DIPLOMARBEIT MAGISTRA DER PHARMAZIE - Pharmakotherapie der Adipositas Yasemin Celebi
←
→
Transkription von Seiteninhalten
Wenn Ihr Browser die Seite nicht korrekt rendert, bitte, lesen Sie den Inhalt der Seite unten
DIPLOMARBEIT zur Erlangung des akademischen Grades einer MAGISTRA DER PHARMAZIE an der Naturwissenschaftlichen Fakultät der Karl-Franzens-Universität Graz Pharmakotherapie der Adipositas vorgelegt von Yasemin Celebi Graz, Jänner 2023
DANKSAGUNG Ich möchte mich an dieser Stelle herzlich bei meiner Betreuerin Mag. Dr. rer. nat. Gabriele Schoiswohl für die Übernahme der Betreuung meiner Diplomarbeit und ihre wertvollen Verbesserungsvorschläge bedanken. Mein großer Dank gilt meiner liebevollen Familie, die mich bei der Anfertigung dieser Arbeit und während meines Studiums geduldig motiviert haben. Vielen Dank für ihre emotionelle und finanzielle Unterstützung.

KURZFASSUNG Die weltweit zunehmende Prävalenz von Adipositas, die als chronische Stoffwechselerkrankung mit assoziierten Folgeerkrankungen anerkannt ist, erhöht die Notwendigkeit von präventiven und wirksamen Therapien. Das Basisprogramm zur Behandlung von Adipositas, welches sich aus einer mit der Ernährungsumstellung und Bewegungstherapie zusammensetzt, ist bei der Gewichtsreduktion oft nicht ausreichend und zeigt in den meisten Fällen einen kurzfristigen Erfolg. Eine medikamentöse Behandlung von Adipositas kann diesen Maßnahmen unterstützen. Das Ziel der Pharmakotherapie ist die langfristige Reduktion des Körpergewichtes und Verbesserung der Adipositas-assoziierten Folgeerkrankungen. Aufgrund der möglichen unerwünschten oder teilweise noch unbekannten Nebenwirkungen wird die medikamentöse Therapie jedoch nur kurzfristig eingesetzt. Aufgrund dieser limitierten pharmazeutischen Behandlungsmöglichkeiten ist Untersuchung der pathophysiologischen Grundlagen der Adipositas unumgänglich, um neue Wirkstoffe mit verbesserter Verträglichkeit und Wirksamkeit entwickeln zu können. In dieser Diplomarbeit werden der Lipidstoffwechsel und die Regulation des Appetits, welche bei der Entstehung von Adipositas eine wichtige Rolle spielt, beschrieben. Des Weiteren beschäftigen sich diese Arbeit mit unterschiedlichen Behandlungsansätzen sowie zugelassene Medikamente und deren Wirkungsmechanismen für die Therapie von Adipositas. Abschließend werden neu-entwickelte anti-adipöse Wirkstoffe beschrieben, welche sich noch in klinischen Studien befinden.

ABSTRACT The increasing prevalence of obesity, which is recognized as a chronic metabolic disease with associated consequential health disorders, highlights the importance of preventive and effective therapies. The basic program for the treatment of obesity, which incorporates a dietary change and exercise therapy, is often not sufficient for weight loss and usually only shows short-term success. Thus, a medication-based treatment of obesity can be used as a support in conjunction with the basic program. The aim of pharmacotherapy is the long-term weight reduction and improvement of obesity-associated secondary diseases. However, due to the possible unwanted or partially unknown side effects, drug therapy is only used in the short-term. Given the limited options for pharmaceutical treatment, investigating the pathophysiological basis of obesity is unavoidable in order to develop new drugs with improved tolerability and efficacy. This diploma thesis describes the lipid metabolism and the regulation of appetite, which play an important role in the development of obesity. Furthermore, this thesis deals with different treatment approaches as well as approved medications and their mechanisms of action for the therapy of obesity. Lastly, newly developed anti-obesity substances are described, which are still in clinical trials.
Inhaltsverzeichnis 1. Das Fettgewebe und seine Funktion 7 1.1 Energie- und Fettaufnahme 13 2. Adipositas 17 2.1 Klassifikation der Adipositas 18 2.2 Ursachen 19 3. Appetitregulation im Hypothalamus 19 3.1 Die Kurz- und Langzeitregulation des Appetits 22 3.1.1 Kurzzeitregulation der Nahrungsaufnahme 22 3.1.2 Ghrelin 22 3.1.3 Peptid YY 22 3.1.4 Glucagon-like-Peptid-1 23 3.1.5 Glucose als Signal für Sättigung 23 3.2 Langzeitregulation der Nahrungsaufnahme 24 3.2.1 Leptin 24 3.3 Orexigene und Anorexigene Neuropeptide 25 3.3.1 Neuropeptid Y 26 3.3.2 Agouti-related-Peptid 26 3.3.3 Melanin-konzentrierendes Hormon 26 3.3.4 Orexin A / B 27 3.3.5 Das Peptid Galanin 27 3.3.6 Proopiomelanocortin und α-Melanozyten-stimulierendes Hormon 28 3.3.7 Serotonin 29 4. Medikamentöse Therapie: Antiadiposita 31 4.1 Entkoppler der Atmungskette 32 4.2 Indirekte Sympathomimetika 33 4.3 Cannabinoid-Rezeptor-1-Antagonist-Rimonabant 33
4.4 Lipasehemmer 35 4.5 Serotonin Agonist 36 4.6 Sympathomimetika / Antikonvulsiva 36 4.7 Opioid Rezeptor Antagonist / Dopamin und Noradrenalin Wiederaufnahme-Hemmer 37 4.8 GLP-1-Rezeptoragonisten 39 4.8.1 Liraglutid 39 4.8.2 Semaglutid 40 5. Neue medikamentöse Entwicklungen in der Adipositastherapie 40 5.1 Leptin-Sensitizer 40 5.1.1 Der Wirkstoff Celastrol 41 5.1.2 Der Wirkstoff Withaferin A 42 5.2 Ghrelin 43 5.2.1 Der Ghrelin-Antikörper 43 5.2.2 Das Anti-Ghrelin Spiegelmer Nox-B11 43 5.3 Y2-Rezeptoragonisten 44 5.4 Melanocortin-4-Rezeptor Agonist 44 6. Zusammenfassung 45 7. Abkürzungsverzeichnis 47 8. Abbildungsverzeichnis 49 9. Tabellenverzeichnis 50 10. Literaturverzeichnis 51
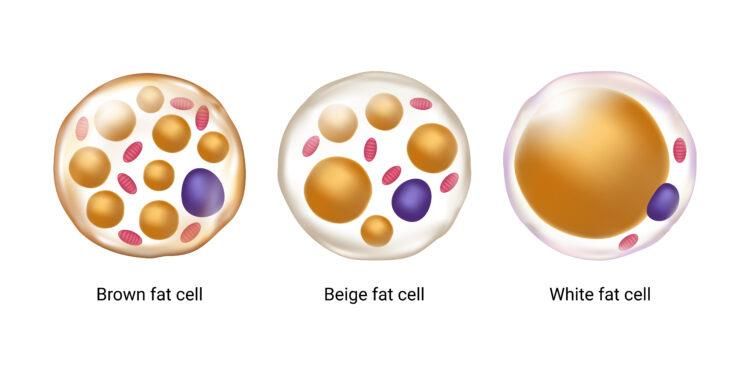
1. Das Fettgewebe und seine Funktion Das Fettgewebe ist ein metabolisch aktives Gewebe, welches reife Adipozyten (Fettzellen), Vorläuferzellen der Adipozyten (Prä-Adipozyten), Blutgefäße, Nervenzellen und Immunzellen wie Makrophagen beinhaltet. [1] Es gibt zwei Arten von Fettgewebe, die aus braunen und weißen Adipozyten bestehen. Das Fettgewebe erfüllen viele wichtige Funktionen im menschlichen Körper wie Energiespeicherung und Energiebereitstellung, mechanischer Schutz, Wärmeisolation, endokrine Aktivität, Thermogenese und Speicherung von fettlöslichen Vitaminen (A, D, E und K). [2] Abbildung 1: Braunen, beigen und weißen Adipozyten; Braune und Beige Adipozyten besitzen einen Kern (blau), viele Mitochondrien (rot) und kleine Lipidtröpfchen (gelblichbraun). Weiße Adipozyten haben ein großer Lipidtropfen, der die gesamte Zelle ausfüllt und somit die wenige Mitochondrien und den Kern an die Peripherie dräng. aus [3] Das weiße Fettgewebe wird in subkutanes und viszerales Fettgewebe unterteilt. Das subkutane Fettgewebe liegt in der Subkutis und wird auch als Unterhautfettgewebe bezeichnet. [4] Es ist unter anderem für die Wärmeisolation verantwortlich. Die Fettverteilung des subkutanen Fettgewebes ist geschlechtsabhängig. [5] Es befindet sich bei Frauen vor allem an der Brust sowie im Bereich des Gesäßes. Bei Männern kommt es am Bauch und im Nacken vor. Das viszerale Fettgewebe befindet sich hauptsächlich in der Bauchhöhle und wird auch als intraabdominales Fettgewebe bezeichnet. Es bietet einen mechanischen Schutz, indem es die innere Organe umschließt. Im Gegensatz zu subkutanem Fett kann übermäßiges viszerale Fett das Risiko für verschiedene Krankheiten wie Herzerkrankungen, Bluthochdruck und Diabetes mellitus Typ 2 erhöhen. [6] 7
