SILVA FERA Wissenschaftliche Nachrichten aus dem Wildnisgebiet Dürrenstein - Band 6 - Wildnisgebiet Dürrenstein
←
→
Transkription von Seiteninhalten
Wenn Ihr Browser die Seite nicht korrekt rendert, bitte, lesen Sie den Inhalt der Seite unten

ISSN 2227-3387
Band 6
SILVA FERA
Wissenschaftliche Nachrichten aus dem
Wildnisgebiet Dürrenstein
2 Silva Fera, Bd. 6/Mai 2017
Silva Fera, Band 6, 2017
Erschienen: Mai 2017
Silva Fera ist die wissenschaftliche Zeitschrift der Schutzgebietsverwaltung Wildnisgebiet Dürrenstein.
Sie dient der Veröffentlichung neuer Erkenntnisse und Forschungsergebnisse aus dem Wildnisgebiet Dürrenstein.
ISSN:2227-3387
Herausgeber und Medieninhaber:
Schutzgebietsverwaltung Wildnisgebiet Dürrenstein, Brandstatt 61, A – 3270 Scheibbs
www.wildnisgebiet.at, office@wildnisgebiet.at
Für den Inhalt verantwortlich:
DI Dr. Christoph Leditznig
Redaktion:
Dr. Sabine Fischer
Druck:
Druckerei Queiser, 3270 Scheibbs
Kosten:
30,- €/Heft
Titelfoto: Eisenhuthummel (Bombus gerstaeckeri), © Theo Kust
Seite 3: Der Rothausbach im Großen Urwald, © Martin Hartmann
Seite 5: Sterndolde (Astrantia major) mit Krabbenspinne und Graubindigem Mohrenfalter (Erebia aethiops), © Werner Gamerith
Seite 6: Großer Urwald (Rothwald), © Martin Hartmann
Seite 97: „Fenster zum All“ (Blick aus der Legsteinhütte am Dürrenstein auf den Nachthimmel), © Markus Reithofer
Seite 98: „Stargazers“ bei der Legsteinhütte, © Markus Reithofer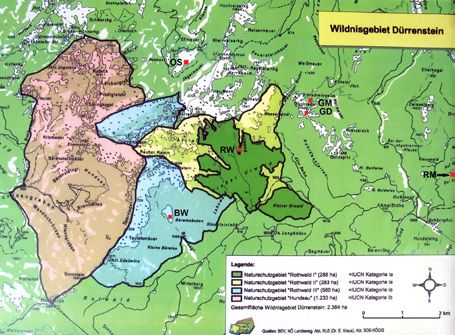
Silva Fera, Bd. 6/Mai 2017 3

4 Silva Fera, Bd. 6/Mai 2017
Inhalt
....................................................................................................................................................................................................................................................... Seite
Vorwort ................................................................................................................................................................................................................................................5
Brande A., K. Splechtna & S. Zerbe:
Pollenanalytische Untersuchungen zur Vegetationsgeschichte im Wildnisgebiet Dürrenstein ...............................................................................................................7
Kohl I. & C. Leditznig:
Ein Vergleich der unterschiedlichen Telemetrie-Systeme im Rahmen der Wiederansiedlung von Habichtskäuzen (Strix uralensis) im Wildnisgebiet Dürrenstein
in den Jahren 2009 bis 2016 ...............................................................................................................................................................................................................23
Kust T.:
Entomofaunistische Untersuchungen im Wildnisgebiet Dürrenstein.
