabelle 2 aufgeführt.
Tabelle 2: Daten zu Gewicht, Länge sowie Datum und Ort der Zürichseefische
Probe Nr. Labor Nr. Fangort Datum Länge (cm) Gewicht in g Alter Geschlecht
Nase-FIKO-30 L06801 Strandbad Meilen 24.02.2021 45 1100 5+ M
Nase-FIKO-31 L06802 Herrliberg 26.02.2021 44 1000 W
Nase-FIKO-32 L06803 Hurden / Obersee 26.01.2021 42 6+
Nase-FIKO-33 L06804 Feldbach 27.01.2021 43 4+
Nase-FIKO-34 L06805 Feldbach 27.01.2021 44 4+
Nase-FIKO-35 L06806 Altendorf / Obersee 30.01.2021 40 3+
Nase-FIKO-36 L06807 Altendorf / Obersee 30.01.2021 46 5+
Nase-FIKO-37 L06808 Lachen / Obersee 15.02.2021 42 5+
Nase-FIKO-38 L06809 Feldbach 18.02.2021 43 6+
Nase-FIKO-39 L06810 Nuolen / Obersee 04.03.2021 48 5+
Nase-FIKO-40 L06811 Nuolen / Obersee 04.03.2021 42 5+
Nase-FIKO-41 L06812 Seebecken 11.03.2021 44 1020 M
Nase-FIKO-42 L06813 Seebecken 22.01.2021 44 1000
Nase-FIKO-43 L06814 Seebecken 22.01.2021 43 1060
Nase-FIKO-44 L06815 Seebecken 22.01.2021 49 1460
Nase-FIKO-45 L06816 Seebecken 22.01.2021 45 1060
Nase-FIKO-46 L06817 Seebecken 29.01.2021 41 900
Nase-FIKO-47 L06818 Seebecken 29.01.2021 41 920
Nase-FIKO-48 L06801 Strandbad Meilen 24.02.2021 45 1100
Bemerkenswert sind das Gewicht und die Länge der untersuchten Fische. Das Gewicht betrug bei allen Fi-
schen von denen Proben genommen wurden, ca. 1 kg (bei einem sogar 1460g). Ebenso waren alle untersuch-
ten Fische mindestens 40cm lang. Daher kann davon ausgegangen werden, dass es sich um ausgewachsene,
geschlechtsreife Nasen handelt (vgl. Abschnitt Biologie der Nase) (Kugler, persönliche Mitteilung 2021). Al-
tersbestimmungen zeigen, dass die Nasen zwischen 3+ und 6+, also vier bis sieben Jahre alt waren (Altersbe-
stimmungen M. Kugler + P. Vonlanthen). Zum Geschlecht der einzelnen Fische liegen nur Informationen von
drei Fischen vor, bei diesen wurde auch die Gonadenentwicklung angeschaut - alle drei waren fortpflanzungs-
fähig. Zu Grösse und Gewicht der 41 weiteren gefangen Nasen liegen keine weiteren Daten vor.
10Genetische Untersuchungen der Nasen in der Zürichseeregion
Vorgehensweise zum Vergleich der Populationen
Zusätzlich zu den Proben aus dem Zürichsee, wurden folgende Proben als Referenzen für die Analysen mit-
einbezogen: Nasen-Besatz Linthkanal 2015-2019, Murg, Schanzengraben, Wiese, Birs und Binnenkanäle des
Alpenrheins. Mit Ausnahme der Schanzengrabenpopulation und der Binnenkanäle gehören alle anderen Re-
ferenzproben zur Mittellandmetapopulation. In Abbildung sind die Probenahmestandorte der in dieser Stu-
die untersuchten Nasenpopulationen dargestellt. Genaue Koordinaten finden sich in Tabelle 3.
Tabelle 0: Übersicht zu den Probenahmen der vorliegenden Studie.
Population Herkunft / Fangdatum Koordinaten LV95 Probenanzahl
Zürichsee (gesamt) Zürichsee / Fang Frühjahr 2021 Verschiedene Standorte 17
Teilpopulation (grosses Seebecken) Zürichsee (grosses Seebecken)/ 04/2021 Verschiedene Standorte 12
Teilpopulation (Obersee) Zürichsee (Obersee)/ 04/2021 Verschiedene Standorte 5
Murg (Elterntiere) Murg / 04.04.2019 2'709'603.2, 1'269'495.9 18
2'719'227.0, 1'225'692.3
Nasen-Besatz Linthkanal 2019 Besatz am 06.08.2019 Eltern aus Murg 21
(Besatzkoordinaten)
2'719'227.0, 1'225'692.3
Nasen-Besatz Linthkanal 2015 Besatz am 25.09.2015 Eltern aus Murg 21
(Besatzkoordinaten)
2'719'227.0, 1'225'692.3
Nasen-Besatz Linthkanal 2016 Besatz am 26.08.2016 Eltern aus Murg 21
(Besatzkoordinaten)
CH-Murg (Mu20) Murg (keine Daten zum Zeitpunkt ) 2'709'622.8, 1'269'429.6 18
Murg, / 19.04.2008 vgl. Vonlanthen et al.