Weiße Adipozyten sind hauptsächlich für die Speicherung von Lipiden in Form von Triglyceriden und deren Abbau bei erhöhtem Energie Bedarf verantwortlich. [7]Sie besitzen einen großen Lipidtropfen (Univakuoläre Adipozyten) [Abb. 1], indem unter anderem Triglyceride, Diglyceride und Cholesterinester gespeichert werden. Der Lipidtropfen ist mit einem Monolayer aus Phospholipiden vom Zytoplasma begrenzt. An diese Phospholipide sind Proteine aus der PAT-Familie wie Perilipin und Adipophilin assoziiert. Das Intermediärfilament Vimentin stabilisiert die Form des Lipidtropfen. [8] Die gespeicherten Lipide im Fettgewebe können aus der Nahrung stammen oder werden selbst in den Adipozyten synthetisiert. Triglyceride sind chemisch gesehen ein Ester, der aus einem Molekül Glycerin und drei gesättigten oder ungesättigten Fettsäuren aufgebaut ist. Biosynthese (Lipogenese) und Speicherung von Triglyceriden sind stark regulierte Stoffwechselvorgänge, die unter anderem durch das Hormon Insulin reguliert werden. [9] [Abb. 2] Die Aktivierung des Insulin-Rezeptors mit Insulin stimuliert die PIK3-Aktivät, wodurch der Glucose-Transporter (GLUT4-Transporter) vom Zytoplasma zur Plasmamembran transloziert und somit Glukose aufgenommen werden kann. [10] Die aufgenommene Glucose wird in der Glykolyse, welche im Zytosol stattfindet, verstoffwechselt und durchläuft mehrere Prozessierungsschritte mit den wichtigen Stoffwechselzwischenprodukten Dihydroxyaceton-Phosphat (DHAP) und Glycerinaldehyd-3-Phosphat (GAP). DHAP wird zu GAP isomerisiert, welches die Glykolyse weiter durchläuft und letztendlich zu Pyruvat verstoffwechselt wird. Zusätzlich kann das entstandene DHAP für die Synthese von Glyzerol-3- Phosphat (G-3-P) verwendet werden, welches in den Adipozyten essenziell für die Lipogenese ist. Das entstandene Pyruvat wird anschließend in den Mitochondrien durch die oxidative Decarboxylierung zu Acetyl-CoA umgewandelt, welches in den Citratzyklus eingeschleust wird und Citrat entsteht. Das Citrat wird in das Zytosol transportiert, welches dort mithilfe der ATP-Citrat Lyase wieder zu Acetyl-CoA umgewandelt wird. Das Acetyl-CoA ist das Ausgangsgerüst für die de novo Synthese von Fettsäuren. Hierbei wird Acetyl-CoA zu Malonyl-CoA und anschließend zur Fettsäure Palmitate konvertiert. Palmitat stellt somit die erste und einfachste neu synthetisierte Fettsäure in der de novo Lipogenese dar. Anschließend unterläuft Palmitat diverse Prozessierungsvorgänge zur Synthese von unterschiedlich langen und komplexen Fettsäuren. [11] Die neu gebildeten 8
Fettsäuren werden zu Acyl-CoA aktiviert und mit dem ebenfalls neu gebildetem G-3- P verestert, um Triglyceride zu bilden. Der erste und limitierende Schritt in der Triglycerid-Synthese ist die Bildung von Lysophosphatidsäure (LPA) durch die Veresterung der ersten Hydroxylgruppe des G-3-P Rückgrates mit Acyl-CoA LPA wird anschließend mit einem weiteren Acyl-CoA zu Phosphatidsäure (PA) acyliert. Über einen Zwischenschritt wird PA dephosphoryliert, wodurch ein Diglycerid entsteht. Der letzte Schritt der Triglycerid-Synthese ist die erneute Veresterung des letzten Kohlenstoffes des G-3-P mit einem Acyl-CoA. [12] Zusätzlich zu den de novo synthetisierten Fettsäuren werden auch Fettsäuren aus der Nahrung für die Triglycerid Synthese herangezogen. Diese freien Fettsäuren werden im Blutplasma an Albumin gebunden und transportiert. Über spezifische Fettsäure Transporter werden diese in die Adipozyten aufgenommen und transportiert, zu Acyl-CoA aktiviert und mit G-3-P verestert (siehe oben). Ebenso werden auch Fettsäuren von Triglyceriden aus der Nahrung für die Triglycerid- Synthese verwendet. Die Triglyceride aus der Nahrung werden als Bestandteil Chylomikronen im Blut transportiert. Die Adipozyten-membranständige Lipoproteinlipase hydrolysiert die Triglyceride der Chylomikronen und die dabei entstehenden freien Fettsäuren werden von den Fettsäure Transportern aufgenommen und anschließend für die nachstehende Synthese aktiviert. [13] 9
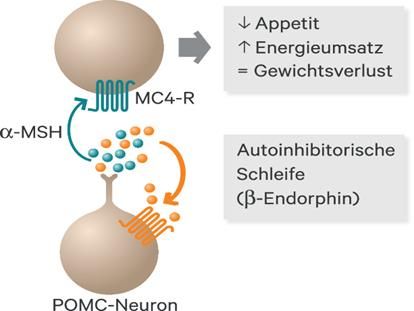
Abbildung 2: Lipogenese und Lipolyse in Adipozyten: Lipoproteine Chylomikronen und VLDL werden durch Lipoproteinlipase hydrolysiert und die freien Fettsäuren gelangen in die Adipozyten. Diese werden in mehreren Schritten in Speicherform (Triglyceride) umgewandelt. Der Abbau von Triglyceriden wird durch Insulin und Catecholamine wie Adrenalin und Noradrenalin reguliert. Durch hormonsensitive Lipasen werden sie wieder in freie Fettsäuren und Glycerin abgebaut; aus [14] Bei Energiebedarf können die gespeicherten Triglyceride wieder mobilisiert werden. Diesen Vorgang nennt man Lipolyse (Abbau von Triglyceriden). [15]Dabei wirkt das Hormon Noradrenalin lipolytisch und das Hormon Insulin anti-lipolytisch. [16] Die Adipozyten besitzen β3-Adrenorezeptoren und durch Noradrenalin wird die Lipolyse aktiviert. Die G-Protein-gekoppelten β-Rezeptoren aktivieren die Adenylatcyclase und über Proteinkinase A werden die Lipasen aktiviert. Die gespeicherten Triglyceride durch die Adipocyten-Triglyceridlipase, die hormonsensitive Lipase und die Monoglyceridlipase in Fettsäuren und Glycerin hydrolytisch gespalten. Die Fettsäuren werden von den Adipozyten freigesetzt und gelangen über das Blut zu unterschiedlichen Organen, wie Herz, Muskel, Leber, wo sie je nach Bedarf für Synthese herangezogen werden. Das Glycerin aus der Lipolyse wird in der Leber für die Neusynthese von Glucose (Gluconeogenese) verwendet. [17][Abb. 2] 10