2. Teil: Hautflügler (Hymenoptera): Pflanzenwespen (Symphyta) und Hummeln (Bombus) .................................................................................................................59
Wuchterl G.:
Das Licht der Nacht über dem Wildnisgebiet Dürrenstein ..................................................................................................................................................................77
Publikationsrichtlinien........................................................................................................................................................................................................................99
Silva Fera, Bd. 6/Mai 2017 5
UÊ *
Ê i>>ÞÌÃV iÊ1ÌiÀÃÕV Õ}iÊÛÊ ÀiÀ- und das Pflanzengift Glyphosat, die aufgrund ihrer
nen aus Mooren östlich des DürrensteinMassivs Toxizität niemals hätten zugelassen werden dürfen.
geben Aufschluss über mehrere tausend Jahre Die finanzstarken Saatgut- und Chemiekonzer-
Vegetationsentwicklung in unserem Gebiet. ne konnten bisher – unterstützt von der österrei-
chischen Landwirtschaftskammer – ein dringend
UÊ
Ê V ÌÊ > ÀiÊ ÌÀ}Ê Ê 7i`iÀ>Ãi`Õ}Ã- notwendiges Totalverbot verhindern, die EU-weite
projekt der Habichtskäuze lassen Vergleiche von Wiederzulassung von Glyphosat zeichnet sich ge-
unterschiedlichen Telemetriemethoden zu. Die- rade ab.
sen wertvollen Erkenntnissen wünschen wir eine
weite Verbreitung, damit das Rad (bei ähnlichen Trotz der fortschreitenden Umweltzerstörung und
Aufgaben) nicht immer neu erfunden werden der zermürbenden Wachstumshörigkeit der kon-
muss. ventionellen Wirtschaft, dürfen wir uns nicht von
Ohnmacht und Verzweiflung lähmen lassen, denn
UÊ 1
Ê ÃiÀÊ7ÃÃiÊØLiÀÊ`>ÃÊÀÌiëiÌÀÕÊiÀÜiÌiÀÌÊ jeder einzelne Mensch, jeder kleine Schritt zu ei-
diesmal ein Beitrag über die Pflanzenwespen nem bewussteren Leben zählt. Engagement macht
– ursprüngliche Hautflügler, die meist an be- Mut und es gibt so viel, was wir tun können.
stimmte Wirtspflanzen gebunden sind – und die
Hummelfauna. Ihre
Sabine Fischer
UÊ -
Ê iÌÊ Óä£äÊ ÜÀ`Ê LiÊ />}Ê Õ`Ê >V ÌÊ `iÊ L>- (Redaktion)
strahlung über der Legsteinhütte am Dürren-
stein gemessen. Die Werte sind von natürlichen
Lichtverhältnissen nicht unterscheidbar, d.h. das
Vorwort Gebiet ist eines der letzten mit Naturnacht in
Mitteleuropa!
Liebe Leserinnen und Leser!
Die weitgehend intakte Natur im Wildnisgebiet ist
Die bisher erschienenen fünf Bände unserer wissen- natürlich ein Ansporn für unsere Schutzbestrebun-
schaftlichen Nachrichten zeigen die Vielfältigkeit gen, denn je seltener etwas wird, desto wertvoller
der Forschungs- und Artenschutzprojekte, die dank wird es.
fruchtbarer Kooperationen und dem Engagement
vieler (natur)begeisterter Personen im und rund um Als naturliebender Mensch bin ich erschüttert, dass
das Wildnisgebiet Dürrenstein umgesetzt werden der Wert unversehrter Ökosysteme, die atemberau-
können. bende Schönheit unserer Welt und ihrer Lebensfor-
men, sowie unsere Abhängigkeit von der Biosphäre,
Im vorliegenden sechsten Band wird diese Multidis- der breiten Masse und vor allem den politischen
ziplinarität wieder deutlich. Sie finden Beiträge aus Entscheidungsträgern immer noch nicht ausrei-
der Palynologie, dem angewandten Artenschutz, chend ins Bewusstsein gedrungen ist. Beispielhaft
der Astronomie und der Entomologie: dafür sind z.B. die Neonicotinoide (Insektizide)
6 Silva Fera, Bd. 6/Mai 2017

Silva Fera, Bd. 6/Mai 2017 7
Pollenanalytische Untersuchungen Abstract kultur, Wien) an. Sie stützen sich somit auf deren