CH-Murg (Mu08) 2'709'603.2, 1'269'495.9 44
(2010)
Zürich, Schanzengraben / 09.04.2008 vgl.
Schanzengraben 2'682'852.5, 1'247'731.2 21
Vonlanthen et al. (2010)
Basel / 21.04.2008 vgl. Vonlanthen et al.
Wiese 2'611'550.0, 1'270'103.8 31
(2010)
Nahe Basel /06.05.2008 vgl. Vonlanthen et
Birs 2'613'520.0, 1'267'016.2 29
al. (2010)
CH-Binnenkanäle (BiC) Rietach (St. Galler Rhein) / 28.05.2008 2'764'417.2, 1'251'294.4 3
“ Mühlbach (St. Galler Rhein) / 28.05.2008 2'753'687.3, 1'232'990.4 2
“ Simmi 1 (St. Galler Rhein) / 28.05.2008 2'755'037.4, 1'231'302.7 14
“ Simmi 2 (St. Galler Rhein) / 28.05.2008 2'754'602.2, 1'231'228.1 6
“ Simmi 3 (St. Galler Rhein) / 28.05.2008 2'754'289.8, 1'231'004.9 3
Total 267
Die meisten DNA-Proben sind 2008 von den Wildpopulationen der Murg gesammelt worden. Hier sind, auf-
grund der hohen Stichprobengrösse (44 Fische), sehr robuste Resultate zu erwarten. Zudem wurden auch
noch weitere Referenzproben aus der Murg von 2020 (n=18) analysiert (vgl. Vonlanthen et al., 2010) die
sowohl einzeln, als auch zusammen mit der grösseren Stichprobe in die Analyse einbezogen wurden. Die
Population der Schweizer Binnenkanäle dient in dieser Studie als Kontrollgruppe. Sie gehört der Bodensee-
Alpenrhein Metapopulation an und wurde bereits in vorhergehenden Studien genetisch charakterisiert (Kre-
ienbühl & Vonlanthen 2021). Sie sollte sich daher von allen anderen in dieser Studie untersuchten Nasenpo-
pulationen unterscheiden.
11Genetische Untersuchungen der Nasen in der Zürichseeregion
Abbildung 4: Übersicht zu den verschiedenen Probeentnahmestandorten der Nasen im Einzugsgebiet des Rheins.
Laborarbeiten
Die DNS (Desoxyribonukleinsäure) jedes Individuums wurde mithilfe der Chelex-Methode extrahiert (de Lam-
ballerie et al. 1992). Dazu wurden die Gewebestücke mit 0.15 ml Chelex-Lösung (5% Lösung, Hersteller: Bio-
rad), 0.01 ml TE-Buffer und 0.005 ml Proteinase K während 2 Stunden bei 55 Grad inkubiert, um die DNS aus
den Zellen herauszulösen. Bei 17 der 18 genetisch untersuchten Nasen wurde die DNA erfolgreich isoliert
und die PCR fehlerfrei durchgeführt. Bei einer Probe funktionierte dieser Prozess nicht. Insgesamt wurden 8
Mikrosatelliten-Marker analysiert. Es handelt sich dabei um die gleichen Marker, die in den Studien von Von-
lanthen et al., (2010) und Wetjen et al., (2020) verwendet wurden. Dies sind LC27, LSOU21, LSOU05,
SARN7G5, SARN7F8, SARN7K4, LSOU08, LC290 sowie SARN2F11B. Ein weiterer Marker - LC290, wurde, wie
schon in früheren Studien, aufgrund seiner schlechten Datenqualität von den Auswertungen ausgeschlossen.
Die Primersequenzen für die Marker wurden ursprünglich aus folgenden Publikationen entnommen
(Mesquita et al., 2003; Vyskočilová et al., 2007; Muenzel et al., 2008). Die PCR-Amplifikation der Mikrosatel-
liten erfolgte nach folgendem Protokoll:
• 1. Zyklus bei 95°C für 12 min
• 35 Zyklen bei 95°C für 20 s, 60°C für 50 s und 72°C für 120 s
• letzter Zyklus bei 72°C für 5 min
Die dabei entstandenen Produkte wurden in einem DNA-Sequenziergerät (Applied Biosystems 3730XL DNA
Analyzer) aufgetrennt und mit Genemarker (Hulce et al. 2011) manuell ausgewertet. Die dadurch erhaltenen
Informationen über die Fragmentlängen wurden anschliessend für die statistischen Auswertungen und Tests
weiterverwendet.
Statistische Auswertungen und Tests
Für statistische Auswertungen und Tests wurden folgende populationsgenetische Programme verwendet:
FSTAT (Goudet, 2001), GeneClass2 (Piry et al., 2004), Arlequin (Excoffier & Lischer, 2010), Structure (Pritchard
et al., 2000). Falls sinnvoll und notwendig wird die Funktionsweise einer Analyse oder eines Tests direkt in
den Resultaten besprochen.
