Einfluss von Fumarsäureestern auf T-Zellsubpopulationen bei Patienten mit Psoriasis
←
→
Transkription von Seiteninhalten
Wenn Ihr Browser die Seite nicht korrekt rendert, bitte, lesen Sie den Inhalt der Seite unten
Aus der Klinik für Dermatologie und Allergologie
Geschäftsführender Direktor: Prof. Dr. M. Hertl
des Fachbereichs Medizin der Philipps-Universität Marburg
Einfluss von Fumarsäureestern auf
T-Zellsubpopulationen bei
Patienten mit Psoriasis
Inaugural-Dissertation zur Erlangung des Doktorgrades der
gesamten Humanmedizin
dem Fachbereich Medizin der Philipps-Universität Marburg
vorgelegt von
Stella Riemann
aus Lübeck
Marburg, 2021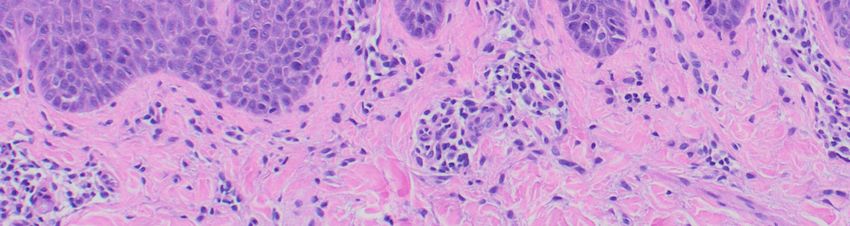
Angenommen vom Fachbereich Medizin der Philipps-Universität Marburg am: 19.03.2021 Gedruckt mit Genehmigung des Fachbereichs Medizin Dekanin: Frau Prof. Dr. D. Hilfiker-Kleiner Referent: Herr Prof. Dr. R. Eming 1. Korreferent: Herr Prof. Dr. M. Schnare

Inhaltsverzeichnis
Abkürzungsverzeichnis ................................................................................ I
Abbildungsverzeichnis ............................................................................. IV
Tabellenverzeichnis .....................................................................................V
Zusammenfassung .................................................................................... VI
Summary .................................................................................................. VIII
1 Einleitung ............................................................................................... 1
1.1 Psoriasis ........................................................................................................ 1
1.1.1 Klinik der Psoriasis vulgaris ........................................................................................... 1
1.1.2 Psoriasis guttata ................................................................................................................ 1
1.1.3 Psoriasis pustulosa palmaris et plantaris.................................................................. 2
1.1.4 Erythrodermia psoriatica ................................................................................................ 2
1.1.5 Psoriasis arthropathica .................................................................................................... 3
1.2 Epidemiologie .............................................................................................. 3
1.3 Pathophysiologie der Psoriasis .................................................................. 5
1.3.1 T-Zellen ................................................................................................................................. 5
1.3.2 Das antimikrobielle Peptid LL37 .................................................................................. 9
1.3.3 Mechanismen bei der Entstehung von psoriatischen Hautläsionen............ 10
1.4 Diagnostik .................................................................................................. 12
1.5 Therapie der Psoriasis ............................................................................... 13
1.5.1 Basistherapie ..................................................................................................................... 13
1.5.2 Topische Therapie ........................................................................................................... 14
1.5.3 Fototherapie ...................................................................................................................... 14
1.5.4 Systemische Therapie .................................................................................................... 14
2 Material ................................................................................................ 18
2.1 Patientenmaterial ...................................................................................... 18
2.2 Lösungen und Zusätze zur Kultivierung humaner Zellen ..................... 18
2.3 Zytokine und Proteinantigene ................................................................. 18
2.4 Chemikalien ................................................................................................ 18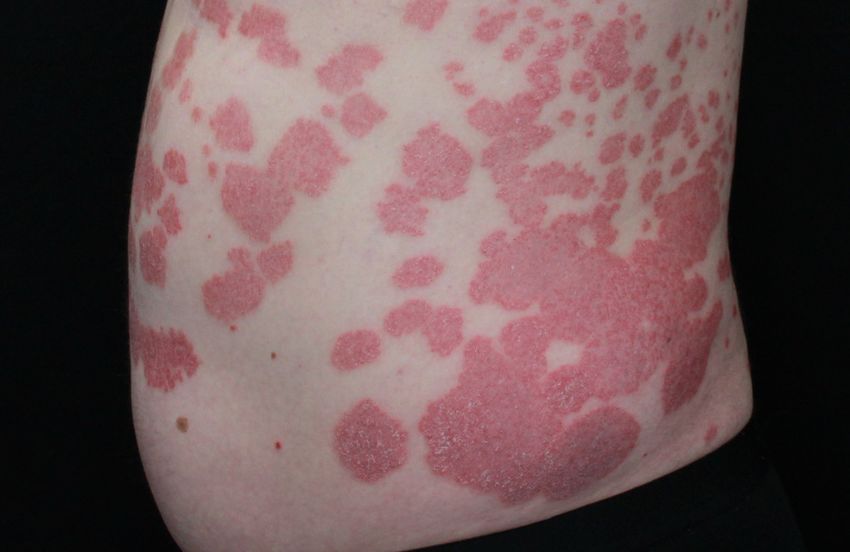
2.5 Puffer und Lösungen ................................................................................. 19
2.6 Antikörper .................................................................................................. 19
2.7 Kommerzielle Tests für die zelluläre Diagnostik .................................... 19
2.8 Verbrauchsartikel....................................................................................... 19
2.9 Geräte .......................................................................................................... 20
2.10 Software...................................................................................................... 20
3 Methoden............................................................................................. 21
3.1 Patienten..................................................................................................... 21
3.1.1 Einschlusskriterien für die Studienteilnahme........................................................ 22
3.1.2 Ausschlusskriterien für die Studienteilnahme ...................................................... 22
3.2 Studiendesign ............................................................................................ 22
3.3 Klinische Evaluation................................................................................... 23
3.4 Untersuchungsmethoden ......................................................................... 24
3.4.1 Zellseparation mittels Dichtegradienten-Zentrifugation ................................. 24
3.4.2 Bestimmung der Zellzahl.............................................................................................. 25
3.4.3 Kryokonservierung .......................................................................................................... 25
3.4.4 Auftauen von peripheren mononukleären Zellen............................................... 25
3.4.5 Frequenzanalyse zytokinsezernierender Zellen mittels ELISPOT .................. 26
3.4.6 Durchführung am Beispiel des antigenspezifischen ELISPOT ........................ 27
3.4.7 Frequenzanalyse nach antigenunabhängiger Zellstimulation ....................... 27
3.4.8 Frequenzanalyse nach Zugabe von Fumarsäurederivaten in vitro ............... 28
3.4.9 Auswertung mittels ELISPOT-Reader....................................................................... 28