umfangreiche Erfahrungen in der Ausarbeitung
zur Vegetationsgeschichte Pollen analyses were conducted at four sites east of und Interpretation zahlreicher Pollendiagramme
im Wildnisgebiet Dürrenstein the Dürrenstein mountain massif, between 1100 zur natürlichen und anthropogenen Waldgeschich-
and 1400 m elevation and within the natural spru- te im mittleren und östlichen Alpenraum unter
Arthur Brande, Karl Splechtna ce-fir-beech belt. The pollen diagrams display that den wechselnden klima- und nutzungsgeschichtli-
the climax forest was formed 4000 years ago (Rot- chen Bedingungen der Nacheiszeit (ursprüngliche
& Stefan Zerbe
mösel), followed by a regional beech dominance be- Waldzusammensetzung und ihre Entstehung, Hö-
fore the onset of human impact. A first regression henstufendifferenzierung, Standorteigenschaften,
Zusammenfassung
of beech due to cuttings is found during Roman klimatische und anthropogene Schwankungen der
times (Bärwies), resp. medieval and modern times Waldgrenzen, Walddegradation durch Almweide
Die vier pollenanalytischen Untersuchungspunkte
(Grubwies sites, Rotmösel). Subsequently, a very und Holznutzung u.a.). Für die hier vorgelegten
liegen östlich des Dürrenstein-Massivs zwischen
strong regression of fir occurs diachronically at the Pollendiagramme Bärwies aus dem Wildnisgebiet
1100 und 1400 m Höhe in der Fichten-Tannen-
sites with a difference of about 1200 years and up Dürrenstein und Grubwies sowie Rotmösel aus den
Buchenstufe. Nach der im Diagramm Rotmösel
to the 18th/19th century (Rotmösel). Mountain östlich anschließenden Teilflächen konnten daher
erfassten Ausbildung des Klimaxwaldes vor 4000
pasture after and together with forest clearance is viele Einzelangaben zum Zeigerwert der Pollenty-
Jahren zeigen alle Diagramme eine regionale Bu-
documented by indicators such as juniper, aconi- pen und zur Pollenherkunft aus den verschiedenen
chendominanz. Ein erster rodungsbedingter Bu-
te, other herbs and grasses. The expansion of the Höhenstufen und Standorten der Arbeit von Kral &
chenrückgang liegt in der Römerzeit (Bärwies)
mountain pine shrub belt depending on site and Mayer (1968) mit den dort veröffentlichten Pollen-
bzw. in Mittelalter und Neuzeit (Grubwies Moor
time conditions is shown by the increase of pine diagrammen Rotmösel und Rothwald und weiteren
und Doline, Rotmösel). Auch der nachfolgende,
pollen amount. The origin and development of the Publikationen der Autoren entnommen werden. Die
sehr einschneidende Tannenrückgang ist an den
investigated mires is found to be topo-climatogene obere Hälfte der Torfabfolge im Rotmösel wurde
Untersuchungspunkten zeitverschoben mit einer
(Rotmösel), soligene (Bärwies) or topo-soligene bereits von Kral pollenanalytisch untersucht (Kral
Differenz von 1200 Jahren bis in das 18./19. Jahr-
(Grubwies). It covers a time span from the Subbo- & Mayer 1968), allerdings ohne 14C-Daten zur zeit-
hundert (Rotmösel). Die mit der Waldauflichtung
real to the Older Subatlantic chronozone, i.e. the lichen Gliederung der Vegetationsentwicklung. Ein
einhergehende Almweidenutzung mit Weidezei-
late warmperiod of the Holocene to the older part Vergleich beider Pollendiagramme und ihrer Unter-
gern wie Wacholder, Eisenhut, weiteren Kräutern
of the post-warm period. The results do not point schiede findet sich bei Brande et al. (2002).
sowie Gräsern kommt je nach Standort zeitlich und
to an anthropogenic origin of mire formation at all.