12Genetische Untersuchungen der Nasen in der Zürichseeregion
Ergebnisse
Die Nasen, welche im Zürichsee gefangen wurden, können grundsätzlich von drei Quellen abstammen. Einer-
seits ist es möglich, dass im Linthkanal eingesetzte Nasen in den See gewandert sind. Andererseits könnten
auch Nasen aus dem Limmat - Schanzengraben System in den Zürichsee aufgestiegen sein. Da die beiden
Populationen genetisch verschieden sind (Vonlanthen 2010) sollte es daher möglich sein, genetisch zu erken-
nen, ob die Zürichseenasen der einen oder der anderen Population angehören. Als dritte Möglichkeit wäre
der Aufstieg von Nasen aus der Aare über Limmat und Schanzengraben zu nennen. Die gefangenen Fische
können nicht aus einer bereits erfolgten Einkreuzung von Besatzfischen in die lokale Population hervorge-
gangen sein, da der Besatz erst im Jahr 2015 begann und Nasen erst nach 4-7 Jahren Geschlechtsreif werden.
Die gefangenen Tiere im See waren aber alle gross und dürften gemäss Altersbestimmungen ca. 5 Jahre alt
sein (Kugler pers. Mitteilung 2021). Sie gehören also entweder der Besatzpopulation von 2015/16 oder der
Schanzengrabenpopulation an, bzw. sind von ausserhalb eingewandert. Die genetischen Unterschiede zwi-
schen Besatznasen und denen aus der Mittellandmetapopulation (Murg /Aare etc.) sind zu gering, als dass
diese Populationen mit den in dieser Studie verwendeten Marker unterschieden werden könnten. Daher
kann ein natürlicher Aufstieg der Nasen aus der Mittellandpopulation via Limmat nicht ganz ausgeschlossen
werden (vlg. Kapitel Durchgängigkeit Limmat).
Basisanalysen
Anhand der Basisanalysen wird die Qualität der Daten und somit der Untersuchung beurteilt (vgl. Tabelle 0).
Als erstes wurde die genetische Variabilität der einzelnen Populationen bestimmt. Das Resultat sind zwei
Werte - die durchschnittliche Anzahl beobachteter Allele pro Lokus und Population (AN) sowie deren für die
Stichprobengrösse korrigierter Wert (AR). Je höher die beiden Werte, desto höher ist die genetische Vielfalt.
In der Schanzengrabenpopulation fällt der Wert, der für die Allele pro Lokus mit AR = 3.305 wie erwartet
niedriger aus, als bei den anderen untersuchten Populationen. Nur bei den im Zürichsee gefangenen Fischen
ist dieser Wert mit AR = 3.109 noch niedriger. Die Berechnungen wurden mit dem Programm FSTAT durchge-
führt (Goudet, 2001).
Tabelle 0: Übersicht zu den Basisanalysen und ihren Resultaten (n = Anzahl Proben; AN = durchschnittlich beobachtete Anzahl Allele
pro Lokus; AR = korrigierter AN; HO = beobachtete Heterozygotie; HE = erwartete Heterozygotie; p-WertH = Signifikanz Abweichung H
vom Hardy-Weinberg-Gleichgewicht; FIS = Inzuchtkoeffizient; p-Wert = Signifikanz des Inzuchtkoeffizienten; NLD = Anzahl Loci mit phy-
sischer Assoziation).
Standort N AN AR FIS p-Wert H0 HE p-Wert NLD pGenetische Untersuchungen der Nasen in der Zürichseeregion
oder sozial eingegrenzten Populationen, die sich nur oder vorwiegend untereinander paaren, vermehrt zu
Erbkrankheiten und sog. Inzuchtdepression kommen. Keine der untersuchten Populationen wies jedoch ei-
nen signifikanten Inzuchtkoeffizient auf.
Anschliessend wurden die Werte für die erwartete Heterozygotie (HE) sowie die beobachtete Heterozygotie
(HO) berechnet. Diese Analysen wurde mit dem Programm Arlequin durchgeführt (Excoffier & Lischer, 2010).
Die beiden Werte sind ein Mass dafür, wie häufig an einem Lokus zwei unterschiedliche Allele (Heterozygotie)
beobachtet werden können. Dabei wird auch der zugehörige p-Wert angegeben. Dieser zeigt auf, ob sich die
beobachtete Heterozygotie von der erwarteten Heterozygotie signifikant unterscheidet, wobei angenommen
wird, dass sich die Populationen im Hardy-Weinberg-Gleichgewicht befinden (zufällige Verpaarung von Indi-
viduen, keine Zu- oder Abwanderung). Die p-Werte für HE und HO liegen für alle Populationen zwischen 0.364
und 0.707. Sie liegen damit in einem mit anderen Studien vergleichbaren Bereich (Vonlanthen et al., 2010,
Wetjen et al., 2020). Keine der Populationen weicht signifikant vom Hardy-Weinberg-Gleichgewicht ab.