3.5 Statistik ....................................................................................................... 29
4 Ergebnisse ............................................................................................ 30
4.1 Klinisches Ansprechen auf die Therapie mit Fumarsäureestern ........... 30
4.2 Auswahl des optimalen Stimulus für die ELISPOT-Analyse .................. 31
4.3 Stimulusabhängige T-Zellfrequenzverläufe unter
Therapie mit Fumarsäureestern................................................................ 34
4.4 LL37-spezifische T-Zellen bei Patienten mit Psoriasis ........................... 38
4.5 Auswirkungen von Fumarsäureestern in vitro ....................................... 40
5 Diskussion ............................................................................................ 46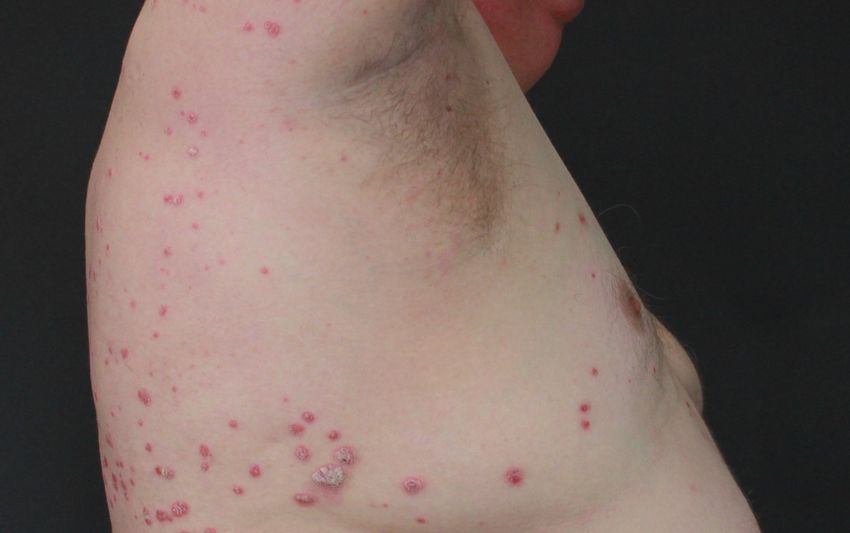
5.1 Klinische Effizienz der Therapie mit Fumarsäureestern ........................ 46
5.2 Effekte der Fumarsäureester-Therapie auf T-Zellebene........................ 48
5.2.1 Besonderheiten des ELISPOT-Verfahrens .............................................................. 48
5.2.2 T-Zell-Deviation bei Patienten mit Psoriasis ......................................................... 49
5.2.3 Frequenzverläufe von T-Zellsubpopulationen unter
Fumarsäureester-Therapie .......................................................................................... 50
5.3 LL37 als Autoantigen bei Patienten mit Psoriasis .................................. 53
5.4 In vitro Effekte von Fumarsäureestern auf T-Zellsubpopulationen ..... 55
5.4.1 Effekte von Dimethylfumarat ...................................................................................... 56
5.4.2 Effekte von Monomethylfumarat .............................................................................. 57
5.4.3 Effekte von Fumarsäure ................................................................................................ 57
5.4.4 Dimethylfumarat und Monomethylfumarat im Kontext
der in vivo Situation ...................................................................................................... 58
5.5 Fazit ............................................................................................................. 59
6 Anhang ................................................................................................. 61
7 Literaturverzeichnis ............................................................................ 63
8 Verzeichnis der akademischen Lehrer .............................................. 77
9 Danksagung ......................................................................................... 78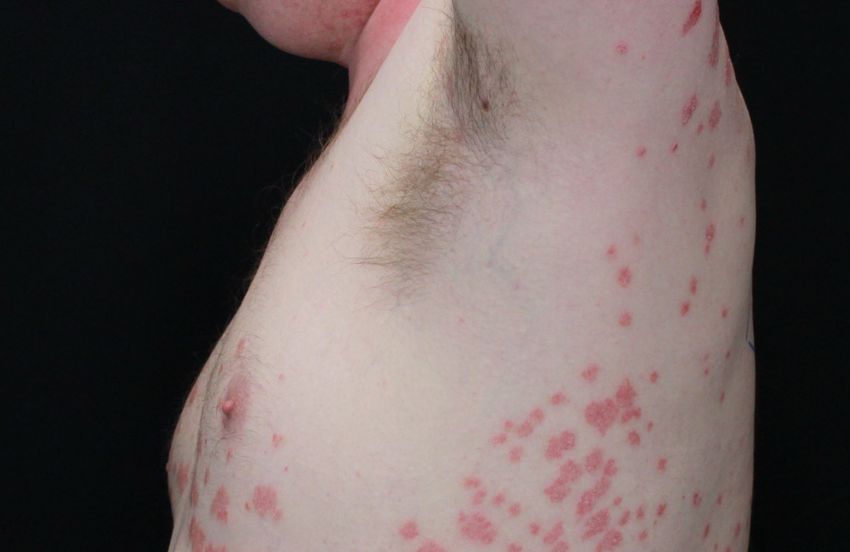
Abkürzungsverzeichnis
Abkürzungsverzeichnis
°C Grad Celsius
Abb. Abbildung
AMP Antimikrobielles Peptid
AP Alkalische Phosphatase
APC engl.: antigen presenting cell (Antigen-präsentierende Zelle)
Bcl-2 engl.: B-cell lyphoma 2 (B-Zell-Lymphoma 2)
BSA engl.: body surface area (Körperoberfläche)
Bovines Serumalbumin
bspw. beispielsweise
bzw. beziehungsweise
ca. circa
CCL27 CC-Chemokinligand 27
CCR CC-Chemokinrezeptor
CD engl.: cluster of differentiation (Differenzierungscluster)
cm Zentimeter
CO2 Kohlenstoffdoxid
CPDA Citrat-Phosphat-Dextrose-Adenin
CTLA engl.: cytotoxic T lymphocyte antigen (zytotoxisches T-Lymphozytenantigen)
CXCR3 CXC-Motiv-Chemokinrezeptor 3
d.h. das heißt
dDZ dermale dendritische Zelle
DLQI engl.: Dermatology Life Quality Index (Dermatologischer Lebensqualitäts-Index)
DMF Dimethylfumarat
DMSO Dimethylsufoxid
DNA engl.: deoxyribonucleic acid (Desoxyribonukleinsäure)
DZ dendritische Zelle
ELISA engl.: enzyme-linked immunosorbent assay (enzymgekoppelter
Immunadsorptionstest)
ELISPOT engl.: enzyme-linked immunospot assay (enzymgekoppelter Immunospottest)
et al. lat.: et alii (und andere)
etc. lat.: et cetera (und die übrigen)
FA engl.: fumaric acid (Fumarsäure)
FAE engl.: fumaric acid esters (Fumarsäureester)
FCS engl.: fetal calf serum (fötales Kälberserum)
Foxp3 engl.: transcription factor forkhead box protein P3 (Transkriptionsfaktor Forkhead-
Box-Protein P3)
I
Abkürzungsverzeichnis
g a) Erdbeschleunigung (9,81 m/s2)
b) Gramm
GSH Glutathion
HLA engl.: human leucocyte antigen (humanes Leukozytenantigen)
HO-1 Hämoxygenase-1
HRP engl.: horseradish peroxidase (Meerrettich-Peroxidase)
ICAM engl.: intercellular adhesion molecule (interzelluläres Zelladhäsionsmolekül)
IFN-γ Interferon-γ
IL Interleukin
k kilo
Kap. Kapitel
l Liter
LSM Lymphozyten-Separationsmedium
m a) milli (x10-3)
b) Meter
c) Monat
M Molar
MHC engl.: major histocompatibility complex (Hauptgewebeverträglichkeitskomplex /
Haupthistokompatibilitätskomplex)
min Minute(n)
MMF Monomethylfumarat
mRNA engl.: messenger ribonucleic acid (Boten-Ribonukleinsäure)
n a) engl.: number (Anzahl)
b) nano (x10-9)
NET engl.: neutrophil extracellular traps (Netzwerke extrazellulärer Fasern in
neutrophilen Granulozyten)
NF-kB engl.: nuclear factor “kappa-light-chain-enhancer“ of activated B cells
NK-Zelle natürliche Killerzelle
p Irrtumswahrscheinlichkeit
PASI engl.: Psoriasis Area and Severity Index (Index zur Bestimmung des Schweregrades
von Psoriasis)
PBMC engl.: peripheral blood mononuclear cells (periphere mononukleäre Blutzellen)
PBS engl.: phosphate buffered saline (phosphatgepufferte Salzlösung)
pDZ plasmazytoide dendritische Zelle
ppDZ plasmazytoide prädendritische Zelle
PGA engl.: physician global assessment (ärztliche Gesamtbeurteilung der
Krankheitsaktivität)
PHA Phytohämagglutinin
PHS engl.: pooled human serum (gepooltes Humanserum)
PML progressive multifokale Leukenzephalopathie
II
Abkürzungsverzeichnis
PsA Psoriasis-Arthritis
PSORS 1 engl.: psoriasis suspectibility 1 locus (Psoriasis-Suszeptibilitäts-Locus 1)
PVDF Polyvinylidenfluorid
RORγt engl.: retinoic acid receptor-related-orphan-receptor-gamma t
Rpm engl.: rounds per minute (Umdrehungen pro Minute)
s.o. siehe oben
s.u. siehe unten
SCID engl.: severe combined immunodeficiency (schwerer kombinierter Immundefekt)
sog. sogenannt
STAT engl.: signal transducer and activator of transcription (Signalüberträger und
Transkriptionsaktivator)
Tab. Tabelle
TCR engl.: T cell receptor (T-Zellrezeptor)
TGF engl.: transforming growth factor (transformierender Wachstumsfaktor)
Th-Zelle T-Helferzelle
TLR engl: toll-like receptor (Toll-artiger Rezeptor)
TNF Tumornekrosefaktor
TNFSF15 engl.: TNF superfamily member 15 (Mitglied der TNF-Superfamilie 15)
Treg-Zelle regulatorische T-Zelle
U engl.: units (Einheiten)
u.a. unter anderem
vgl. vergleiche
z.B. zum Beispiel
μ mikro (x10-6)
Fremdsprachige Bezeichnungen (z.B. Eigennamen) sind in der vorliegenden Abfassung kursiv dargestellt.