örtlich unterschiedlich zum Ausdruck, ebenso die
Im Folgenden werden die vier Pollendiagramme
Ausdehnung der Latschengebüsche. Die Entste-
1. Einleitung erstmals vollständig und im wechselseitigen Zusam-
hung und Entwicklung der untersuchten Moore ist
menhang vorgestellt. Die Untersuchung Rotmösel
topo-klimatogen (Rotmösel), soligen (Bärwies) und
Die pollenanalytischen Untersuchungen an der TU ist im Rahmen einer Übersicht zur Waldentwick-
topo-soligen (Grubwies) im Zeitraum vom Subbo-
Berlin waren Teil des niederösterreichischen EU- lung des Nordostalpenraumes gesondert behandelt
real (Späte Wärmezeit) bis Subatlantikum (Ältere
Life-Projektes „Wildnisgebiet Dürrenstein“ (1997 worden (Brande et al. 2002), außerdem eine kurze
Nachwärmezeit). Eine anthropogene Entstehung
- 2001; vgl. Ripl et al. 2004). Sie schließen metho- Übersicht im Endbericht des Forschungsprojektes
der Moore ist auszuschließen.
disch und inhaltlich an frühere Arbeiten und Pu- (Brande 2004a) und ein Vergleich mit den Untersu-
blikationen vor allem von Prof. Dr. Friedrich Kral chungen der Bärwies im Arbeitskreis Vegetations-
in Zusammenarbeit mit Prof. Dr. Hannes Mayer geschichte (Tagung Innsbruck, Brande & Splecht-
(Institut für Waldbau der Universität für Boden- na 2005).
8 Silva Fera, Bd. 6/Mai 2017
2. Das Untersuchungsgebiet lomitrücken auf. Die Buche dominiert hier, zeigt
aber auf den verhagerten Rücken oftmals geringen
Zur Auswertung der Pollendiagramme sind insbe- Wuchs. Bergahorn, Fichte und Wald-Kiefer können
sondere die Nutzungsgeschichte und die aktuellen beigemischt sein.
Vegetationstypen des Gebietes von Bedeutung.
Eine kurze Übersicht zur Geschichte der Waldnut- In den Schuttkegeln unterhalb von Felswänden sto-
zung im Dürrensteingebiet, besonders zum Wald- cken kleinflächige Schlucht- bzw. Hangwälder mit
verlust zwischen 1782 und 1875, geben Splechtna Bergahorn, Esche und Bergulme. Splechtna & Kö-
& Splechtna (2016) mit Hinweisen auf die zugrun- nig (2001) schätzen die Natürlichkeit der Bestände
de liegenden detaillierten Publikationen. aufgrund der schwierigen Nutzungsbedingungen
als sehr hoch ein.
Die Vegetation im Dürrensteingebiet zeichnet sich
durch Wälder aus, die im Kernbereich (Urwald Subalpine bodensaure Fichtenwälder sind im Gebiet
Rothwald) nie bewirtschaftet wurden. In den relativ selten, da die Fichten-Tannen-Buchenwälder
Randbereichen (Erweiterungsgebieten) wurde die weit hinaufreichen, an der Baumgrenze aber durch
Bewirtschaftung eingestellt. Oberhalb der Wald- Weidenutzung zurückgedrängt wurden. Sie gedei-
grenze gedeihen Latschengebüsche und alpine hen auf steilen und grobblockigen Rücken und Rip-
Matten. Der folgenden Darstellung liegen die um- pen mit Säure- und Rohhumuszeigern in der Kraut-
fassenden Studien von Zukrigl et al. (1963), Mayer schicht. Neben Fichte kommen vereinzelt Lärche,
et al. (1979), Mayer & Neumann (1981) und Carli Buche und Bergahorn vor. Eine besondere Ausprä-
(2007) über den Urwald Rothwald und die Kartie- gung zeigen die „Hochstauden-Fichtenwälder“ auf
rung der FFH-Lebensraumtypen von Splechtna & den Hochplateaus (Mucina et al. 1993). Dieser lich-
König (2001) zugrunde. Die hier vorgestellte Über- te Waldtyp ist vielfach durch Beweidung und da-
sicht ist die gekürzte Fassung einer pflanzensoziolo- mit einen Nährstoffeintrag überformt, so dass die
gisch-vegetationsökologischen Beschreibung. Rasenschmiele (Deschampsia cespitosa) flächig auf-
tritt, z.B. auf der Grubwiesalm. Bei fast allen Fich-
Bergwälder Abb. 1: Fichten-Tannen-Buchenwald im Großen Ur- tenwäldern im Gebiet muss von einer historischen
Ein Großteil des Dürrensteingebietes bzw. des wald am Rothausbach. (Foto: A. Brande) bzw. aktuellen Beweidung ausgegangen werden.