Als letztes wurde überprüft, wie viele der einzelnen Mikrosatelliten-Loci physisch auf dem Genom assoziiert
sind (LD = engl. linkage disequillibrium). Die Berechnungen wurden mit dem Programm Arlequin gemacht
(Excoffier & Lischer, 2010). Eine stark gehäufte Assoziation zwischen unterschiedlichen Loci konnte nur beim
Nasenbesatz im Linthkanal 2019 nachgewiesen werden. Da die gefundenen Assoziationen verglichen zwi-
schen den Populationen an unterschiedlichen Loci vorgefunden wurden, kann davon ausgegangen werden,
dass die verschiedenen Loci unabhängig voneinander vererbt werden (keine physische Assoziation auf den
Chromosomen). Somit sind weiterführende populationsgenetische Analysen zulässig.
Die Basisanalysen bestätigen die Ergebnisse aus früheren Studien (vgl. Vonlanthen et al., 2010, Kreienbühl &
Vonlanthen 2021), dass alle untersuchten lokalen Nasenpopulationen über eine ausreichende genetische
Vielfalt verfügen, auch wenn diese im Schanzengraben aufgrund dem fehlenden genetischen Austausch mit
anderen Populationen geringer ist. Auch bei den Zürichseenasen ist eine geringere genetische Vielfalt zu be-
obachten. Die Inzuchtkoeffizienten sowie die beobachtete Heterozygotie liegen in einem Bereich, der auch
bei anderen Wildpopulationen beobachtet werden konnte (Vonlanthen et al., 2010, Wetjen et al., 2020). Ob
die genetische Verarmung in Schanzengraben und Zürichsee ein Problem für die Fitness der Fische darstellt
kann aus den Ergebnissen nicht abgeleitet werden. Die geringe genetische Vielfalt der Zürichseenasen ist
noch kein Beweis dafür, dass die Fische der Schanzengrabenpopulation angehören. Dafür sind weitere Ana-
lysen notwendig.
Auch wenn die Populationen der freilebenden Nasenpopulationen heute klein sind, scheinen sie doch eine
ausreichend grosse Vielfalt bewahrt zu haben, die ein erfolgreiches Anwachsen der Populationen ermögli-
chen, wenn sich die Lebensumstände für die Fische in Zukunft wieder verbessern sollten.
Vergleiche zwischen den Populationen
Die genetische Differenzierung zwischen zwei Populationen wird mit dem sogenannten FST-Wert (engl. fixa-
tion index) beschrieben. Der FST-Wert basiert auf der Varianz der Allelfrequenzen von zwei unterschiedlichen
Populationen an einem oder mehreren Loci. Je höher der FST-Wert ist, desto grösser ist die genetische Eigen-
ständigkeit der beiden verglichenen Populationen. Ist der FST-Wert nahe 0, so sind die Populationen identisch.
Eine schwache genetische Differenzierung zwischen Populationen mit viel Genfluss liegt vor, wenn der FST
zwischen 0 und 0.05 liegt (Vonlanthen & Hefti, 2016). Die beiden Populationen sind also genetisch sehr ähn-
lich. Eine mittlere genetische Differenzierung liegt ab einem FST von 0.05 vor. Ab einem FST von 0.15 kann von
einer starken genetischen Differenzierung mit sehr geringem Genfluss gesprochen werden. In diesem Fall
unterscheiden sich die Populationen also genetisch stark voneinander. Die Aussagekraft von FST-Werten ist
14Genetische Untersuchungen der Nasen in der Zürichseeregion
abhängig von der Stichprobenanzahl. Es muss daher eingeschränkt werden, dass FST-Werte von Populationen
mit geringer Stichprobengrösse (n0.05). Im Bereich über der Diagonale befindet sich
an der entsprechenden Stelle ein (-). In einem solchen Fall sind die beiden verglichenen Populationen also eng verwandt. Signifikante
FST-Werte sind durch ein + dargestellt. Die Populationen sind genetisch also signifikant unterschiedlich.