III
Abbildungsverzeichnis
Abbildungsverzeichnis
Einleitung
Abb. 1: Psoriasis vulgaris und Psoriasis guttata ................................................................................................... 2
Abb. 2: Trigger für das Auftreten von Psoriasis .................................................................................................... 4
Abb. 3: Übersicht über die Differenzierung naiver Th0-Zellen. ...................................................................... 6
Abb. 4: Pathophysiologische Mechanismen bei der Entstehung von Psoriasis ....................................11
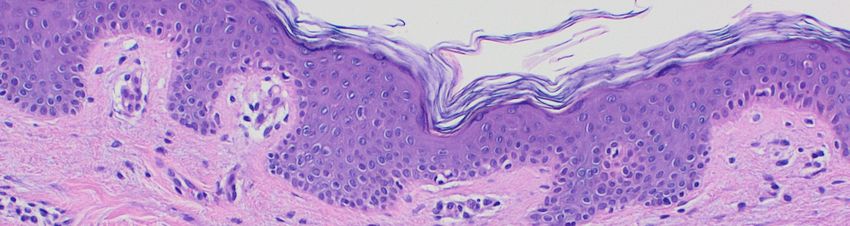
Abb. 5: Histopathologische Veränderungen bei Psoriasis……………………………………………………………12
Abb. 6: Therapiealgorithmus nach geltender S3-Leitlinie ..............................................................................15
Methoden
Abb. 7: Darstellung der Schichtung nach Zellseparation ...............................................................................24
Abb. 8: Schematische Darstellung des ELISPOT-Verfahrens .........................................................................26
Abb. 9: Box-Whisker-Plot ............................................................................................................................................29
Ergebnisse
Abb. 10: Entwicklung der Krankheitsschwere unter FAE-Therapie .............................................................30
Abb. 11: Vergleich zwischen Psoriasispatienten und gesunden Kontrollen ...........................................32
Abb. 12: Vergleich zwischen zytokinsezernierenden Zellen nach Stimulation
mit anti-CD3/CD28-beads und PHA......................................................................................................33
Abb. 13: Stimulationsabhängige Frequenzverläufe von zytokinsezernierenden Zellen
unter FAE-Therapie ......................................................................................................................................35
Abb. 14: Frequenzverläufe nach Stimulation mit anti-CD3/CD28-beads unter FAE-Therapie. .......37
Abb. 15: Konzentrationsabhängige LL37-spezifische T-Zellsubpopulationen .......................................39
Abb. 16: In vitro Effekte von Dimethylfumarat nach Stimulation mit anti-CD3/CD28-beads ..........41
Abb. 17: Konzentrationsabhängige Effekte von Dimethylfumarat .............................................................42
Abb. 18: In vitro Effekte von Monomethylfumarat nach Stimulation mit anti-CD3/CD28-beads ..43
Abb. 19: In vitro Effekte von Fumarsäure nach Stimulation mit anti-CD3/CD28-beads.....................44
Abb. 20: In vitro Effekte von Dimethylsulfoxid nach Stimulation mit anti-CD3/CD28-beads ..........45
IV
Tabellenverzeichnis
Tabellenverzeichnis
Einleitung
Tab. 1: Psoriasis-Phänomene .....................................................................................................................................12
Methoden
Tab. 2: Demografische Daten der Patientenkohorte ........................................................................................21
Tab. 3: Kontraindikationen gegen die Anwendung von Fumarsäureestern bei Psoriasis .................22
Tab. 4: Zusammensetzung von Fumaderm ..........................................................................................................23
Tab. 5: Dosierungsschema der Fumarsäureester-Therapie ...........................................................................23
Tab. 6: Physician Global Assessment .......................................................................................................................23
Anhang
Tab. S1: Psoriasis Area and Severity Index .............................................................................................................61
Tab. S2: Kostenvergleich zwischen Dimethylfumarat und repräsentativen Biologika ........................61
Tab. S3: Literaturrecherche zu ex vivo Effekten einer FAE-Therapie auf T-Zellen .................................62
Tab. S4: Literaturrecherche zu in vitro Effekten von Dimethylfumarat......................................................62
Tab. S5: Literaturrecherche zu in vitro Effekten von Monomethylfumarat ..............................................62
VZusammenfassung
Zusammenfassung
Bei Psoriasis handelt es sich um eine häufige, typischerweise chronisch verlaufende, entzündliche
Hauterkrankung, die circa 2% der Gesamtbevölkerung betrifft. Die Hautveränderungen sind durch
erythematöse, scharf begrenzte Plaques oder Papeln gekennzeichnet, die von silbrig-weißen
Schuppen bedeckt sind. Während die Erkrankung in der Regel nicht lebensbedrohlich ist, sticht
eine hohe psychische Belastung der Patienten durch Stigmatisierung hervor. Da sich die
Symptome mithilfe einer topischen Therapie oft nicht ausreichend kontrollieren lassen, werden
zunehmend systemische Medikamente verordnet. In Deutschland gehören Fumarsäureester (FAE)
zu den häufigsten verschriebenen First-Line-Systemtherapeutika für die Behandlung der Psoriasis.
Bisherige Forschungsergebnisse zu den Pathomechanismen der Psoriasis deuten auf
immunologische Prozesse, insbesondere auf ein Mitwirken von T-Zellen, hin. Obwohl FAE mit dem
Präparat Fumaderm bereits 1994 zur Behandlung der moderaten bis schweren Psoriasis
zugelassen wurden, konnte der Einfluss des Hauptwirkstoffs Dimethylfumarat (DMF) und seines
in vivo Metaboliten Monomethylfumarat (MMF) auf verschiedene T-Zellpopulationen nicht
vollständig geklärt werden.
Mithilfe der vorliegenden Studie wurden daher die Effekte von FAE auf T-Zellen bei Patienten mit
Psoriasis mithilfe des ELISPOT-Verfahrens (enzyme-linked immunospot assay), einer äußerst
sensitiven Methode zur Bestimmung von Zellfrequenzen anhand ihrer Zytokinsekretion,
weitergehend untersucht. Hierfür wurde bei 15 Patienten mit Psoriasis eine FAE-Therapie
eingeleitet und über einen Beobachtungszeitraum von bis zu 12 Wochen die Frequenzen von T-
Helfer (Th) 1-, Th17-, Th2- und regulatorischer T(reg)-Zellen anhand ihrer Markerzytokine IFN-γ-,
IL-17-, IL-5- und IL-10 in ex vivo aktivierten peripheren mononukleären Blutzellen (PBMC)
gemessen. Die FAE-Therapie führte innerhalb dieses Zeitraums bei der Mehrheit der untersuchten
Patienten zu einer Symptomverbesserung mit Verringerung des fünfstufigen Physician Global
Assessment (PGA). Die Therapie wurde frühzeitig von einem Rückgang der zytokinsezernierenden
T-Zellen begleitet. So ergab die vorliegende Studie einen signifikanten Abfall von allen
untersuchten T-Zellpopulationen nach Stimulation mit T-Zell-spezifischen anti-CD3/CD28-beads
sowie für sämtliche mit dem Mitogen Phytohämagglutinin (PHA) stimulierten PBMC in Ausnahme
der IL-10-sezernierenden Zellen. Es wurde insbesondere ein Abfall der IFN-γ-sezernierenden Th1-
Zellen beobachtet. Eine Induktion von immunsuppressiv wirksamen, IL-10-sezernierenden T-
Zellen unter der Therapie mit FAE wurde ex vivo nicht nachgewiesen. Um den Einfluss des
jeweiligen Wirkstoffs auf die verschiedenen T-Zellsubpopulationen zu testen, wurde zusätzlich zu
den longitudinal erhobenen ex vivo Daten PBMC von Patienten und gesunden Kontrollen in vitro
entweder mit DMF, MMF oder Fumarsäure (FA) behandelt. Im Unterschied zu MMF und FA zeigte
dabei DMF in beiden Kohorten einen signifikanten, dosisabhängigen Abfall sämtlicher
VIZusammenfassung
untersuchter T-Zellfrequenzen. Diese Ergebnisse verdeutlichen weiteren Forschungsbedarf
angesichts des ungeklärten Widerspruchs zwischen dem rasanten Metabolismus von DMF zu
MMF in vivo und den ausgeprägten in vitro Effekten von DMF, welche die Wirkung des in vivo
aktiven MMF deutlich übertreffen.