Das braune Fettgewebe ist ein plurivakuoläres Fettgewebe. [18]Das braune Fettgewebe macht bei Neugeborenen etwa 5 % des Körpergewichtes aus. Bis zum Erwachsenenalter kommt es zur Abnahme der braunen Fettmasse. Sie befindet sich nur noch um die Arterien, im Brustkorb, am Hals, im Nacken, an den Nieren und unter den Achseln. [19] Die braunen Adipozyten besitzen mehrere kleine Lipidtropfen und Mitochondrien. Diese typische Zellfarbe der Adipoztyen (gelb- bräunlich) ist von der Mitochondrienzahl abhängig. [Abb. 1] Die Hauptaufgabe der braunen Adipozyten ist die Produktion von Wärme, die sogenannte Thermogenese. [20] Mit der Energie aus der Atmungskette wird durch das Protein Thermogenin Wärme produziert. Das Membranprotein Thermogenin wird auch uncoupling protein 1 (UCP-1) bezeichnet. Hier dient es als Entkoppler. [21] Außer den weißen und braunen Adipozyten existiert noch eine weitere Form von Adipozyten. Sie werden beige oder brite Adipozyten (aus „brown“ und „white“) benannt. Beige Adipozyten entstehen bei Kältereiz aus weißen Adipozyten. Diese Adipozyten sind wie braunen Adipozyten multivakuolär und mitochondrienreich aber in das weiße Fettgewebe eingestreut. [Abb.1] Sie besitzen auch UCP-1 und können somit Wärme produzieren. [22] Eine weitere wichtige Funktion des Fettgewebes ist die sekretorische Funktion. Als stoffwechselaktives und endokrines Regulationsorgan setzt das Fettgewebe zahlreiche Sekretionsprodukte frei. [Abb. 3] Etwa 100 solcher Botenstoffe aus Adipozyten sind bisher entdeckt. [23] Viele davon werden als Adipokinine und Adipozytokine bezeichnet. Die Stoffe wirken nicht nur autokrin und parakrin, sondern auch endokrin und werden über das Blut zu ihren Zielorganen transportiert, wo sie und viele Prozesse im Stoffwechsel wie den Lipid- und Glukosemetabolismus, Blutdruckregulation, Immun- und Komplementsystem und Insulinsensitivität beeinflussen. [24] 11
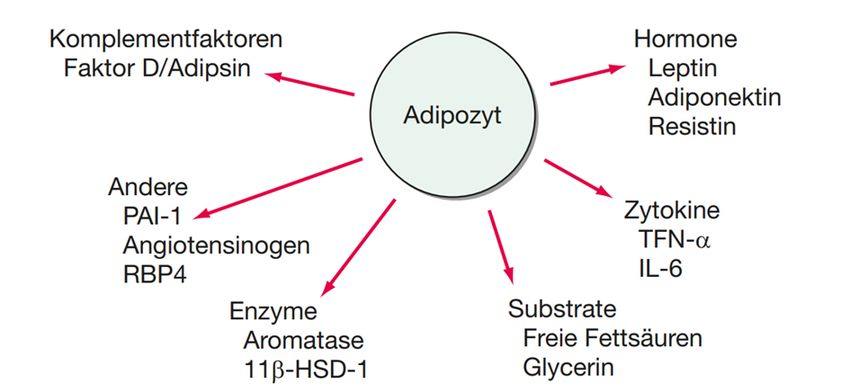
Abbildung 3: Mediatoren aus Adipozyten: Adipozyten sezernieren zahlreiche spezifische Mediatoren, wie die Hormone Leptin, Resistin, Adiponektin und Vaspin, welche die Energiebilanz regulieren. Adipozyten sekretieren auch mehrere Entzündungsmediatoren wie Tumor-Nekrose-Faktor α (TNF-α) und Interleukin 6 (IL-6), Komplementfaktoren wie Faktor D (auch als Adipsin bekannt), prothrombotische Gerinnungsfaktoren wie Plasminogen- Aktivator-Inhibitor 1 (PAI-1) und vasoaktive Proteine des Renin-Angiotensin-Systems wie Angiotensinogen; aus [25] Als erstes beschriebenes Adipokin wurde Leptin 1994 entdeckt. [26] Leptin wirkt appetithemmend, wobei ein Sättigungsgefühl im Hypothalamus ausgelöst wird. Es wird proportional zur Fettmasse gebildet. Das Fettzellhormon Adiponektin, dessen Konzentration bei Patienten mit Adipositas erniedrigt ist, erhöht die Insulinsensitivität in der Muskulatur und steigert die Lipidoxidation. Zusätzlich besitzt es antiinflammatorische und gefäßschützende Eigenschaften. Resistin und Retinol- bindendes Protein (RBP4), deren Konzentrationen bei Adipositas erhöht sind, verursachen Insulinresistenz. Eine erhöhte Konzentration von dem Adipokin Vaspin, einen Serinprotease-Inhibitor, führt zur Hyperglykämie (Überzuckerung) und Verminderung der Nahrungsaufnahme. [27]Die vermehrte Freisetzung von Plasminogen-Aktivator-Inhibitor I aus den Adipozyten führt zu Blutgerinnungs- störungen und weiters zu einer Hyperkoagulabilität mit Thromboembolien. Die Anschaltung des Renin-Angiotensin-Aldosteron-Systems (RAAS) durch Freisetzung von Angiotensinogen führt zur arteriellen Hypertonie (Bluthochdruck). Die Freisetzung von proinflammatorischen Adipozytokine wie Tumornekrosefaktor- α (TNF-α), Interleukin 6 (IL-6) und anderen Zytokinen können zu chronischer 12
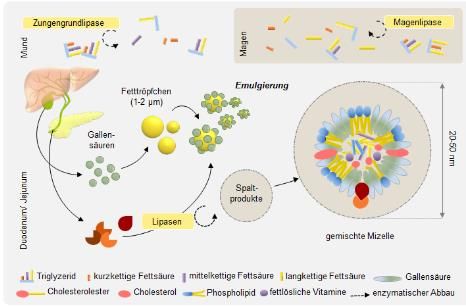
Inflammation des Fettgewebes. [25] Zusammenfassend, beeinflussen die von Adipozyten freigesetzten Mediatoren die Lipidhomöostase, Insulinsensitivität, die Regulation des Blutdrucks, Gerinnung und Gefäßfunktion und spielen bei der Entstehung der Folgeerkrankungen bei adipösen Patienten eine wesentliche Rolle. 1.1 Energie- und Fettaufnahme Mit der Nahrung werden nicht nur Nährstoffe, sondern auch die benötigte Energie für die Grundfunktionen des Körpers aufgenommen. Bei Energieüberschuss werden Nahrungsfette in Form von Lipiden im Fettgewebe und die Glukose als Glykogen in Muskel- und Leberzellen gespeichert. Nach der Nahrungsaufnahme finden die physiologische Verdauungs- und Resorptionsprozesse von Kohlenhydraten, Proteinen und Fetten statt. Abbildung 4: Verdauung und Resorption der Fette Teil 1: Durch Zungengrundlipasen und Magenlipasen werden die Nahrungsfette gespalten. Spaltprodukte und Gallensäure bilden gemischte Mizellen. aus [29] Die Fettverdauung beginnt bereits im Mund mit der Sekretion der Zungengrundlipase. [28] Durch gastrische Lipasen werden die Fette teilweise im Magen abgebaut. Die freie Fettsäuren, welche nach der Hydrolyse von Triglyceriden entstehen, stimulieren die Freisetzung des Gewebshormons Cholecystokinin aus endokrinen Zellen (I-Zellen) des Duodenum und Jejunum. Dies löst die Freisetzung von Pankreasenzymen (Trypsin und Chymotrypsin) und eine Kontraktion der 13
Gallenblase aus. Die Fett-Moleküle werden durch die Pankreaslipase in Glycerin und kurz- bzw. mittelkettige Fettsäuren gespalten. Durch die Gallensäure werden die Fette emulgiert und es entstehen kleine amphiphile Fettkügelchen oder Mizellen genannt. [Abb.4] Die Mizellen werden von den Zellen (Enterozyten) des Dünndarms resorbiert und die darin gespeicherten langkettige Fettsäuren und Monoglyceride werden wieder zu Triglyceriden verestert. [28] [ Abb.5] Abbildung 5: Verdauung und Resorption der Fette Teil 2: Die gebildeten Mizellen werden von den Enterozyten des Dünndarms aufgenommen. Resynthese von Triglyceriden finden in den Mukosazellen; aus [29] Die resynthetisierte Triglyceride werden zusammen mit Cholesterin, Cholesterinestern, Phospholipiden und Apolipoprotein B-48 in Chylomikronen eingebaut. [Abb.6] Die Chylomikronen werden von Lymphgefäßen aufgenommen und gelangen über den Ductus thoracicus in den systemischen Kreislauf. Im Blut erhalten sie noch den Cofaktor der Lipoproteinlipase Apolipoprotein C-II (Apo C-II). [30] 14
Abbildung 6:Chylomikronen sind Produkte des exogenen Lipoproteinmetabolismus Sie enthalten Trigylceride, Colesterin, Cholesterinestern, Phospholipiden und Apolipoproteine ApoB-48, ApoE und ApoC II. Lipoprotein-Lipase( LPL) spaltet Triglyceride der Lipoproteine in Fettsäuren und Gylcerin. ApoC II dient als Cofaktor für diese enzymatische Reaktion ; aus [31] Kurz- und mittelkettige Fettsäuren sind aufgrund ihrer geringen Größe hydrophiler und werden über den Pfortaderkreislauf zur Leber transportiert. Dort werden sie mit Cholesterin, Lipiden mit Phosphatgruppe und den Apolipoproteinen ApoB100, ApoC1und ApoE zu Very Low Density Lipoprotein (VLDL) vereinigt und in die Blutbahn geschleust. In der Blutbahn wird noch Apolipoprotein C-II eingebaut. [31] Abbildung 7: Produkte des endogenen Lipoproteinmetabolismus: Lipoproteine VLDL und IDL enthalten Apolipoproteine ApoE und ApoB-100. Das Lipoprotein LDL besitzt nur mehr ApoB-100. aus [31] Das extrazelluläre Enzym Lipoproteinlipase befindet sich auf der Endothelzellmembranen von Blutkapillaren des Herzens, der Muskulatur und des 15