Standort N Nr. 1 2 3 4 5 6 7 8 9 10 11 12 13 14
Zürichsee gesamt 17 1 0 - - + + + + + + + + + + +
Zuerichsee Seebecken 12 2 -0.028 0 - + + + + + + + + + + +
Zuerichsee Obersee 5 3 -0.028 0.044 0 - + + + + + + + + + +
Elterntiere Murg 18 4 0.041 0.058 0.021 0 - + - - - + - - + -
Nasen Besatz Linth 2019 15 5 0.042 0.057 0.054 0.012 0 + + - + + + + + +
Nasen Besatz Linth 2015 20 6 0.07 0.084 0.068 0.031 0.088 0 + + + + + + + +
Nasen Besatz Linth 2016 21 7 0.059 0.071 0.053 0 0.012 0.082 0 - + + + + + +
Murg20 16 8 0.03 0.033 0.044 -0.002 0.003 0.053 0.005 0 - + - - + -
Murg08 42 9 0.067 0.071 0.075 0.008 0.025 0.056 0.035 0.005 0 + - - + -
Schanzengraben 21 10 0.044 0.03 0.117 0.112 0.073 0.129 0.113 0.072 0.118 0 + + + +
Wiese 31 11 0.054 0.063 0.051 -0.002 0.024 0.048 0.019 0.003 -0.002 0.121 0 - + -
Birs 29 12 0.053 0.068 0.035 -0.009 0.019 0.024 0.029 0.011 0.006 0.117 0.001 0 + -
CH-Binnenkanal 25 13 0.091 0.09 0.097 0.062 0.073 0.067 0.106 0.069 0.072 0.141 0.075 0.064 0 +
Murg08 + Murg20 58 14 0.055 0.06 0.065 0.004 0.017 0.053 0.026 -0.006 -0.008 0.102 -0.002 0.006 0.072 0
Folgende weitere Beobachtungen können bei den genetischen Differenzierungen (mittels FST) festgehalten
werden:
• Die Berechnung der FST-Werte zeigt, dass sich die meisten Populationen mit ausreichender Stichpro-
bengrösse signifikant von den anderen untersuchten Populationen unterscheiden (Tabelle 5)
• Innerhalb desselben Gewässers konnte kein genetischer Unterschied zwischen der Population der
Murg die 2009 und 2021 untersucht wurden, gefunden werden (FST = 0.005, Unterschied nicht signi-
fikant). Da es sich jeweils um die gleiche Population handelt, ist ein solches Ergebnis zu erwarten. Es
zeigt auch, dass die Daten von 2009 mit denen von 2021 vergleichbar sind und dass im Labor bei der
Zuweisung der Allele die Interkalibrierung funktioniert hat.
• Die genetischen Unterschiede zwischen den Mittellandnasenpopulationen (Wiese, Birs, Murg) sind
moderat. Die genetischen Unterschiede der Mittelandmetapopulation zu den Alpenrheinnasen (CH-
Binnenkanäle) lassen sich allerdings klar aufzeigen. Dies obwohl bis vor einigen Jahren noch Besatz-
massnahmen mit Murgfischen in den Binnenkanälen durchgeführt wurden. Genetische Spuren die-
ser Besatzmassnahmen sind, mit den in dieser Studie verwendeten Methoden, heute nicht mehr
nachweisbar.
• Interessant ist der geringe genetische Unterschied (nicht signifikanter FST-Wert) der Proben aus dem
Obersee mit den Elterntieren des Murgbesatzes von 2019. Allerdings ist die Stichprobengrösse aus
dem Obersee mit 5 Fischen sehr klein, genauere Rückschlüssen können daher nur aus individuellen
Analysen gezogen werden (siehe unten).
15Genetische Untersuchungen der Nasen in der Zürichseeregion
Herkunft der Zürichseenasen
Um diese Frage der Herkunft der Zürichseenasen zu klären, wurde vorerst mit dem Programm Structure (Prit-
chard et al., 2000) eine mögliche Populationszuteilung errechnet. Mit diesem Ansatz kann untersucht wer-
den, ob und wie die einzelnen Individuen in unterschiedliche, genetisch eigenständige Populationen einge-
teilt werden können. Die Software ermöglicht darüber hinaus zu erkennen, ob einzelne Individuen, die am
gleichen Standort gefangen wurden, allenfalls unterschiedlichen Populationen zugewiesen werden können.
Mit der Software werden Individuen anhand des mit Mikrosatelliten erhobenen genetischen Fingerabdrucks
einer vordefinierten Anzahl Populationen (k) mit einer gewissen Wahrscheinlichkeit zugeordnet. Die Anzahl
Populationen, die untersucht wird, muss vom Nutzer bei jeder Simulation vorbestimmt werden. Insgesamt
wurden bei den vorliegenden Auswertungen jeweils zehn Simulationen pro k-Wert berechnet. Es wurde mit
k= 2 und k=3 gerechnet (burnin=30'000, MCMC =300'000). Anhand der Ausgangslage wurde entschieden,
dass k=2 (Mittellandmetapopulation und Schanzengraben) die schlüssigsten Ergebnisse liefert. Darauf auf-
bauend wurden die Ergebnisse in Abbildung 5 dargestellt.
Abbildung 5: Structure-Plot, der die unterschiedlichen Nasenpopulationen aus dem Schanzengraben (Sch08) mit den Besatzfischen
aus der Murg (Mu19) und Proben aus dem Zürichsee vergleicht (k=2, Burnin=30'000, MCMC=300'000; von links nach rechts: Elterntiere
Murg (Mu19) in grün; Zürichsee (ZS21) und Schanzengraben (Sch08) in blau. Dazu sind die Bezeichnungen für die jeweiligen Einzelfi-
sche angegeben. Je höher der Farbanteil in einer Kolonne umso höher ist der Anteil der Gene einer bestimmten Population.