Ein weiteres Ziel dieser Arbeit bestand in dem Vergleich der Frequenzen von
zytokinsezernierenden T-Zellen zwischen der Patienten- und der Kontrollkohorte. Während nach
antigenunspezifischer Stimulation (anti-CD3, PHA und anti-CD3/CD28-beads) keine signifikanten
Unterschiede detektiert werden konnten, wurde nach Verwendung des potenziellen Psoriasis-
spezifischen Autoantigens LL37 ausschließlich in der Patientenkohorte eine Induktion pro-
inflammatorischer T-Zellen beobachtet. Mithilfe des ELISPOT-Verfahrens wurden bei 37,5% der
untersuchten Psoriasispatienten LL37-spezifische T-Zellen detektiert. In der Kontrollkohorte
(gesunde Probanden) wurde hingegen ausschließlich eine anti-inflammatorische IL-10-Antwort
beobachtet. Eine Subanalyse unter den LL37-Respondern der Patientenkohorte zeigte zudem
Hinweise auf eine ausgeprägtere Krankheitsschwere der Psoriasis gegenüber den Patienten ohne
LL37-spezifische T-Zellen (LL37-Non-Responder). Die Rolle von LL37 als ein Autoantigen in einer
Subgruppe von Patienten mit Psoriasis konnte somit bestätigt werden.
Zusammengefasst zeigen die Ergebnisse der vorliegenden Studie, dass FAE frühzeitig zu einer
Reduktion verschiedener T-Zellsubpopulationen im peripheren Blut führen. Die Daten deuten
darauf hin, dass in vivo insbesondere die Abnahme von Th-1- und Th-17-Zellen und weniger eine
Induktion von IL-5- oder IL-10-sezernierenden T-Zellen für die klinische Verbesserung durch eine
FAE-Therapie verantwortlich ist. Anhand der in vitro Daten lässt sich ein Rückgang dieser T-
Zellsubpopulationen eher auf DMF als auf MMF zurückführen.
VIISummary
Summary
Psoriasis is a common, chronic inflammatory skin disease typically appearing as erythematous
plaques or papules covered with silvery-white scales. Skin changes are normally not life-
threatening, nevertheless they are stigmatizing and can put a heavy burden on the patients’ lives.
Since sole topical treatment is often not successful, systemic medication is increasingly applied.
In Germany, fumaric acid esters (FAE) are the most commonly prescribed first-line systemic
treatment for moderate to severe psoriasis.
The development of psoriasis appears to be of an immunological origin. Distinct changes on the
T-cellular level have been reported for psoriasis. Although the licenced FAE formulation
Fumaderm is approved since 1994, the impact of the main active substance dimethyl fumarate
(DMF) and its metabolite monomethyl fumarate (MMF) is still not fully elucidated. Thus, the aim
of the present study was to investigate the effects of FAE on distinct T cell subsets in patients with
psoriasis using the highly sensitive ELISPOT analysis.
For this purpose, 15 patients were treated with FAE and based on the secretion of their marker
cytokines IFN-γ-, IL-17-, IL-5- and IL-10 after ex vivo activation of peripheral blood mononuclear
cells, T helper (Th1), Th17, Th2 and regulatory T cells were followed over a period of up to 12
weeks. FAE-therapy led to an improvement of clinical symptoms in most patients based on the
Physician Global Assessment (PGA). The early treatment process was accompanied by a decrease
of T cells. A significant reduction was determined for all T cell subsets after stimulation of PBMC
with T cell-specific anti-CD3/CD28 beads and the mitogen phytohemagglutinin (PHA), except for
PHA-stimulated IL-10-secreting cells. Interestingly, especially IFN-γ-secreting Th1 cells were
reduced, while FAE did not seem to induce immunosuppressive IL-10-secreting T cells in vivo. In
addition to the ex vivo data during the early course of treatment, in vitro experiments with DMF,
MMF and fumaric acid (FA) were performed with PBMC of patients and healthy controls. Here,
DMF, but neither MMF nor FA, led to a significant, dose-dependent reduction of all examined T
cell frequencies. The discrepancy between the rapid metabolism of DMF in vivo and its distinct
effects in vitro, which exceed those of the in vivo active MMF, underlines the need for further
research.
Another aim of the study was to compare the frequencies of cytokine-secreting cells in patients
and healthy controls. Whereas no differences were detected after non-specific cell stimulation
(anti-CD3, PHA und anti-CD3/CD28-beads), stimulation with the putative psoriasis-specific
autoantigen LL37 induced proinflammatory T cells exclusively in the patient cohort. Performing
the ELISPOT assay, LL37-specific T cells were found in 37.5% of the examined patients with
psoriasis (LL37-responders). In the control cohort only an anti-inflammatory IL-10-response to
VIIISummary
LL37 was observed. Furthermore, a tendency towards more severe psoriasis was found in the LL37-
responders in comparison to the non-responders, thus confirming the role of LL37 as an
autoantigen at least in a subset of patients with psoriasis.
Taken together, the results of this study indicate an early reduction of T cells during FAE-
treatment. Moreover, the data suggest that the clinical efficacy of FAE in the treatment of psoriasis
is mediated by a decrease in Th1- and Th17 cells, and not by an induction of IL-5- and IL-10-
secreting T cells. Based on the in vitro data, the observed reduction in T cell frequencies might
rather be caused by DMF than its metabolite MMF.
IXEinleitung
1 Einleitung
1.1 Psoriasis
Die ersten Berichte über Psoriasis gab es bereits im alten Griechenland (Cowden & van Voorhees,
2008). Zunächst zu den lepromatösen Hauterkrankungen gezählt, wurde sie zu Beginn des 19.
Jahrhunderts durch den englischen Arzt Willan (1757-1812) als eine eigenständige
Krankheitsentität beschrieben (Cowden & van Voorhees, 2008). Heute ist bekannt, dass es sich
bei Psoriasis um eine häufige, im Gegensatz zur Lepra nicht infektiös übertragbare, chronisch-
entzündliche Hauterkrankung handelt. Bislang liegt weder eine klare Kenntnis über die Entstehung
dieser stigmatisierenden Erkrankung noch über eine definitive Heilung vor. Der Fokus der
vorliegenden Abfassung liegt auf pathophysiologischen Prozessen auf T-Zellebene als
Angriffspunkt für therapeutisch eingesetzte Fumarsäureester (FAE).
1.1.1 Klinik der Psoriasis vulgaris
Psoriatische Effloreszenzen bestehen typischerweise aus scharf begrenzten,
erythematosquamösen Papeln und Plaques. Sie können lokalisiert oder generalisiert auftreten.