Fettgewebes und katalysiert die Hydrolyse von Triglyceriden aus Lipoproteinen (VLDL und Chylomikronen). Die freigesetzten Fettsäuren werden in die Zellen (z.B. Adipozyten) aufgenommen. [siehe Kapitel 1: S.2] Die Überreste von Chylomikronen, die Chylomikronen-Remnants werden mithilfe des LDL-Rezeptors durch rezeptorvermittelte Endozytose in die Hepatozyten aufgenommen und vollständig abgebaut werden. Aus den VLDL entstehen zunächst IDL (intermediate density proteins). IDL können durch weitere Abspaltung von Triglyceriden in LDL (low density lipoproteins) prozessiert werden und über einen LDL-Rezeptor zurück zur Leber. [31] [28] [Abb. 7] Die Verdauung von Kohlenhydraten (Polysaccharide) beginnt in der Mundhöhle mit der α-Amylase, welche sich im Speichel befindet. Hierbei werden die Polysaccharide in Oligo- und Disaccharide zerlegt. Im Magen wird die Verdauung aufgrund des sauren Milieus unterbrochen, da das Enzym Amylase kann bei sauren pH-Werten nicht mehr arbeiten kann. Erst im Dünndarm wird der weitere Abbau mit dem alkalischen Pankreassaft fortgesetzt, indem er den sauren Nahrungsbrei neutralisiert. Die α-Amylase der Pankreas baut die Oligosaccharide und Disaccharide zu Monosacchariden ab. Die entstandene Monosaccharide Glucose, Galaktose und Fructose gelangen in die Darmwandzellen. [32] Die Resorption von Glucose und Galactose in den Enterozyten erfolgt mittels Natrium-gekoppelten Transporter aktiv. Fructose wird von einem spezifischen Transporter durch erleichterte Diffusion in die Zellen aufgenommen. Danach werden die Monosaccharide durch einen Glukosetransporter aus der Zelle ins Blut und über die Pfortader zur Leber transportier. [28] Glucose spielt eine wesentliche Rolle bei der verstoffwechselt und die dabei entstandenen Zwischenprodukte und Fettsäuren zusammen zu Triglyceriden verestert. [siehe Kapitel 1: S.2] 16
2. Adipositas Adipositas (von lateinisch adeps = Fett) oder Fettleibigkeit ist eine chronische Stoffwechselerkrankung, die durch eine vermehrte Ansammlung von Körperfett charakterisiert ist. Sie ist eine global verbreitete Erkrankung aller Bevölkerungsschichten und Altersgruppen. Weltweit leben aktuell fast zwei Drittel der Erwachsenen mit Übergewicht oder Fettleibigkeit. [33] Der europäische Adipositas-Bericht 2022 der WHO zeigt, dass 59 % der Erwachsenen und 29 % der Kinder in der Europäischen Region übergewichtig sind oder mit Adipositas leben. [34]Der Körper braucht eine regelmäßige Energiezufuhr zur Aufrechterhaltung der Lebensfunktionen. Die hierfür benötigte Energie wird entweder aus der Nahrung bezogen oder es werden körpereigene Energiereserven mobilisiert. Die überschüssige Energie wird in den Adipozyten als Fettdepot gespeichert, wobei ein einziger Adipozyt sich bis auf das 200-fache vergrößern kann. [35]Bei erhöhtem Energiebedarf wird die gespeicherte Energie der Triglyzeride in Form von freien Fettsäuren ins Blut abgegeben. Wird das Gleichgewicht zwischen der Energieaufnahme und dem Energieverbrauch gestört, kann dies über einen längeren Zeitraum zu einer Gewichtsabnahme oder einer Gewichtszunahme führen. Im Fall von Adipositas führt eine Dysbalance zwischen Energiezufuhr und Energiebedarf zu einer pathologischen Vermehrung bzw. Ansammlung von Körperfett. Die Energiebalance ist vor allem von genetischen Faktoren sowie vom Lebensstil wie übermäßige Nahrungsaufnahme, Bewegungsmangel und Stress aber auch von hormonellen Erkrankungen (z.B. Hypothyreose und Cushing-Syndrom) abhängig. Adipöse Patienten leiden auch unter Adipositas-assoziierten Begleiterkrankungen wie Diabetes mellitus Typ 2, nicht- alkoholischer Fettleber, Polyzystisches-Ovar- Syndrom, Schlaf-Apnoe-Syndrom, Hypertonie, Arthrose, Depression, Sodbrennen und Herzinsuffizienz. [36] 17
2.1 Klassifikation der Adipositas Adipositas wird anhand des Body Mass Index (BMI) ermittelt. Der BMI oder auch Körpermasseindex genannt, wird als Quotient aus Kilogramm (kg) Körpergewicht und dem Quadrat der Körpergröße in Metern (m) definiert: [37] Körpergewicht (kg) = ö öß ( )2 Der BMI wird wie folgt eingeteilt: Tabelle 1: Gewichtsklassifaktion der WHO mit BMI: aus [38] Gewichtskategorie BMI (kg/m²) Normalgewicht 18,5-24,9 Übergewicht (Präadipositas) 25 – 29,9 Adipositas Grad I 30-34,9 Adipositas Grad II 35 – 39,9 Adipositas Grad III ≥40 Adipositas (Grad I und II) wird definiert mit einem BMI zwischen 30-39,99 kg/m2. Ein BMI über 40 kg/m2 deutet auf eine schwere Form der Adipositas (Grad III) hin. [Tabelle 1] Neben der Körperfettmasse spielt auch die Verteilung des Fettgewebes eine entscheidende Rolle für die Entwicklung adipositasbedingter Folgeerkrankungen. Bei der abdominalen Adipositas (auch „Apfeltyp“ genannt) befindet sich die Fettanlagerung vorwiegend im Bereich der Bauchregion und rund um die inneren Organe. Diese Form tritt hauptsächlich bei Männern auf. Daher wird sie auch als androide Fettverteilung bezeichnet. Die bauchbetonte Adipositas kann Herz- Kreislauf-Störungen, Diabetes mellitus Typ-2, Bluthochdruck und Fettleber begünstigen. Bei der peripheren Adipositas (auch gynoide Adipositas oder „Birnentyp“ genannt) sammelt sich das Fett vermehrt im Bereich der Oberschenkel und Hüften an und tritt häufiger bei Frauen auf. Dieser Form der Fettgewebesverteilung erhöht das Risiko für Gefäßerkrankungen. [39] 18
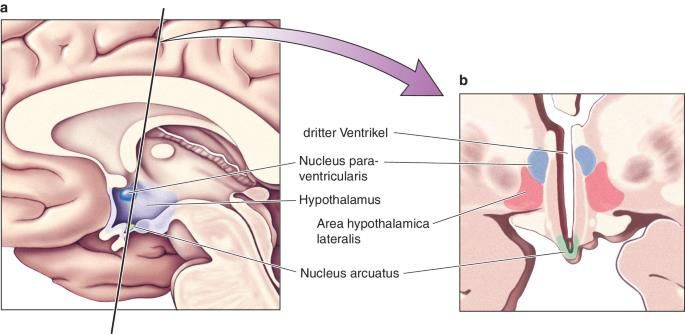
2.2 Ursachen Adipositas ist eine vielseitige Krankheit. Ätiologisch wird zwischen primärer und sekundärer Adipositas unterschieden. [40] Bei der primären Adipositas (mit der Prävalenz von 95 %) spielen genetische Disposition, Bewegungsmangel, übermäßiger Konsum von kalorienreichen Lebensmitteln aber auch psychische Faktoren wie Stress, Essstörungen (Binge-Eating-Störung) oder depressive Erkrankungen eine Rolle. [39]Bei etwa 5 % der Patienten mit Adipositas Grad III handelt es sich um eine monogenetische Erkrankung, welche durch einen Defekt in einem einzigen Gen ausgelöst wird. Bei diesen Genen handelt es sich um das Leptin-Gen LEP, das Leptinrezeptor-Gen, das Melanocortin-4-Rezeptor-Gen und das Proopiomelanocortin-Gens. Mutationen in diesen Genen können für die frühkindlich auftretende Adipositas verantwortlich sein (childhood-onset obesity). [41] Die sekundäre Adipositas wird durch endokrine Erkrankung hervorgerufen und macht etwa 5 % der Fälle aus. Die häufigen Ursachen der sekundären Adipositas sind meist hormonell bedingte Stoffwechselerkrankungen wie Schilddrüsenunterfunktion, Cushing-Syndrom oder Sexualhormonmangel. Außerdem können Medikamente wie Antihistaminika, Psychopharmaka, Betablocker, Antidiabetika und Glucocorticoide eine sekundäre Adipositas begünstigen. Zusätzlich besteht bei vielen adipösen Patienten eine psychische Essstörung, die sogenannte Binge-Eating-Störung, die zu regelmäßig auftretenden Essanfälle führen. [42] 3. Appetitregulation im Hypothalamus Der Hypothalamus (von altgriechisch hypo unter und thálamos Zimmer) befindet sich im Zwischenhirn (Diencephalon) und liegt unterhalb des Thalamus. [43] Die Verbindung zwischen dem Hypothalamus und der Hypophyse wird von dem Hypophysenstiel übernommen. [44] Der Hypothalamus ist das wichtigste Regulationszentrum des vegetativen Nervensystems. Er kontrolliert und reguliert lebenswichtigen Körperfunktionen wie Atmung, Kreislauf, Körpertemperatur, Hormonhaushalt, Schlaf, Sexualfunktionen und auch die Nahrungsaufnahme. 19