Es ist ersichtlich, dass sich die Schanzengrabenpopulation von den Eltern des Murgbesatzes -also genetisch
gesehen Vertretern der Mittellandmetapopulation - deutlich unterscheidet. Bei den gefangenen Zürichseefi-
schen gibt es einige, die sich sehr gut einer der beiden anderen Populationen zuordnen lassen, allerdings sind
auch Nasen vorhanden, die genetisch in der Mitte stehen (mögliche Hybride, bzw. genetisch intermediäre
Individuen). Es scheint sich bei den gefangenen Zürichseenasen also um Individuen zu handeln die mehreren
Ursprungspopulationen zugewiesen werden können.
Eine genauere Zuordnung eines Individuums zu einer bestimmten Population ist mit dem Programm Genec-
lass2 (Piry et al., 2004) möglich. Dabei werden Individuen zwei Referenzpopulationen zugewiesen. Da auf-
grund der „Structuranalysen“ und der Ausgangslage bekannt war, dass nur zwei (mittels Mikrosatelliten) ge-
netisch unterscheidbare Populationen vorliegen, wurden die Fische aus dem Zürichsee nur mit der Schan-
zengraben- und der Mittelandmetapopulation (Murg 2020) verglichen. Die Übereinstimmung ist in Tabelle 6
gezeigt und wird in Prozentwerten angegeben (100% volle Übereinstimmung; 0% keine Übereinstimmung).
16Genetische Untersuchungen der Nasen in der Zürichseeregion
Bei dieser Analyse werden 12 Individuen dem Schanzengraben zugewiesen und 3 Individuen der Murg-Popu-
lation. Drei Individuen können nicht eindeutig zugewiesen werden. Das Resultat bleibt gleich, wenn für die
Murgreferenz Besatzfische als Referenz verwendet werden.
Tabelle 6: Einordung der im Zürichsee gefangen Fische zu den Populationen Schanzengraben oder Elterntiere Murg. Die Einordnung
erfolgte mit dem Programm Geneclass2. Individuen, die eindeutig der Mittellandmetapopulation (Eltern Murg 2019) zugeordnet wer-
den konnten sind grün, solche die der Schanzengrabenpopulation zugeordnet wurden sind blau dargestellt. Bei den beiden orange
dargestellten Fischen (FIKO-30 + FIKO 31) kann keine genaue Aussage getroffen werden. Diese beiden Fische lassen sich also weder
der einen noch der anderen Population zuordnen. Die Übereinstimmung wird in Prozentwerten angegeben (score in %). Dabei gilt
100% volle Übereinstimmung; 0% keine Übereinstimmung.
Diskussion
12 der 17 (ca. 70%) untersuchten Zürichseenasen (beide Seebecken zusammen) lassen sich anhand der Er-
gebnisse der Geneclassanalysen der Schanzengrabenpopulation zuordnen und stammen demnach aus einer
natürlichen Besiedlung des Sees mit Nasen aus der Schanzengrabenpopulation.
Es finden sich auch 3 Fische (FIKO 32, 36 und 41), also 18% die der Mittellandmetapopulation (Murg) zuge-
ordnet werden. Zwei dieser drei wurden im Obersee gefangen. Der Anteil von Nasen aus der Mittellandpo-
pulation ist demnach im Obersee (40%) höher als im Untersee (8.3%). Wir vermuten daher, dass es sich bei
diesen Nasen um Besatzfische aus 2015 oder 2016 handelt. Die Herkunft aus dem Besatz ist für diese beiden
Fische wahrscheinlicher, da sonst statistisch gesehen auch mehr Murgnasen im grossen Seebecken auftreten
müssten.
Bei Nase FIKO 41 aus dem Seebecken, die sich nur mit einer Wahrscheinlichkeit von 71% der Murgpopulation
zuordnen lässt (kein hoher Wert), handelt es sich möglicherweise nicht um eine Nase aus dem Besatz. Glei-
ches gilt für FIKO 30 und 31, die mit derselben Wahrscheinlichkeit dem Schanzengraben und der Mittelland-
population (Murgpopulation) zugewiesen werden. Wieso diese Fische genetisch intermediär sind, kann nicht
geklärt werden. Es müsste überprüft werden, ob in der unteren Limmat allenfalls Nasen leben, die genetisch
intermediär zwischen der Mittellandpopulation und dem Schanzengraben liegen und die den Zürichsee eben-
falls natürlich besiedelt haben könnten. Allenfalls wäre auch zu prüfen, ob sich die Schanzengrabenpopula-
tion seit 2008 genetisch verändert hat. Die Tatsache, dass 2008 eine Nase gefunden wurde, die genetisch zur
17Genetische Untersuchungen der Nasen in der Zürichseeregion
Mittellandpopulation gerechnet wird, lässt vermuten, dass möglicherweise eine Zuwanderung dieser Fische
aus der Limmat möglich sein konnte.