Urwaldes Rothwald wird von unterschiedlichen Kleinflächig treten der Blockfichtenwald und auf
standörtlichen Ausprägungen des für den nordöst- gung in der Kraut- und Strauchschicht. Vereinzelt Waldgrenzstandorten wie Felskanten der Kalkfels-
lichen Alpenraum typischen Fichten-Tannen-Bu- sind Bergahorn und Bergulme beigemischt. Auffäl- Fichtenwald auf.
chenwalds bestockt (Abb. 1). lig ist in den Urwaldresten das weitgehende Fehlen
von Pionierbaumarten wie Birke und Eberesche. Subalpine Latschengebüsche
Die drei Assoziationen auf Kalkbraunlehmbö- An hochmontanen, schneereichen Hängen gedei- Die Gipfel- und Hangbereiche oberhalb der Baum-
den unterscheiden sich deutlich in der Kraut- und hen Buchenwälder des Aceri-Fagetum. Die Fichte grenze werden von drei Assoziationen der Lat-
Moosschicht (Willner & Grabherr 2007) und wei- ist hier kaum vertreten. schengebüsche bestockt (Abb. 2). Sie sind durch die
sen die Hauptbaumarten Fichte, Tanne und Buche historische Weidenutzung und damit einhergehen-
in verschiedenen Mischungsanteilen auf. Die Buche Der Orchideen-Kalk-Buchenwald tritt nur auf be- de Rodungen begünstigt und über ihre natürliche
ist die dominierende Baumart mit starker Verjün- sonders flachgründigen und sonnenseitigen Do- Verbreitung hinaus im Gebiet ausgedehnt.
Silva Fera, Bd. 6/Mai 2017 9 Abb. 2: Latschengebüsch, Kalkfels-Fichtenwald und Seggenrasen am Sperriedel und Abb. 3: Almweide auf der Grubwies. (Foto: A. Brande) Dürrensteingipfel. (Foto: F. Haudum, Wilhering) Alpine und subalpine Rasen hervor und verweisen auf die Notwendigkeit einer grenze oder als Ersatzgesellschaft des Hochstau- Oberhalb der Waldgrenze und durch historische Beweidung bzw. Mahd zum Erhalt. den-Fichten-Tannen-Buchenwaldes. Auf ehemalige Weidenutzung nach vorheriger Rodung in die un- Beweidung und Wildeinfluss weisen Aconitum na- teren Höhenlagen verschoben, finden sich alpine Schutthalden- und Felsspaltenvegetation pellus (Abb. 4), Deschampsia cespitosa und Urtica und subalpine Kalkrasen, vor allem Horstseggen-, An südexponierten Hängen im Rothwald wer- dioica hin. Heute stellen diese hochproduktiven Rostseggen- und Polsterseggen-Rasen, wobei ers- den Schutthalden von thermophilen Schwalben- Vegetationsbestände Wildäsungsflächen dar, und es tere den flächenmäßig größten Anteil einnehmen. wurzfluren bewachsen. Auch findet sich bis in die muss – auch wegen der langen Schneebedeckung – Splechtna & König (2001) stellen auch die Hoch- Gipfelregion der Kalk- und Dolomitfelsen häufig von einer stark verzögerten Wiederbewaldung aus- grasfluren zu den Kalkrasen und verweisen auf die Felsspaltenvegetation unterschiedlicher Zusam- gegangen werden. floristisch-soziologische Verwandtschaft zum Rost- mensetzung. An schattigen Felsen gedeihen Strei- seggenrasen. fen- und Blasenfarnbestände. Moore Montane Borstgrasrasen mit