Anhand von Aussehen und Lokalisation der Hautveränderungen, dem zeitlichen Verlauf und einer
möglichen Gelenkmanifestation werden einzelne Psoriasis-Subtypen differenziert, wobei Psoriasis
vulgaris die häufigste Erkrankungsform darstellt (vgl. Abb. 1). Prädilektionsstellen der Psoriasis
vulgaris sind die Kopfhaut, der äußere Gehörgang, der Rumpf, die Extremitäten-Streckseiten, der
Bauchnabel sowie die Sakralregion. Psoriasis vulgaris kann in Typ-I-Psoriasis (Manifestationsalter
zwischen dem 20. und 30. Lebensjahr, ausgeprägter Schweregrad, familiäre Häufung, genetische
Prädisposition) und in den etwas selteneren Typ II (Manifestationsalter zwischen dem 50. und 60.
Lebensjahr, geringer Schweregrad, keine familiäre Häufung, nur geringe genetische
Prädisposition) eingeteilt werden (Moll & Jung, 2016; Altmeyer, 2012). Nagelveränderungen wie
Ölflecken oder Tüpfelnägeln treten bei bis zu 50% der Patienten auf (Moll & Jung, 2016). In
schweren Fällen kann es zu einer Onychodystrophie oder Onycholyse kommen. 70% der Patienten
mit Psoriasis leiden unter einem Juckreiz der Hautläsionen (Reich et al., 2016).
1.1.2 Psoriasis guttata
Psoriasis guttata ist eine exanthematische Manifestationsform der Psoriasis mit kleineren,
tropfenartig geformten, schuppenden Papeln und Plaques (vgl. Abb. 1). Sie tritt häufig als
Erstmanifestation einer Psoriasis oder in einem akuten Krankheitsschub auf (Altmeyer et al., 2012).
1Einleitung
Charakteristischerweise sind Kinder und Jugendliche nach einer vorausgegangenen Infektion mit
Streptokokken betroffen. Bei Konfluenz der Hautläsionen ist ein Übergang in eine Psoriasis
nummularis mit münzgroßen Plaques oder in eine Psoriasis vulgaris möglich (Braun-Falco et al.,
2005).
Abb. 1: Psoriasis vulgaris und Psoriasis guttata. Dargestellt ist auf der linken Seite ein Patient (PSO8) mit
Psoriasis vulgaris und auf der rechten Seite ein Patient (PSO11) mit Psoriasis guttata, die durch ihre kleineren
tropfenartigen, erythematosquamösen Plaques und Papeln charakterisiert ist (Universitätsklinik für
Dermatologie und Allergologie Marburg; vgl. Tab. 2).
1.1.3 Psoriasis pustulosa palmaris et plantaris
Bei der Psoriasis pustulosa palmaris et plantaris kommt es an Handflächen und Fußsohlen zur
Ausbildung von Hyperkeratosen und kleinen sterilen Pusteln (Moll & Jung, 2016). Ein chronischer
Krankheitsverlauf und ein erschwertes Therapieansprechen sind typisch für diesen Psoriasis-
Subtyp (Miceli & Schmieder, 2017). Die Hautveränderungen können sehr schmerzhaft sein und zu
ausgeprägten Funktionseinschränkungen bis hin zur Immobilisierung führen.
1.1.4 Erythrodermia psoriatica
Eine potenziell lebensgefährliche Maximalvariante der Psoriasis stellt die psoriatrische
Erythrodermie dar. Hierbei ist nahezu das gesamte Integument erythematosquamös infiltriert und
es kann zu gravierenden Komplikationen wie einer kreislaufrelevanten Dehydratation,
Regulationsstörungen der Körpertemperatur und zu Elektrolytentgleisungen kommen (Moll &
Jung, 2016; Altmeyer, 2012).
2Einleitung
1.1.5 Psoriasis arthropathica
Bei ca. 20% der Patienten mit Psoriasis liegt eine Psoriasis arthropathica (Psoriasis-Arthritis, PsA)
vor, die neben fakultativen Hautveränderungen durch eine Gelenkbeteiligung charakterisiert ist
(Amherd-Hoekstra et al., 2010). Typischerweise sind dabei die distalen Interphalangealgelenke
von einer destruierend-proliferierenden Gelenkentzündung mitbetroffen (Amherd-Hoekstra et al.,
2010). In Abgrenzung zu anderen Formen der Arthritis ist für die PsA der Befall eines gesamten
Fingerstrahls (Daktylitis) typisch. Seltener kommt es zu einem Befall der Wirbelsäule und der
Iliosakralgelenke (Amherd-Hoekstra et al., 2010). Klinisch können Arthralgien,
Bewegungseinschränkungen und Deformationen imponieren.
1.2 Epidemiologie
Mit einer Prävalenz von 2% weltweit gehört Psoriasis zu den häufigsten Hauterkrankungen
(Boehncke & Schön, 2015). Im internationalen Vergleich bestehen allerdings große Unterschiede.
Beispielsweise ist die Prävalenz in Norwegen mit 8,5% deutlich höher als in Deutschland mit 2,5%
(Michalek et al., 2016a) oder dem Vereinigten Königreich von Großbritannien und Nordirland mit
1,3% Erkrankten (Springate et al., 2017). Asiaten und Afrikaner sind dagegen besonders selten von
Psoriasis betroffen (Michalek et al., 2016b). Psoriasis kann in jedem Lebensalter auftreten und
betrifft geringfügig häufiger das männliche Geschlecht (Moll & Jung, 2016). Zwillings- und
Familienstudien weisen auf eine starke genetische Komponente bei der Krankheitsentstehung hin
(Nestle et al., 2009). So zeigten empirische Daten ein 7-fach erhöhtes Risiko zur Ausbildung einer
Psoriasis, wenn bereits ein Elternteil betroffen ist, und ein 16-fach erhöhtes Risiko, wenn beide
Elternteile an Psoriasis erkrankt sind (Swanbeck et al., 1997). Die Anzahl von Psoriasis-assoziierten
Genen ist mittlerweile auf 36 gestiegen (Brück et al., 2018), wovon viele mit dem Immunsystem in
Verbindung gebracht werden (Harden et al., 2015). Insbesondere der psoriasis suspectibility locus
(PSORS) 1, der sich im Bereich des Klasse-I-Haupthistokompatibilitätskomplexes (major
histocompatibility complex, MHC) befindet und auch das „Psoriasis-Allel“ HLA-Cw6 enthält, scheint
für bis zu 50% der Krankheitsvererbung verantwortlich zu sein (Nestle et al., 2009; Elder, 2006).
Erst vor kurzem wurde in Assoziationsstudien mit TNFSF15 ein neuer genetischer
Suszeptibilitätslocus ausgemacht (Brück et al., 2018). Neben einer genetischen Prädisposition
können auch Umweltfaktoren das Erkrankungsrisiko für Psoriasis beeinflussen. Studien haben eine
Risikoerhöhung durch Nikotin- und Alkoholabusus, Übergewicht, psychischen Stress und Infekte
gezeigt (Naldi et al., 2005). Die Einnahme von Medikamenten wie Lithium oder β-Blockern,
mechanische Hautirritationen und saisonale Einflüsse stellen weitere Einflussfaktoren dar (Abb. 2;
Boehncke & Schön, 2015). Eine gestiegene Lebenserwartung und ein besseres Bewusstsein für die
3Einleitung
Diagnose Psoriasis scheinen in den letzten Jahren zu einem Prävalenzanstieg beigetragen zu
haben (Springate et al., 2017). Aufgrund langer Krankheitsdauer, eines ausgeprägten
Leidensdrucks und erheblicher Therapiekosten stellt Psoriasis eine weltweite Herausforderung dar
(Michalek et al., 2016b). Während die Symptome in der Regel nicht lebensbedrohlich sind, ist die
psychische Belastung der Patienten durch Stigmatisierung und Diskriminierung hervorstechend
(Schmid-Ott et al., 2003). Inzwischen wird zusätzlich davon ausgegangen, dass Psoriasis eine
Erkrankung ist, die „unter die Haut geht“ (Bowcock & Krueger, 2005) und Eigenschaften einer
entzündlichen Systemerkrankung aufweist (Conrad & Flatz, 2013; Boehncke & Schön, 2015). Im
Sinne des sog. „psoriatischen Marsches“ kann die chronische Entzündungsreaktion bei Psoriasis
zum Auftreten von Insulin-Resistenz und Atherosklerose, zentralen Risikofaktoren für Herzinfarkte
und Schlaganfälle, führen (Koo et al., 2017; Gerdes et al., 2016; Boehncke & Schön, 2015; Hu &
Lan, 2017). Die häufige Komorbidität mit Depressionen, selbstverletzendem Verhalten,
kardiovaskulären Ereignissen, Adipositas und Diabetes mellitus kann die erhöhte
Gesamtmortalität von Patienten mit Psoriasis erklären (Horreau et al., 2013; Springate et al., 2017;
Egeberg et al., 2016; Koo et al., 2017).