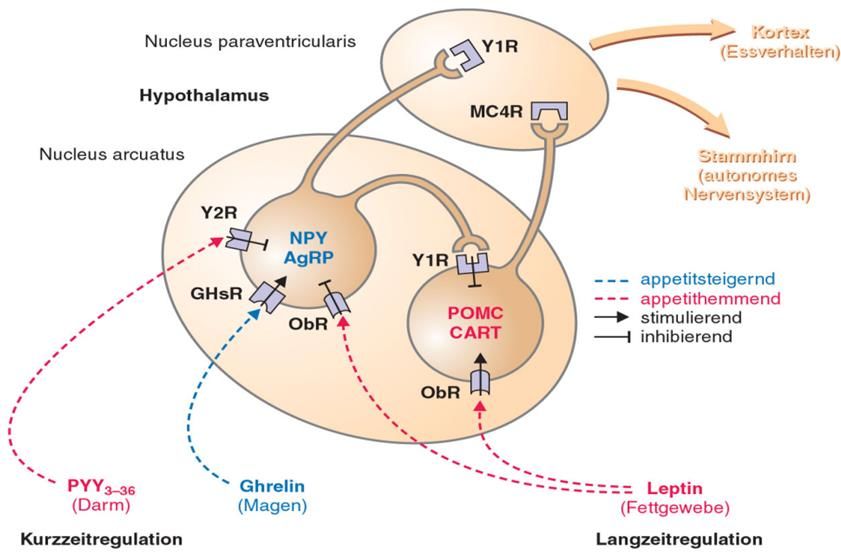
Abbildung 8:a) Die Lage des Hypothalamus im Gehirn. b) im Querschnitt: aus [45] Er besitzt verschiedene Nuclei oder auch Kerngebiete, die in der Neuroanatomie eine Ansammlung von Nervenzellkörpern im zentralen Nervensystem bezeichnet werden. Diese Kerngebiete sind von einer weißen Substanz (Substantia alba) umgegeben. [46] Die wichtigen Kerngebiete des Hypothalamus sind: Nucleus arcuatus, dorsomedialis, paraventricularis, periventricularis, suprachiasmaticus, supraopticus und ventromedialis. Im Hypothalamus werden releasing Hormone und Inhibiting Hormone, welche die Ausschüttung anderer Hormone in die Hypophyse fördern oder hemmen. Die releasing Hormone des Hypothalamus sind das Growth hormone-releasing Hormon, Gonadotropin-releasing Hormon, das Thyreotropin- releasing Hormon und das Corticotropin-releasing Hormon. Zu den inhibiting Hormone gehören Somatostatin und das Prolaktin-inhibiting Hormon. [47] An der zentralnervösen Regulation des Appetits sind die Regionen des Nucleus ventromediale (VMH), Nucleus arcuatus (ARC), Nucleus paraventricularis (PVN) und lateralen Hypothalamus-Areale (LHA) beteiligt. [48]Das Hungerzentrum befindet sich im lateralen Hypothalamus. Für die Sättigung ist der ventromediale Hypothalamus zuständig. Lokale Läsionen im Bereich des ventromediale Hypothalamus und des Nucleus arcuatus haben einen Einfluss auf die Nahrungsaufnahme. Eine Läsion im ventromedialen Nucleus, der eine große Anzahl von Leptin-Rezeptoren (Ob-Rb) besitzt, führt zu Hyperphagie und schlussendlich zu Adipositas. [49] Im Gegensatz dazu führt eine Läsion im lateralen Hypothalamus zu Hypophagie und damit zu Gewichtverlust. [50] Der Nucleus Arcuatus, ein Regulator der Energie-Homöostase, 20
liegt oberhalb der Eminentia mediana, wo die Blut-Hirn-Schranke leicht durchgängig ist und somit die Signalmoleküle aus der Peripherie zu den Neuronen (Nervenzellen) des Nucleus arcuatus gelangen können. Der Nucleus arcuatus leitet mit dem Nucleus paraventricularis die Informationen für Appetit zur Großhirnrinde (Kortex) weiter. [51][Abb.9] Abbildung 9: Die Kurz- und Langzeitregulation des Appetits im Hypothalamus. Die appetitregulierenden Neuronen sind mit ihren Rezeptoren im Nucleus arcuatus und paraventricularis lokalisiert. Sie werden durch Leptin, Ghrelin und PYY reguliert: aus [52] Der Nucleus arcuatus besitzt unterschiedliche Neuronengruppen, die Agouti-related Protein (AgRP), Neuropeptid Y (NPY), Proopiomelanocortin (POMC) und das Cocain- und Amphetamin-regulierende Transkript (CART) freisetzen. Die orexigenen Peptide, NPY und AgRP, sind Appetitstimulatoren und senken den Energieverbrauch des Körpers, wohingegen POMC und CART eine appetithemmende Wirkung besitzen und somit den Energieverbrauch erhöhen. Die Aktivität der Neuronen wird durch die peripheren Peptidhormone wie Leptin, Ghrelin und Peptid YY (PYY) reguliert. [48] [51] [Tabelle 2] 21
3.1 Die Kurz- und Langzeitregulation des Appetits Die Nahrungsaufnahme wird mithilfe verschiedener Hormone und Neurotransmittern in den Regulationszentren des zentralen Nervensystems reguliert. Dabei wird zwischen der Kurzzeitregulation und der Langzeitregulation unterschieden. Die Kurzzeitregulation ist für den akuten Hunger- und Sättigungszustand des Körpers zuständig. Signale, die bei der akuten Appetitregulation beteiligt sind, können durch externe Reize (optische und olfaktorische Wahrnehmungen) oder durch interne Reize (Blutzuckerspiegel, Füllungszustand des Magens und Darms) ausgelöst werden. Dabei spielen die Peptidhormone Ghrelin, Peptid YY, Glucagon-like-Peptide (GLP-1) und die Glucosekonzentration im Blut eine wichtige Rolle. Die Langzeitregulation des Appetits wird hingegen durch das Sättigungs-hormon Leptin gesteuert. [51] Tabelle 2: Hormone, die Nahrungsaufnahme steigern (Orexigene) bzw. vermindern (Anorexigene); aus [48] Orexigene Ghrelin Anorexigene Leptin, Glukagon-Like-Peptide, Peptid YY 3.1.1 Kurzzeitregulation der Nahrungsaufnahme 3.1.2 Ghrelin Ghrelin (Akronym, englisch: Growth Hormone Release Inducing) ist ein gastrointestinales Hormon, welches postprandial in Zellen der Magenschleimhaut produziert wird. Der Ghrelin-Spiegel im Blut steigt im Hungerzustand und sinkt nach der Nahrungsaufnahme. [53]Ghrelin gelangt über die Blut-Hirn-Schranke ins Gehirn, wo es mit hoher Affinität an seinen Growth-Hormon-Secretagogue-Rezeptoren (GHS-R) im Hypothalamus bindet und eine erhöhte Ausschüttung von Neuropeptid Y und Agouti-related-Peptid im Nucleus arcuatus bewirkt. Dadurch wird der Appetit gesteigert. [54] 3.1.3 Peptid YY 22
Das Peptid Tyrosin-Tyrosin (PYY) besteht aus 36 Aminosäuren und gehört zur Gruppe der pankreatischen Polypeptiden. Das Peptid enthält die Aminosäure Tyrosin am N- und C-Terminus in der Primärstruktur. Da im Einbuchstaben-Code die Aminosäure Tyrosin mit Y gekennzeichnet ist, kam es zur Namensgebung von Peptid Tyrosin-Tyrosin bzw. Peptid YY. [55]Das appetithemmende Hormon Peptid YY1-36 wird postprandial von endokrinen L-Zellen im Dünndarm und Dickdarm gebildet. Nach der Abspaltung durch das proteolytisches Enzym Dipeptyl-Peptidase IV (Peptid YY1-36 wird zu Peptid YY3-36 gespalten) wird das Hormon biologisch aktiv. PYY ist dem Neuropeptid Y strukturell sehr ähnlich. PYY bindet an Y2–Rezeptoren und bewirkt eine Hemmung von orexigenen Neuropeptid Y-Neuronen. Dadurch wird das Sättigungsgefühl gesteigert. Die klinischen Untersuchungen zeigen, dass die PYY-Konzentration bei übergewichtigen Patienten niedrig und bei Personen mit Untergewicht deutlich erhöht ist. Als ein spezifischer Agonist der Y2-Rezeptoren bietet das Peptid YY3-36-Analogon eine wichtige therapeutische Alternative für Patienten mit Adipositas und Diabetes Mellitus Typ 2. [56] 3.1.4 Glucagon-like-Peptid-1 GLP-1 wird nahrungsabhängig von den neuroendokrinen L-Zellen des Darms produziert und gehört zur Gruppe der gastrointestinalen Hormone (Inkretine). Bei erhöhter Glukosekonzentration steigert die Insulinsekretion aus den Betazellen der Bauchspeicheldrüse. Außerdem hemmt das Peptidhormon die Glukagon-Sekretion und verzögert die Magenentleerung. [57]GLP-1-Rezeptoren befinden sich im Pankreas, Hypophyse, Magen, Dünndarm, Gehirn, Lunge, Nieren, Myokard und Endothelzellen.Die Bindung an GLP-1-Rezeptoren im Bereich der Area postrema führt zu vermindertem Appetit. [58] 3.1.5 Glucose als Signal für Sättigung Die Glukosekonzentration im Blut kann die Regulation der Nahrungsaufnahme beeinflussen. Im Gehirn, im Darm und in der Leber befindet sich Sensoren, die für 23