Anhand der vorliegenden Ergebnisse ist insgesamt klar, dass der überwiegende Teil der im Untersee gefan-
genen Nasen natürlich in den See eingewandert ist. In diesem Seeteil wurden von den Berufsfischern auch
am meisten Nasen gefangen. Im Obersee ist der Anteil wahrscheinlich von Besatz stammenden Nasen deut-
lich höher (40%), die Berufsfischer fingen insgesamt im Obersee aber deutlich weniger Nasen.
Es kann also der Schluss gezogen werden, dass insgesamt ein Grossteil der Nasen von natürlichen Zuwande-
rungen abstammen und ein kleiner Teil, der insbesondere im Obersee zu finden ist, sehr wahrscheinlich von
den Besatzmassnahmen abstammt. Zudem lassen die Ergebnisse vermuten, dass auch aus dem Limmatsys-
tem Nasen in den Zürichsee aufgestiegen sein könnten.
Empfehlungen
Die Untersuchung hat gezeigt, dass sowohl natürliche Zuwanderungen als auch Besatzmassnahmen zur Wie-
derbesiedlung des Zürichsees durch die Nase geführt haben. Wiederansiedlungsmassnahmen sind grundsätz-
lich nur so lange notwendig, bis sich eine lokale sich selbst reproduzierende Population etabliert hat. Vor
diesem Hintergrund werden für die zukünftige Bewirtschaftung der Nasen im Zürichsee-Einzugsgebiet fol-
gende Empfehlungen formuliert:
• Es sollte überprüft werden, ob sich Nasen in potenziellen Zuflüssen des Zürichsees fortpflanzen (Be-
obachtungen während der Laichzeit, allenfalls EDNA Wasseranalysen).
• Im Herbst sollte in Gewässern mit und ohne Besatz überprüft werden, ob Jungnasen in den Gewäs-
sern gefunden werden. Dazu eignen sich z.B. Punktbefischungen von geeigneten Nasenhabitaten.
• Wenn die Elterntiere der Fischzucht genetisch beprobt werden, dann könnten in Zukunft Besatzfi-
sche mittels Elternanalyse identifiziert werden.
• Mittels einer EDNA Untersuchung könnte überprüft werden, in welchen Zuflüssen des Zürichsees
heute Nasen vorkommen.
18Genetische Untersuchungen der Nasen in der Zürichseeregion
Literaturverzeichnis
Bammatter, L.; Adjunkt Fischerei, Kanton Zürich, Amt für Landschaft und Natur (Fischerei- und Jagdverwal-
tung)
Bauer C., Rebsamen H., Capol J., (1992): Zürich. In: INSA: Inventar der neueren Schweizer Architektur, 1850-
1920: Städte. Orell Füssli, Abschnitt Bahnhofbrücke, S. 305, https://doi.org/10.5169/SEALS-10931
https://www.e-periodica.ch/digbib/view?pid=ins-001:1992:10::555#198 abgerufen am 01.07.2021
Bölli M. (2014). Wiederherstellung der Fischgängigkeit beim KW Stroppel. Das Kleinkraftwerk Nr. 83 1; ISKB
https://swissmallhydro.ch/wp-content/uploads/2015/12/Das-Kleinkraftwerk_-Nr.-83.compressed.pdf abge-
rufen am 01.07.2021
Bonell, F. W. (2009). Frühjahrsmonitoring an der Dornbirner Ach mit dem Schwerpunkt Nase (Chondrostoma
nasus). University Innsbruck.
Dedual, M. (1990). Biologie et problèmes de dynamique de population du nase (Chondrostoma nasus nasus)
dans la Petite-Sarine. Universität Freiburg.
De Lamballerie, X., C. Zandotti, C. Vignoli, C. Bollet, und P. De Micco, (1992) A one-step microbial DNA extrac-
tion method using “Chelex 100” suitable for gene amplification. Research in microbiology. 143(8): 785-790.
Dönni, W., & Spalinger, L. (2017). Erhaltung und Förderung der Wanderfische in der Schweiz – Zielarten, Ein-
zugsgebiete, Aufgaben.
Excoffier, L., & Lischer, H. E. L. (2010) Arlequin suite ver 3.5: A new series of programs to perform population
genetics analyses under Linux and Windows. Molecular Ecology Resources, 10, 564–567.
Fielenbach, J. (1996) Zeitliche und räumliche Verteilungsmuster der Nase Chondrostoma nasus (L.) in der
Sieg. Diplomarbeit Friedrich Wilhelms Universität Bonn.
Freyhof, J. (1997) Age-related longitudinal distribution of nase, Chondrostoma nasus in the river Sieg,
Germany. Folia Zool. 46 (Suppl. 1): 89–96.