Abb. 2: Trigger für das Auftreten von Psoriasis. Dargestellt sind typische auslösende bzw.
befundverschlechternde Faktoren bei Psoriasis.
4Einleitung
1.3 Pathophysiologie der Psoriasis
Während Psoriasis bis in die 1990er aufgrund der ausgeprägten epidermalen Hyperplasie vor
allem für eine Keratinozyten-assoziierte Erkrankung gehalten wurde (Bowcock & Krueger, 2005),
konnte inzwischen durch intensive Forschung ein besseres Verständnis von der Pathogenese
erreicht und die wesentliche Bedeutung immunologischer Mechanismen erkannt werden.
Hinweise auf immunologische Pathomechanismen bei Psoriasis finden sich u.a. durch die
folgenden Erkenntnisse: die erhöhte Anzahl an Immunzellen in psoriatischen Hautläsionen (Bos
et al., 1983), die Krankheitsübertragbarkeit nach allogener Knochenmarkstransplantation
(Gardembas-Pain, 1990) und das gute Therapieansprechen auf immunmodulierende
Medikamente wie Methotrexat oder Ciclosporin A (Griffiths et al., 2000). Den T-Zellen kommt
dabei eine Schlüsselrolle bei der Induktion von psoriasisartigen Hautveränderungen zu (Boehncke
& Schön, 2007, 2015; Breban et al., 1996). Dies wurde experimentell u.a. anhand von severe
combined immunodeficiency (SCID)-Mäusen, bei denen ein Mangel bzw. eine Dysfunktion der
Lymphozyten besteht (Boehncke & Schön, 2007; Bosma et al., 1983), nachvollzogen. So konnten
mithilfe dieses Mausmodells in nicht-läsionalen Hauttransplantaten von Patienten mit Psoriasis
durch intradermale Injektion von autologen T-Zellen der Spender psoriasisartige
Hautveränderungen hervorgerufen werden (Wrone-Smith & Nickoloff, 1996).
1.3.1 T-Zellen
T-Zellen gehören dem adaptiven Immunsystem an. Sie entstammen dem Knochenmark, reifen im
Thymus und gelangen schließlich in die Blutbahn und die Lymphknoten (Murphy et al., 2009).
Anhand ihrer Oberflächenproteine werden T-Zellen in cluster of differentiation (CD)8+, sog.
zytotoxische T-Zellen, und in CD4+ T-Zellen, sog. T-Helferzellen (Th-Zellen), differenziert
(Golubovskaya & Wu, 2016), wobei im Folgenden der Fokus auf den Th-Zellen liegt.
Für die Polarisierung einer naiven Th0-Zelle in eine Th-Effektorzelle sind mindestens drei Signale
erforderlich (Rink et al., 2015). Signal 1 stellt die antigenspezifische Erkennung von dem MHC-
Peptid-Komplex auf Antigen-präsentierenden Zellen (antigen-presenting cell; APC) durch den T-
Zell-Rezeptor (T cell receptor, TCR) naiver CD4+ Th0-Zellen dar (Rink et al., 2015). Neben dem TCR
besitzen T-Zellen einen CD3-Komplex, der für die Signalgebung nach Antigenbindung an den TCR
unerlässlich ist (Murphy et al., 2009). Für die weitere T-Zellaktvierung (Signal 2) sind
kostimulatorische Moleküle erforderlich. Dabei kommt es zu einer Interaktion zwischen B7-
Molekülen auf der Oberfläche von aktivierten dendritischen Zellen (DZ) und dem CD28-Molekül
der naiven Th0-Zelle, wodurch diese in die G1-Phase des Zellzyklus eintritt (Hof et al., 2014; Rink
et al., 2015). Unter dem Einfluss von Interleukin (IL)-2 kommt es innerhalb mehrerer Tage zu einer
starken Zellproliferation, bei der Tausende klonale T-Zellen entstehen (Rink et al., 2015). In
5Einleitung
Abhängigkeit ihres dargebotenen Antigens, freigesetzter Entzündungsmediatoren und anderen
Faktoren des Mikromilieus können DZ die Polarisierung zu unterschiedlichen Th-Subpopulationen
induzieren (Abb. 3; Rink et al., 2015). Die Art dieses Mikromilieus, das auch autokrin von der
proliferierenden Th-Zelle beeinflusst wird, stellt das Signal 3 dar (Rink et al., 2015). So begünstigt
bspw. eine Umgebung aus IL-12 und IFN-γ die Entwicklung von Th1-Zellen (Golubovskaya & Wu,
2016). Neben Th1-Zellen werden anhand spezifischer Transkriptionsfaktoren und daraus
resultierenden charakteristischen Zytokinmustern weitere Subgruppen von Th-Zellen wie Th2-,
Th9-, Th17-, Th22- und regulatorische T-Zellen (Treg-Zellen) differenziert (Golubovskaya & Wu,
2016; Rink et al., 2015). Durch die verschiedenen Effektorzytokine der einzelnen Subpopulationen
wird die Abwehr eines breiten Erregerspektrums gewährleistet (Rink et al., 2015).
Abb. 3: Übersicht über die Differenzierung naiver Th0-Zellen zu ausgewählten Th-Subpopulationen.
Dargestellt ist eine naive Th0-Zelle, die sich abhängig vom vorherrschenden Zytokinmilieu zu einer Th1-,
Th2-, Th17- oder Treg-Zelle polarisiert. Auf der rechten Seite sind Effektorzytokine der jeweiligen Th-
Subpopulationen abgebildet (modifiziert nach Golubovskaya & Wu, 2016).
1.3.1.1 Th1-Zellen und ihre Effektorzytokine
Th1-Zellen verfügen über ausgeprägte pro-inflammatorische Wirkungen und dienen der Abwehr
von extra- und intrazellulären Erregern wie Tuberkelbakterien (Hof et al., 2014; Rink et al., 2015).
Sie sezernieren IFN-γ, IL-2, IL-3, granulocyte macrophage colony-stimulating factor (GM-CSF) und
Tumornekrosefaktor-α (TNF-α; Golubovskaya & Wu, 2016; Rink et al., 2015). Über diese
Effektorzytokine haben sie Einfluss auf die Differenzierung antigenspezifischer B-Zellen, den
Isotypwechsel bei der Antikörperproduktion, die Apoptoseinduktion, die Stimulation von
Endothelzellen und die Attraktion von Leukozyten aus der Blutbahn (Hof et al., 2014). Weiterhin
können Th1-Zellen Makrophagen zytokinvermittelt oder über CD40-Interaktion zur Produktion
6Einleitung
antibakterieller, toxischer und pro-inflammatorischer Substanzen stimulieren (Hof et al., 2014).
Diese begünstigen ihrerseits über die Produktion von IL-12 eine T-Zelldifferenzierung in Richtung
von Th1-Zellen (vgl. Abb. 3; Hof et al., 2014). Th1-Zellen spielen eine wichtige Rolle bei
Autoimmunerkrankungen und Hypersensitivitätsreaktionen vom verzögerten Typ (Rink et al.,
2015). Auch Psoriasis ist mit einer pathologischen Th1-Zelldifferenzierung assoziiert (Ghoreschi et
al., 2011; Meissner et al., 2012). In der Epidermis bewirkt die Expression von IFN-γ im transgenen
Mausmodell neben einer Hyperproliferation auch eine veränderte Zelldifferenzierung mit
Induktion von MHC-II-Molekülen und interzellulären Zelladhäsionsmolekülen (ICAMs) sowie die
Bildung von vergrößerten dermalen Kapillaren (Boehncke & Schön, 2007; Carroll et al., 1997).