die Messung der Glukosekonzentration zuständig sind. [59] Wenn der Glukose-Wert unter dem Soll-Wert (zwischen 100 und 125 mg/dl) liegt, wird ein Hungergefühl ausgelöst, wohingegen das Erreichen des Soll-Wertes ein Sättigungsgefühl hervorruft. Bei normalem Insulinwert führt eine Erhöhung der Glukosekonzentration zu einer Hemmung des Hungergefühls und zu einer Steigerung des Sättigungsgefühls Dadurch wird ein Zustand von Sättigung erreicht. Im anderen Fall löst eine verminderte Glucosekonzentration ein Hungergefühl aus. [60] 3.2 Langzeitregulation der Nahrungsaufnahme 3.2.1 Leptin Das Polypeptidhormon Leptin (von griech. leptós, dünn) wird hauptsächlich von den weißen Adipozyten produziert und ins Blut abgegeben. [61] Es bindet an den löslichen Leptin-Rezeptoren (Slep-R; soluble Leptin Receptor) und überwindet die Blut-Hirn-Schranke. [62] [63]Rezeptoren für Leptin (Ob-Rs) befinden sich in beiden Neuronengruppen des Nucleus arcuatus. Es besteht eine Korrelation zwischen der Leptin-Konzentration im Blut und der Körperfettmasse, da eine Zunahme der Fettzellmasse die Bildung von Leptin steigert. In Folge führt eine erhöhte Leptin- Konzentration zur Stimulierung der POMC-Expression, wodurch die appetitsteigernden NYP/AGRP-Neuronen gehemmt werden. Dies führt zur Appetitunterdrückung und Gewichtsabnahme. Bei niedrigerer Leptin-Konzentration kommt es zu einer verminderten POMC-Expression und die NYP/AGRP-Neuronen werden wieder aktiviert. Dies führt zu einer Steigerung des Appetits und somit zu einer gesteigerten Gewichtszunahme. [64]Ähnlich wie die Insulinresistenz bei der erworbenen Form von Diabetes bewirkt eine andauernd hohe Leptin-Konzentration ein vermindertes Ansprechen der Nervenzellen auf das Sättigungshormon Leptin was als Leptinresistenz bezeichnet wird. [65]Aufgrund der reduzierten Leptin Sensitivität kommt es trotz erhöhter Leptin-Konzentration zu keinem Sättigungsgefühl bei adipösen Menschen. Deshalb wird Leptinresistenz als eine der Hauptursachen für Adipositas gesehen. Daher ist es wichtig die Ursachen für Leptinresistenz zu bestimmen. Die Wirkstoffe Withaferin A und Celastrol sind als Leptin-Sensitizer bereits identifiziert. [66] Weiters kann eine Mutation im Leptin-Gen LEP zu einem angeborenen Leptinmangel führen. [67] Da der Körper das Hormon nicht mehr produzieren kann, sind eine schwere Hyperphagie mit früh entstehender 24
Adipositas die Folge. Bei Patienten mit biologisch inaktivem Leptin ist das Hormon zwar im Blut nachweisbar jedoch besitzt es keine biologische Aktivität mehr. Patienten mit Leptinmangel werden mit dem Wirkstoff Metreleptin, ein rekombinantes humanes Leptin-Analogon, behandelt. Das Arzneimittel steht als Injektionslösung zur Verfügung. [68]Zusätzlich zu Mutationen im Leptin-Gen, können Mutationen im Leptinrezeptor zur Beeinträchtigung der Leptin Sensitivität führen. Die Mutationen von Leptinrezeptoren bewirken eine erhöhte Nahrungsaufnahme, da die anorexigene Wirkung von Leptin ausbleibt. Der Wirkstoff Setmelanotid ist zur Behandlung von Adipositas aufgrund des Proopiomelanocortinmangels oder Leptin- Rezeptor-Mangels zugelassen. [69] 3.3 Orexigene und Anorexigene Neuropeptide Man unterscheidet zwischen orexigenen und anorexigenen Neuropeptide bzw. Cotransmittern. Im Hypothalamus regulieren sie die Nahrungsaufnahme und den Energiehaushalt des Körpers. Orexigene (anabole) Neuropeptide steigern den Appetit und verringern den Energieverbrauch. Anorexigene (katabolische) Neuropeptide steigern den Energieverbrauch und unterstützen das Abnehmen. [48] [Tabelle 3] Tabelle 3: Neuropeptide/-transmitter, welche die Nahrungsaufnahme steigern (Orexigene) bzw. vermindern (Anorexigene). aus [48] Orexigene Neuropeptid Y, Agouti-related-Peptid, Orexin A und B Melanin-Concentraiting Hormon (MCH), Galanin Anorexigene Cocaine-und Amphetamine-related Transcript Proopiomelanocortin, Serotonin, α-Melanozyten-stimulierendes Hormon (α-MSH) 25
3.3.1 Neuropeptid Y Der Appetitstimulator NPY besteht aus 36 Aminosäuren. Die NPY-produzierende Nervenzellen kommen vorwiegend im Nucleus arcuatus und im Nucleus paraventricularis vor. Freisetzung von NYP aus ARC-Neuronen wird durch Leptin und Peptid YY gehemmt, während Ghrelin stimulierend wirkt. [70]Das Neuropeptid hat 6 verschiedene G-Protein-gekoppelten Rezeptoren Y1-Y6, die in unterschiedlichen Geweben vorkommen. Im Gehirn befindet sich der Rezeptor Y5, der für die Appetitregulation wichtig ist. Rezeptor Y4 kommt im Darm vor und reguliert die Darm-Motilität. [55] Neben der Regulation der Nahrungsaufnahme und des Körpergewichts hat NYP zahlreiche Funktionen im Zentralennervensystem, unter anderem fördert es die Freisetzung von Insulin und Glucocorticoide, verbessert die kognitiven Leistungsfähigkeiten (Y2), kontrolliert Krampfanfälle und wirkt antikonvulsiv (Y1, Y5), hat analgetische (Y1, Y2), anxiolytische und sedierende Wirkung (Y1). Y1 und Y5- Rezeptorantagonisten werden als potenzielle Arzneistoffe zur Behandlung von Adipositas gesehen. [71] 3.3.2 Agouti-related-Peptid Das Agouti-related-Peptid ist ein aus 132 Aminosäuren bestehendes Neuropeptid, welches im Nucleus arcuatus gebildet wird. Die Freisetzung von AgRP wird durch Leptin gehemmt und durch Ghrelin gefördert. [72] Es hat eine antagonistische Wirkung an Melanocortin-3,4-Rezeptoren. Mit dem α-Melanozyten-stimulierenden Hormon bindet AgRP an die gleichen Rezeptoren und wirkt appetitfördernd. [73] Dementsprechende zeigten AgRP-Transgene Mäuse eine gesteigerte Nahrungsaufnahme. Die Blockade von AgRP-Rezeptoren führte hingegen zu einer massiven Unterdrückung der Nahrungsaufnahme. [48] 3.3.3 Melanin-konzentrierendes Hormon Das zyklische Polypeptid Melanin-konzentrierendes Hormon, das auch als Melanin- Concentrating Hormon (kurz: MCH) bezeichnet wird, wird von Neuronen des 26