Freyhof, J. (1998) Strukturierende Faktoren für die Fischgemeinschaft der Sieg. Dissertation Friedrich Wil-
helms Universität Bonn. Cuvillier Verlag.
Goudet, J. (2001). FSTAT, a program to estimate and test gene diversities and fixation indices (version 2.9.3).
Hoppler, L. (2014). Monitoringkonzept für die Nase (Chondrostoma nasus) in der Limmat, Sihl und Schan-
zengraben. Zürcher Hochschule für angewandte Wissenschaften.
Huber, M., & Kirchhofer, A. (1997). Habitat use of radiotagged adult nase (Chondrostoma nasus) in a regu-
lated river. Folia Zoologica, 46, 67–77.
Hudson, A. G., Vonlanthen, P., & Seehausen, O. (2014). Population structure, inbreeding and local adapta-
tion within an endangered riverine specialist: The nase (Chondrostoma nasus). Conservation Genetics, 15,
933–951.
19Genetische Untersuchungen der Nasen in der Zürichseeregion
Hulce, D., X. Li, T. Snyder-Leiby, und C.J. Liu, (2011) GeneMarker genotyping software: tools to increase the
statistical power of DNA fragment analysis. Journal of biomolecular techniques: JBT, 22(Suppl): S35.
INFO FAUNA, CSCF, Datenbank, Neuenburg; http://lepus.unine.ch/carto/70042 abgerufen am 1.7.2021
Kanton Aargau; Departement Bau, Verkehr und Umwelt; Abteilung Landschaft und Gewässer. Wasserkraft-
werke im Kanton Aargau https://www.ag.ch/media/kanton_aargau/bvu/dokumente_2/umwelt__na-
tur___landschaft/naturschutz_1/2018_07_01_GN_Ansprechpersonen_Flusskraftwerke.pdf abgerufen am
01.07.2021
Kirchhofer A. (2011). Grenzenlose Mobilität – auch für die Fische der Limmat. Umwelt Aargau, Nr 53.
Kreienbühl & Vonlanthen (2021). Genetische Untersuchungen der Nasen der Bodenseeregion. Aquabios
GmbH. Auftraggeber: Internationale Bevollmächtigtenkonferenz für die Bodenseefischerei (IBKF). In press.
Kugler, M. Kanton St. Gallen; Amt für Natur, Jagd und Fischerei; Abteilung Fischerei;
Maier, K. J., Zeh, M., Ortlepp, J., Zbinden, S., & Hefti, D. (1995). Verbreitung und Fortpflanzung der in der
Schweiz vorkommenden Chondrostoma-Arten. Mitteilungen zur Fischerei, 53.
Mesquita, N., Cunha, C., Hanfling, B., Carvalho, G. R., Ze-Ze, L., Tenreiro, R., & Coelho, M. M. (2003). Isola-
tion and characterization of polymorphic microsatellite loci in the endangered Portuguese fresh-water fish
Squalius aradensis (Cyprinidae). Molecular Ecology Notes, 3(4), 572–574. https://doi.org/10.1046/j.1471-
8286.2003.00515.x
Muenzel, F. M., Sanetra, M., Salzburger, W., & Meyer, A. (2008). Microsatellites from the vairone Leucis-cus
souffia (Pisces: Cyprinidae) and their application to closely related species. Molecular Ecology Notes, 7, 1048–
1050.
Pedroli J.C., Zaugg B., Kirchhofer A. (1991). Verbreitungsatlas der Fische und Rundmäuler der Schweiz;
Schweizerisches Zentrum für die kartografische Erfassung der Fauna
Peter A., Stölzel N., Wilmsmaier L., (2020) Fischökologische Wirkungskontrolle am Kraftwerk Aue an der
Limmat. Ingenieurbiologie: 37-44
Piry, S., Alapetite, A., Cornuet, J.-M., Paetkau, D., Baudouin, L., & Estoup, A. (2004). GeneClass2: A Software
for Genetic Assignment and First-Generation Migrant Detection. Journal of Heredity, 95, 536–539.
Pisci Page https://www.pivi.de/arten/fische/karpfenfische/nase/ abgerufen am 08.07.2021
Pritchard, J. K., Stephens, M., & Donnelly, P. (2000). Inference of Population Structure Using Multilocus Gen-
otype Data. Genetics, 155, 945–959.
Rey, P. (2019). Die Nase (Chondrostoma nasus) im Einzugsgebiet des Bodensees—Grundlagenbericht für in-
ternationale Maßnahmenprogramme; Internationale Bevollmächtigtenkonferenz für die Bodenseefischerei
(IBKF).
Schneider A. (1992) Food availability and diet compositon of 0+ Chondrostoma nasus in temporally variable
environment; Program and Abstracts; Ecotones Unesco Map; Lunz; Vienna; Austria.
20Sie können auch lesen

















