1.3.1.2 Th2-Zellen und ihre Effektorzytokine
Th2-Zellen sezernieren die Zytokine IL-4, IL-5, IL-6, IL-10, IL-13 und TGF-β (transforming growth
factor-β) und spielen insbesondere bei der humoralen Erregerabwehr eine bedeutende Rolle
(Golubovskaya & Wu, 2016; Rink et al., 2015; Hof et al., 2014). Sie differenzieren sich in einer IL-4-
reichen Umgebung, die bspw. durch eosinophile Granulozyten bei der Abwehr von Parasiten
erzeugt wird (Abb. 3; Hof et al., 2014). Ähnlich wie Th1-Zellen unterstützen Th2-Zellen die
Differenzierung naiver B-Zellen und den Isotypwechsel (Murphy et al., 2009). Th2-Zellen fördern
dabei die Produktion von IgA- und IgE-Antikörpern durch B-Zellen. Allergien vom Soforttyp,
Asthma bronchiale sowie die atopische Dermatitis sind mit einem pathologischen Überwiegen
von Th2-Zellen assoziiert (Murphy et al., 2009; Rink et al., 2015; Gatzka & Scharffetter-Kochanek,
2015). Bei Psoriasis wird hingegen ein Mangel an Th2-Zellen mit einer resultierenden Dysbalance
zugunsten pro-inflammatorischer Th1-Zellen angenommen (Quaglino et al., 2011; Meissner et al.,
2012).
1.3.1.3 Th17-Zellen und ihre Effektorzytokine
Bei Th17-Zellen handelt es sich um eine inhomogene Gruppe, die nach sezerniertem
Zytokinmuster in IL-17 einfach-positive und IL-17A/IL-22 sowie IL-17/IFN-γ doppelt-positive
Th17-Zellen unterschieden werden kann (Zhou et al., 2014). Th17-Zellen entwickeln sich aus
naiven CD4+ T-Zellen unter dem Einfluss der Zytokine IL-1, IL-6, IL-23 und TGF-β (Abb. 3; Zhou et
al., 2014). IL-17 fördert die Produktion von pro-inflammatorischen Zytokinen (IL-1, IL-6, TNF-α)
und antimikrobiellen Peptiden (AMP), mobilisiert neutrophile Granulozyten und verstärkt die
Migration von T-Zellen in die Haut (Lowes et al., 2008; Arican et al., 2005; Golubovskaya & Wu,
2016; Speeckaert et al., 2016). Insbesondere bei der Abwehr von Bakterien und Pilzen kommt den
Th17-Zellen damit eine große Bedeutung zu (Speeckaert et al., 2016). Eine Vermehrung von Th17-
7Einleitung
Zellen wurde im peripheren Blut (Kagami et al., 2010) sowie in Hautläsionen von Patienten mit
Psoriasis beschrieben, wo die Effektorzytokine IL-17 und IL-22 die Keratinozyten-Proliferation und
die Entzündungsprozesse fördern (Lowes et al., 2008; Nestle et al., 2009). Auch bei anderen
autoimmunvermittelten Erkrankungen, wie der rheumatoiden Arthritis und der Spondylitis
ankylosans, wurde ein Mitwirken von Th17-Zellen gezeigt (Speeckaert et al., 2016).
1.3.1.4 Regulatorische T-Zellen und ihre Effektorzytokine
Die Subgruppe der Treg-Zellen weist ausgeprägte entzündungshemmendende und
immunregulierende Eigenschaften auf (Golubovskaya & Wu, 2016). Treg-Zellen sind durch die
Expression des Transkriptionsfaktors forkhead box protein 3 (Foxp3) gekennzeichnet (Ziegler &
Buckner, 2009). Es wird unterschieden in natürliche Treg-Zellen (nTreg-Zellen), die sich im Thymus
entwickeln, und induzierte Treg-Zellen (iTreg-Zellen), welche in der Peripherie gebildet werden
(Ziegler & Buckner, 2009). Ein charakteristisches Merkmal für Treg-Zellen ist die Sekretion des
anti-inflammatorischen Zytokins IL-10 (Golubovskaya & Wu, 2016). IL-10 hemmt z. B. die Bildung
pro-inflammatorischer Zytokine wie IFN-γ, IL-2, IL-3, GM-CSF und TNF-α (Owczarczyk-Saczonek
et al., 2018; Saxena et al., 2015), inhibiert die Th1-Immunantwort durch Suppression der IL-12-
Synthese (Owczarczyk-Saczonek et al., 2018) und unterdrückt direkt das Wachstum von Th17-
Zellen und APC (Saxena et al., 2015; Owczarczyk-Saczonek et al., 2018). Damit schützt IL-10 vor
unkontrollierten Entzündungsprozessen und Gewebeschäden (Bovenschen et al., 2011).
Wesentliche Produzenten von IL-10 sind neben Treg-Zellen Makrophagen, DZ und B-
Lymphozyten (Owczarczyk-Saczonek et al., 2018). Bei Psoriasis scheint die Funktion von Treg-
Zellen vermindert zu sein (Sugiyama et al., 2005) und somit ein Ungleichgewicht zwischen dem
anti-inflammatorischen IL-10 und pro-inflammatorischen Zytokinen zu bestehen (Mussi et al.,
1994; Saxena et al., 2015). In einem Tiermodell mit CD18-Knockout-Mäusen wurde zudem eine
Dysfunktion von Treg-Zellen mit dem Auftreten psoriasisähnlicher Hautveränderungen assoziiert
(Wang et al., 2008).
1.3.1.5 Das Th1/Th2-Zell-Paradigma und T-Zellplastizität
Das Konzept einer Dichotomie der Th-Zellen in Th1- und in Th2-Zellen wurde 1986 eingeführt
und bestand für ungefähr 20 Jahre (Mosmann et al., 2005; Rink et al., 2015). Nach diesem Modell
führt eine Th1-Zell-dominierte Immunantwort zu Hypersensitivitätsreaktionen vom verzögerten
Typ und autoimmunvermittelten Erkrankungen, wohingegen eine Th2-Zell-dominierte
Immunantwort mit Erkrankungen des atopischen Formenkreises in Verbindung gebracht wird
(Steinman, 2007). Th1- und Th2-Zellen werden hierbei als Gegenspieler betrachtet und eine
8Einleitung
reziproke Hemmung angenommen (Steinman, 2007; Hof et al., 2014). Das Th1/Th2-Zell-
Paradigma musste allerdings erweitert werden, weil einerseits einige experimentelle Befunde
anhand dieses Modells nicht erklärt werden konnten und andererseits mit Treg- und Th17-Zellen
neue T-Zell-Subpopulationen entdeckt wurden, die wesentliche Rollen bei Entzündungsprozessen
spielen (Steinman, 2007; Zhou et al., 2014). Weiterhin ist mittlerweile das Konzept der T-
Zellplastizität anerkannt, wonach bereits ausdifferenzierte T-Zellsubpopulationen
kontextabhängig den Phänotyp anderer Subpopulationen mit der jeweiligen Zytokinsekretion
annehmen können (Gatzka & Scharffetter-Kochanek, 2015; Tartar et al., 2009; Kannan et al., 2019).
Während die Th1/Th2-Zell-Differenzierung aufgrund von epigenetischen Veränderungen, die
jeweils die Expression der anderen Th-Subpopulation unterdrücken, relativ stabil erscheint, wirkt
die Differenzierung zu Treg- und Th17-Zellen unbeständig (Gatzka & Scharffetter-Kochanek,
2015; Ziegler & Buckner, 2009). Sie erfolgt u.a. über die zytokinabhängige Bildung von
Transkriptionsfaktoren wie Foxp3 und signal transducer and activator of transcription (STAT)5 für
die Differenzierung von Treg-Zellen sowie von RORγt und STAT3 für Th17-Zellen (Gatzka &
Scharffetter-Kochanek, 2015). In einem entzündlichen Zytokinmilieu aus IL-1β, IL-21 und IL-23
können Treg-Zellen zu pro-inflammatorischen Foxp3+IL-17A-produzierenden Zellen
differenzieren, die Th17-Zellen ähneln und auch in Hautläsionen von Patienten mit Psoriasis
nachgewiesen wurden (Bovenschen et al., 2011). Bei Psoriasis scheint die T-Zellplastizität somit
einen bedeutenden Mechanismus zur Chronifizierung der Erkrankung darzustellen (Gatzka &
Scharffetter-Kochanek, 2015).