lateralen Hypothalamus und der Zona incerta synthetisiert. Ursprünglich wurde MCH in Fischen identifiziert. Dort beeinflusst das Hormon die Pigmentation, weshalb es als Melanin-konzentrierendes Hormon bezeichnet wurde. [74]MCH-Expression wird durch Leptin reguliert, wobei ein Leptinmangel die MCH-Synthese fördert. [48] Die Bindung von MCH an MCH-1 und MCH-Rezeptoren bewirkt eine Steigerung des Appetits. Deshalb sind MCH-Rezeptoren interessante Angriffspunkte für neuronale Appetitsteuerung. Zurzeit werden die Substanzen racSNAP-7941, GW3430 und T- 226296 für ihre antagonistische Wirkung an MCH-Rezeptoren untersucht. [75] 3.3.4 Orexin A / B Die zwei Neuropeptide Orexin A und B (Hypokretin 1 und 2) sind im lateralen und dorsamedialen Hypothalamus zu finden. Sie werden durch enzymatische Spaltung aus einem Propeptid, Pre-Pro-Orexin gebildet. Orexin wird durch das HCRT-Gen kodiert. [76] Orexin hat einen starken Einfluss auf das Essverhalten und den Schlafrhythmus, indem es die Nahrungsaufnahme und auch den Wachzustand erhöht. [48] 3.3.5 Das Peptid Galanin Galanin ist ein Neuropeptid mit orexigenen Effekt und besteht aus 30 Aminosäuren. ARC und PVN Neuronen produzieren Galanin. Die Wirkung von Galanin wird über drei G-Protein-gekoppelte Rezeptoren: GALR1, GALR2 und GALR3 vermittelt. [77]Im Hungerzustand steigt die Expression von Galanin und stimuliert die Nahrungsaufnahme. Es wurde eine Korrelation zwischen Verlangen nach fettreicher Nahrung und der Galaninkonzentration im Nucleus paraventricularis festgestellt. Die Hemmung von Galanin-Rezeptoren führte bei Ratten zur verminderten Nahrungsaufnahme. [78] Es kommt auch im Gastrointestinaltrakt vor, wobei es dort die Motilität beeinflusst. [79] 27
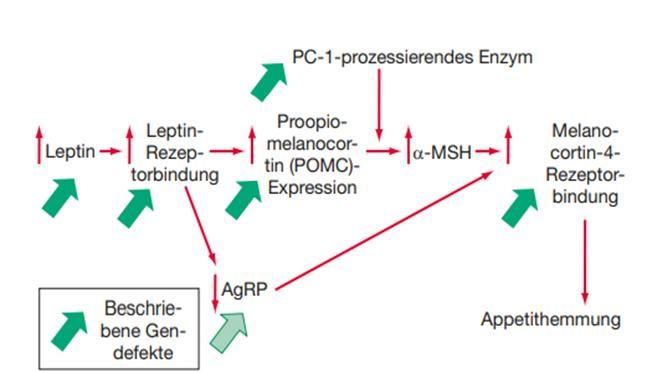
3.3.6 Proopiomelanocortin und α-Melanozyten-stimulierendes Hormon Das aus 267 Aminosäuren bestehende Protein Proopiomelanocortin kommt in der Haut, im Magendarmtrakt und in vielen anderen Geweben vor. Das Prähormon wird zellspezifisch in ihre Derivate gespalten: in Adrenocorticotropes Hormon (ACTH), Corticotropin-like intermediate Peptide (CLIP), α, β- MSH, Lipotropin und Beta- Endorphin. [80]Zuerst spaltet das Enzym Prohormonconvertase 1 (PC1) das adrenocorticotrope Hormon enzymatisch ab. α-MSH, besteht aus 13 Aminosäuren, wird nach der Aufspaltung des ACTH durch Prohormonconvertase 2 gewonnen. [81] [Abb.10] Abbildung 10: Proopiomelanocortin und seine Derivate. Aus POMC können ACTH, MSH, Lipotropin, CLIP und β-Endorphin entstehen; aus [82] Die Freisetzung von POMC und α-MSH wird durch das Hormon Leptin geregelt. Nach der Bindung von Leptin an seinen Rezeptor kommt es zur Expression von Poopiomelanocortin. Dies wird durch Prohormonconvartase 1 zu α-MSH prozessiert. Die Bindung von α-MSH an Melanocortin-4-Rezeptoren bewirkt eine Appetithemmung. [ Abb.11] Der Mangel an POMC durch eine Mutation im POMC- Gen kann zu Übergewicht führen. Auch Mutationen im Melanocortin-4-Rezeptor und in der Prohormonconvartase 1 können Hyperphagie und Adipositas verursachen. [25] 28
Abbildung 11: Das Melanocortinsystem; zentrale Regulation von Appetit und Körpergewicht . Die Bindung von Leptin an seinen Rezeptor führt zur Aktivierung von POMC, das mit Enzym PC-1 zu α-MSH gepalten wird. Bindung α-MSH an MC4R führt zur Appetithemmung. Die grünen Pfeile zeigen die bekannte Gendefekte, die zur Adipositas führen; aus [25] Setmelanotid, ein Analogon von α-MSH, aktiviert die Melanocortin-4-Rezeptoren, wodurch der Appetit kontrolliert wird. Der Wirkstoff wird zur Behandlung von genetisch bedingtem Proopiomelanocortin - oder Leptinrezeptor (LEPR)-Mangel eingesetzt. [69] 3.3.7 Serotonin Serotonin, auch 5-Hydroxytryptamin (5-HT) oder Enteramin genannt, wirkt im zentralen Nervensystem als Neurotransmitter. Als Gewebshormon wird es in den enterochromaffinen Zellen der Darmmukosa gebildet. Da es die Blut-Hirn- Schranke nicht passieren kann, wird in serotonergen Neuronen Serotonin durch Hydroxylierung und Decarboxylierung aus der Aminosäure Tryptophan synthetisiert. [83] Die Aufnahme von Tryptophan ins Gehirn wird durch Insulin gefördert. Es wurde festgestellt, dass Personen mit Übergewicht eine niedrigere Serotonin-Konzentration aufweisen. [84] 29
Abbildung 12: 5-HT-Wirkungsstellen auf Melanocortinweg. 5-HT aktiviert POMC- Neuronen über die Aktivierung von 5-HT2C -Rezeptoren 5-HT hyperpolarisiert und hemmt AgRP-Neuronen; aus [85] Die Serotoninrezeptoren des Subtyps 5-HT1B und 5-HT2C sind bei der Appetitregulation wichtig. Für die eigentliche appetitsenkende Wirkung von Serotonin sind die Serotoninrezeptoren 5-HT1B oder 5-HT2C, die auf den POMC- und AgRP- Neuronen lokalisiert sind, verantwortlich. [ Abb.12] Nach der Aktivierung von 5-HT2C- Rezeptoren werden die POMC-Neuronen stimuliert. Dies führt zur Freisetzung von α-MSH, das wiederum die Melanocortin-4-Rezeptoren aktiviert und das Sättigungsgefühl fördert. Die Aktivierung von 5-HT1B-Rezeptoren durch Serotonin wirkt inhibitorisch auf AgRP-Neuronen, die appetitfördern sind. [85] 30
4. Medikamentöse Therapie: Antiadiposita Antiadiposita (Schlankheitsmittel) sind Arzneistoffe für die Behandlung von Übergewicht (BMI 25‒29,9 kg·m−2) und Adipositas (BMI ≥ 30 kg·m−2). Sie werden meistens unterstützend und zeitlich begrenzt mit dem Ziel einer Gewichtreduktion angewendet. Die Wirkstoffe haben unterschiedliche Wirkungsmechanismen, aber erzielen den gleichen Effekt, unkontrollierbaren Appetit zu hemmen oder das Sättigungsgefühl zu steigern. [86] Tabelle 4: Einteilung der Antiadiposita nach Wirkungsmechanismus. aus [87] Substanzklasse /Substanz Firmenname Zulassung Entkoppler der Atmungskette 2,4-Dinitrophenol Stanford University 1933–1938 (USA) Sympathamimetika Fenfluramin und Dexfenfluramin Wyeth Ayerst 1973–1997 (USA) Sibutramin Abbott Laboratories 1997–2010(USA, EU) Phentermin Teva Pharmaceuticals 1959–aktuell in USA Cannabinoid-Rezeptor-1-Antagonist Rimonabant Sanofi SA 2006–2009 (EU) Lipase-Hemmer Orlistat Roche Pharmaceuticals 1999–aktuell (USA, EU) 5-HT2C Serotonin Agonist Lorcaserin Arena Pharmaceuticals, Eisai 2012–2020 (USA) Sympathomimetika / Antikonvulsiva Phentermin und Topiramat Vivus 2012-aktuell (USA) Opioid Rezeptor Antagonist / Dopamin und Noradrenalin Wiederaufnahmehemmer Naltrexone SR/Bupropion SR Orexigen Therapeutics Inc 2014–aktuell (USA, EU) GLP-1-Rezeptoragonisten Liraglutid Novo Nordisk 2014–aktuell (USA, EU) Semaglutid Novo Nordisk 2021 (USA) 31
4.1 Entkoppler der Atmungskette 2,4-Dinitrophenol, eine Industriechemikalie, war das erste synthetische Arzneimittel, welches zur Gewichtsabnahme in USA eingesetzt wurde. [88] An Arbeitern, welche 2,4-Dinitrophenol in Sprengstofffabriken verarbeitet haben, wurde eine extreme Gewichtsabnahme beobachtet. Diese gewichtreduzierende Wirkung von 2,4-Dinitrophenol wurde vom Pharmakologe Maurice L. Tainter von der Stanford University im Jahr 1933 erkannt. [89] Abbildung 13: Strukturformel von 2,4-Dinitrophenol. [90] 2,4-Dinitrophenol hebt die Kopplung von Elektronentransport und ATP- Synthese in den Mitochondrien auf, indem es den Protonentransport aus dem Intermembranraum in die Matrix durch passive Diffusion als Protonophore (Protonentranslokator) erleichtert. Es wird der Aufbau der Protonengradienten verhindert und somit die ATP-Synthese verringert, aber gleichzeitig die Wärmeproduktion (Thermogenese) gesteigert. [91] Die Entkopplung führt zu einer Steigerung des Kohlenhydrat- und Fettstoffwechsels, um ATP-Mangel zu kompensieren. 2,4-Dinitrophenol hat eine geringe therapeutische Breite und ist hochgiftig. Mögliche schwere Nebenwirkungen sind: Herzrhythmusstörungen, Tachykardie, Atemnot, Kopfweh, Hyperthermie, Übersäuerung des Blutes, Zerfall der quergestreiften Muskulatur, Hyperglykämie, Grauer Star, Nierenversagen, Leberversagen und letztendlich Multiorganversagen. [92] Aufgrund der lebensbedrohlichen Nebenwirkungen wurde das Mittel 1938 nach nur 5 Jahren vom Markt genommen. [87] 32