1.3.2 Das antimikrobielle Peptid LL37
AMP kommen insbesondere in entzündetem Gewebe vor, wo sie die Erregerabwehr mithilfe ihrer
bakteriziden, fungiziden, antiviralen und immunmodulatorischen Eigenschaften unterstützen
(Vandamme et al., 2012). Ein wichtiges AMP ist LL37, dessen Name bereits den Aufbau aus 37
Aminosäuren beginnend mit Leucin offenbart (Vandamme et al., 2012). LL37 wird vorrangig von
Keratinozyten und neutrophilen Granulozyten, aber auch von Epithelzellen, Monozyten,
Makrophagen, DZ, natürlichen Killerzellen (NK), Mastzellen und einigen Lymphozyten gebildet
(Vandamme et al., 2012; Fuentes-Duculan et al., 2017; Agerberth et al., 2000). Es wird nach
unkontrolliertem Zelltod freigesetzt und bei Entzündungen aktiv sezerniert. Durch Zellschäden
kann negativ geladene DNA freigesetzt werden, die mit dem positiv geladenen LL37 im
Extrazellularraum Komplexe bildet (Abb. 4; Vandamme et al., 2012). Diese Komplexe können über
die toll-like Rezeptoren (TLR) 7/8/9 oder zytosolische DNA-Sensoren in Zellen eindringen und
intrazelluläre TLR von DZ aktivieren, wodurch die Immuntoleranz gegenüber körpereigener DNA
und RNA gestört wird und diese zu pro-inflammatorischen Stimuli werden (Vandamme et al.,
9Einleitung
2012; Lande et al., 2007; Nestle et al., 2009). Darüber hinaus postulierten Lande et al. (2014), dass
LL37 ein mögliches Autoantigen bei Psoriasis darstellt.
1.3.3 Mechanismen bei der Entstehung von psoriatischen Hautläsionen
Als Pathomechanismen bei Psoriasis sind die Interaktion zwischen innatem (DZ, Makrophagen,
Mastzellen, Granulozyten) und adaptivem Immunsystem (vor allem Th1- und Th17-Zellen),
Schlüssel-Zytokine wie IFN-γ sowie Wechselwirkungen mit kutanen Zellen wesentlich (Boehncke
& Schön, 2015). Die Entstehung einer neuen Hautläsion kann bei Vorliegen von genetischen
Risiko- und Umweltfaktoren bspw. durch mechanische Reizung initiiert werden. Diese führt über
Zytokinfreisetzung (IL-1β, TNF-α, IL-16) und Bildung von o.g. LL37-DNA-Komplexen zur
Aktivierung von plasmazytoiden DZ (pDZ; Abb. 4; Nestle et al., 2009; Boehncke & Schön, 2007;
Lande et al., 2014). Diese führen wiederum zur Produktion von Interferon und zur Reifung von
dermalen DZ (dDZ), die in drainierende Lymphknoten einwandern und über die Sekretion von IL-
12 und IL-23 die Differenzierung naiver Th0-Zellen zu pro-inflammatorischen Th1- und Th17-
Zellen anregen (Abb. 4; Nestle et al., 2009; Vandamme et al., 2012). Hiermit stellen die DZ eine
wichtige Verbindung zu dem adaptiven Immunsystem bei Psoriasis her (Nestle et al., 2009;
Vandamme et al., 2012). Die Th-Zellen rezirkulieren im Blut und wandern gesteuert durch
Chemokin-Rezeptoren wie CCR4, CCR6 und CXCR3 in die Haut ein (Abb. 4; Nestle et al., 2009).
Der CC-chemokine ligand 27 (CCL27) und andere Chemokine werden insbesondere von
Keratinozyten sezerniert (Bowcock & Krueger, 2005). Nach der Initiationsphase kann die Epidermis
schließlich von T-Zellen über die Expression von Integrin-α1β1 erreicht werden (Conrad & Flatz,
2013). Weitere Zellpopulationen wie Makrophagen, DZ und neutrophile Granulozyten kommen
hinzu und bilden das psoriatische Hautinfiltrat (van de Kerkhof, 2007; Bowcock & Krueger, 2005;
Nestle et al., 2009; Conrad & Flatz, 2013). Die Th1- und Th17-Zytokine führen zu einer epithelialen
Dysfunktion mit Hyperproliferation der Keratinozyten (Lande et al., 2014; Lowes et al., 2008;
Conrad & Flatz, 2013). Mit der Zunahme der epidermalen Hyperplasie und Leukozyten-Infiltration
geht schließlich eine akute Läsion in eine chronische über, deren zelluläre Veränderungen sich
über parakrine Interaktionen verstärken und zunehmend selbst unterhalten können (Bowcock &
Krueger, 2005). Die persistierende T-Zell-Stimulation durch aktivierte DZ und epidermale Antigene
(Conrad & Flatz, 2013) sowie die Produktion der Schlüssel-Zytokine IFN-γ, IL-12, IL-17, IL-22, IL-
23 und TNF-α durch die Zellen des psoriatischen Hautinfiltrats erscheinen hierbei maßgeblich
(Nestle et al., 2009; Boehncke & Schön, 2007; Conrad & Flatz, 2013). Es existieren zahlreiche
positive Rückkopplungsschleifen mit Keratinozyten, Makrophagen, Endothelzellen und
Fibroblasten, die ihrerseits zum Umbau des Gewebes beitragen (Nestle et al., 2009; Hof et al.,
2014). So werden Keratinozyten von Entzündungsmediatoren zur Proliferation und der Bildung
10Einleitung
von Chemokinen, S100-Proteinen und AMP wie LL37 angeregt, was eine weitere Aktivierung der
Immunprozesse begünstigt (Nestle et al., 2009; Conrad & Flatz, 2013). Makrophagen fördern über
die Sekretion von Chemokinen wie CCL19 die Bildung von perivaskulären T-Zell-Infiltraten (Nestle
et al., 2009) und unterstützen eine Th1-Zelldifferenzierung (Hof et al., 2014). Als ein
pathophysiologisches Korrelat für das Köbner-Phänomen bei Psoriasis wird die
Rückkopplungsschleife mit IL-1β diskutiert, das bei Hautschädigung freigesetzt wird und Th17-
Zellen zur Produktion von IL-17 anregen kann, welches wiederum zu einer Erhöhung von IL-1β
beiträgt (Speeckaert et al., 2016).
Abb. 4: Schematische Darstellung pathophysiologischer Mechanismen bei der Entstehung von
Psoriasis. Durch Triggerfaktoren kommt es zur Freisetzung von LL37 und Eigen-DNA, deren Komplexe
plasmazytoide dendritische Zellen (pDZ) aktivieren, die sich zu dermalen dendritischen Zellen (dDZ)
entwickeln. Über das periphere Blut gelangen die DZ in die drainierenden Lymphknoten und unterstützen
dort zytokingesteuert die Differenzierung von Th0- in Th1- und Th17-Zellen. Diese rezirkulieren im Blut,
wandern in die Haut ein und sezernieren – mutmaßlich nach Präsentation von Autoantigenen – Zytokine, die
Keratinozyten aktivieren und zur Bildung weiterer pro-inflammatorischer Zytokine, Chemokine und
antimikrobieller Peptide (AMP) anregen. Es kommt somit zu einem selbstverstärkenden Kreislauf pro-
inflammatorischer Signale, der auch weitere Zelltypen wie Makrophagen (MΦ), Fibroblasten und
Endothelzellen einbezieht (hier für eine bessere Übersichtlichkeit nicht dargestellt) und klinisch u.a. als
ausgeprägte epitheliale Hyperproliferation imponiert (modifiziert nach Nestle et al., 2009 sowie Conrad &
Flatz, 2013).
11Sie können auch lesen