4.2 Indirekte Sympathomimetika Der appetithemmende Wirkstoff Sibutramin hat eine Phenylethylaminstruktur und hat somit auch eine strukturelle Ähnlichkeit mit älteren Appetitzügler wie Amphetamin, Phenylpropanolamin und Phentermin. [93] Abbildung 14: Strukturformel von Sibutramin [94] Sibutramin ist ein Noradrenalin/Serotonin‐Wiederaufnahmehemmer. [95] Die Hemmung der Wiederaufnahme von beiden Neurotransmittern führt zu einer Erhöhung der Neurotransmitterkonzentration im synaptischen Spalt, wodurch die Adrenorezeptoren verstärkt aktiviert werden. Als ein indirekt wirkendes Sympathomimetikum bewirkt es eine Verstärkung des Sättigungsgefühls und eine Steigerung der Fettverbrennung. [96]Sibutramin wurde im Jahr 1997 in den USA und später in der EU zugelassen. [87] Der Wirkstoff war unter dem Medikamentennamen Reductil® im Handel. Das Arzneimittel wurde wegen der lebensbedrohlichen Nebenwirkungen auf das Herz-Kreislauf-System wie Herzinfarkt, Schlaganfall und Erhöhung des Blutdrucks vom Markt genommen (1997-2010). [97] 4.3 Cannabinoid-Rezeptor-1-Antagonist-Rimonabant Das Endocannabinoid System reguliert viele physiologische Prozesse, wie Schlaf, Schmerzempfindung, Stimmung, Motorik, Gedächtnis, Nahrungsaufnahme und Apppetitregulation. [98]Die Cannabinoid (CB)-Rezeptoren und Endocannabinoiden 33
als Liganden bilden das Endocannabinoid System. Der Cannabinoidrezeptor‐Typ I (CB‐1), ein G‐Protein gekoppelter Rezeptor, kommt im Gehirn, sowie in anderen Geweben wie z.B. im Fettgewebe, in der Leber und in den Muskeln vor. Die Aktivierung der CB-1-Rezeptoren im zentralen Nervensystem führt zur Appetitsteigerung. Dieser appetitsanregende Effekt von Tetrahydrocannabinol (THC), ein Phytocannabinoid der Hanfpflanze, ist seit längerem bekannt. Die periphere Aktivierung der Rezeptoren stimuliert die Lipogenese in den Adipozyten. [99] Durch die Blockade der CB-1-Rezoptoren mit Rimonabant kann die Überaktivierung moduliert werden und es kommt zur Appetithemmung, zum Abbau von Fettreserven und zum Anstieg des Adiponektinspiegels. [100] Strukturell handelt es sich um ein chloriertes Piperidin und Pyrazolcarboxamid-Derivat, das ursprünglich zur Raucherentwöhnung von dem Pharmaunternehmen Sanofi-Aventis entwickelt wurde. Abbildung 15: Strukturformel von Rimonabant [99] Übelkeit, Durchfall, Schwindel und Atemwegsinfektionen gehören zu den somatisch unerwünschten Nebenwirkungen. Die Substanz wurde wegen ihrer psychiatrischen Nebenwirkungen wie Depression, Suizidgedanken und Gedächtnisstörungen vom Markt genommen. [99] 34
4.4 Lipasehemmer Bei Orlistat (=Tetrahydrolipstatin) handelt es sich um ein peripher wirkendes Antiadipositum, das spezifisch gastrointestinalen Lipasen hemmt. [101] Abbildung 16: Strukturformel von Orlistat. [102] Lipasen sind Serin-Hydrolasen und besitzen im aktiven Zentrum eine katalytische Triade aus Serin, Histidin und Aspartat. Als Enzyminhibitor bindet Orlistat an das katalytische Zentrum, wo es die nukleophile Hydroxyl-Gruppe des Serins am ß- Lacton-Ring unter Ausbildung einer kovalenten Bindung angreift, und somit das Enzym irreversible hemmt. [103] Abbildung 17: Wirkungsmechanismus von Orlistat. [102] Die inaktivierten Lipasen können die Esterbindungen der Nahrungsfette nicht mehr hydrolytisch spalten und somit wird das Fett unverdaut ausgeschieden. Bei der Einnahme von Orlistat wird meist von Magen-Darm-Beschwerden wie Fettstühle, Durchfall, Blähungen mit Stuhldrang und Bachschmerzen berichtet. [104] Außerdem 35
kann es bei Dauereinnahme zur Unterversorgung von fettlöslichen Vitaminen (A, D, E und K) kommen. [105] 4.5 Serotonin Agonist Mit serotonerger Wirkung bindet Lorcaserin selektiv an den Serotonin-Rezeptoren vom Typ 5-HT2C, welche sich auf den POMC-Neuronen befinden. [85] Weiter wird der Melanocortin-Weg, welcher zu einer Erhöhung des Sättigungsgefühls und einer Appetithemmung führt, aktiviert. Außerdem wird die Ausschüttung orexigener Mediatoren wie Neuropeptid Y und Agouti-releated peptid verhindert. [106] Abbildung 18: Strukturformel von Lorcaserin. [106] Depression, Euphorie, Selbstmordgedanken und gastrointestinale Beschwerden sind bekannte Nebenwirkungen von Lorcaserin. Ergebnisse aus Karzinogenitätstests mit Ratten zeigen ein erhöhtes Krebsrisiko. [107] Der Wirkstoff wurde nur in den USA (2012-2020) als Anorektikum verwendet, in Europa wegen Sicherheitsbedenken nicht zugelassen. [108] 4.6 Sympathomimetika / Antikonvulsiva Phentermin wurde bereits 1959 von der amerikanischen Zulassungsbehörde FDA für einen Gebrauch von bis zu 12 Wochen für Personen über 16 Jahren als Appetitzügler zugelassen. [109] Der Wirkstoff Phentermin gehört zur Substanzklasse der Amphetamine (Amine mit „aufweckender“ Wirkung). [110] Es ist ein indirektes Sympathomimetikum und stimuliert die vermehrte Freisetzung von Neurotransmittern wie Adrenalin, Noradrenalin und Dopamin aus den 36
präsynaptischen Nervenzellen, wodurch seine appetithemmende Wirkung im Zentralnervensystem entfaltet. Abbildung 19: Strukturformel von Phentermin (oben) und Topiramat(unten) [111] Phentermin wurde ursprünglich in Kombination mit Fenfluramin und Dexfenfluramin als Medikament gegen Übergewicht verwendet. Aufgrund der kardialen Nebenwirkung (Herzklappenfehlern) und pulmonaler Hypertonie wurden die Wirkstoffe Fenfluramin/Dexfenfluramin verboten. (1973-1997 USA) [112] [87] Phentermin wird jetzt in Kombination mit Topiramat als Medikament gegen Übergewicht verwendet. Es ist aber nur in den USA zugelassen. Topiramat ist ein Sulfamat substituiertes Monosaccharid, welches zur Behandlung von Epilepsie und Migräneprophylaxe eingesetzt wird. Als Nebenwirkung kommt es zu einer Minderung des Appetits und zusammen mit Phentermin unterschützt es die Gewichtreduktion. [111] 4.7 Opioid Rezeptor Antagonist / Dopamin und Noradrenalin Wiederaufnahme-Hemmer Bupropion als selektiver Noradrenalin- und Dopamin-Wiederaufnahmehemmer wird vor allem zur Raucherentwöhnung und zur Behandlung von Depression verordnet. [113]Naltrexon als Opioid-Antagonist wird hauptsächlich in der Therapie von Alkohol- und Opiatabhängigkeit eingesetzt. Seit 2018 wird Bupropion SR (360mg/ Tag) mit Naltrexon SR (32mg/ Tag mit verzögerter Freisetzung) zur Behandlung von 37
Sie können auch lesen

















