einer artenreichen Vermoorte Bereiche finden sich vor allem im Gro- Begleitflora (Abb. 3) sind auf der Grubwiesalm Hochstaudenfluren ßen Bärwiesboden, einem letztglazialen Karboden, anzutreffen (Grabherr & Zechmeister 1998). Mit Feuchte, nitrophile Hochstaudenfluren wachsen in dessen Abfluss durch Moränenwälle blockiert ist, so Blick auf die Artenvielfalt heben Splechtna & Kö- Talbereichen der Bäche, entlang von Waldrändern, dass es besonders im niederschlagsreichen Sommer nig (2001) die Bedeutung dieses Vegetationstyps auf durchrieselten Schutthalden oberhalb der Wald- zu Wasserüberstauungen kommt. Im Untergrund

10 Silva Fera, Bd. 6/Mai 2017
Abb. 4: Hochstaudenflur mit Blauem Eisenhut auf
dem Großen Bärwiesboden (Foto: A. Brande)
verhindert eine Lehmdecke die Versickerung. In
dem vorherrschenden Schnabelseggenried kommen
aufgrund der kleinflächig stark wechselnden Hy-
drologie eine Vielzahl verschiedener Pflanzenge-
sellschaften bzw. unterschiedliche Dominanzbe- Abb. 5: Lage der pollenanalytischen Untersuchungspunkte im Dürrensteingebiet. Kartengrundlage: BEV, NÖ
stände vor. Landesreg. Abt. RU5, Abt. BD5-NÖGIS. RM – Rotmösel, BW – Großer Bärwiesboden, GM – Grubwiesalm
Moor, GD – Grubwiesalm Doline, RW – Rothwald I, II und RM – Rotmösel: Kral & Mayer (1968), OS – Lunzer
Weitere Vegetationstypen Obersee: Krisai (1993). Hinweis der Redaktion: Die Karte zeigt die Ausdehnung des Wildnisgebiets Dürrenstein
Neben den beschriebenen naturnahen bis anthro- in den Gründungsjahren 2001/2002, nicht den aktuellen Stand.
pogenen Pflanzengesellschaften mit z.T. hohem
Naturschutzwert finden sich weitere Vegetations- 3. Die Pollendiagramme der montan-subalpinen Fichten-Tannen-Buchen-
typen wie Fichtenaufforstungen, Schlagfluren (vgl. stufe zwischen 1100 und 1400 m Höhe östlich,
Zukrigl et al. 1963), Bergweiden (z.B. Grubwies- Die pollenanalytischen Untersuchungspunkte (Abb. südlich und nördlich des Großen und Kleinen Ur-
alm) und die Vegetation in den Gewässern. 5, Tab. 1) liegen östlich des Dürrenstein-Massivs in waldes (Rothwald).Silva Fera, Bd. 6/Mai 2017 11
(Lang 1994) gewon- folgende Pollensummen zur Prozentberechnung
nen (Abb. 6). Die Bohr- herangezogen: 1. Gehölze (BP excl. Corylus) = 100
punkte sind aufgrund %, mit eingeschaltetem Teildiagramm der Klimax-
von vorher durchge- baumarten Picea/Abies/Fagus = 100 %, 2. Krautige
führten Sondierungen incl. Zwergsträucher (Cerealia bis Farnpflanzen)
nach möglichst hoher in der Gesamtsumme (incl. Gehölze, excl. Moose
Torfmächtigkeit und etc.) = 100 %. In den Diagrammen Grubwies Moor
-qualität ausgewählt und Doline sind Farnpflanzen (Polypodiaceae p.p.)
worden. wegen sehr hoher Werte nicht in der Gesamtsum-
Die Pollenanalysen me enthalten, sondern auf sie bezogen. Schatten-
wurden 1999-2001 risskurven grau über schwarz zeigen die 10-fache
und 2006-2008 am Überhöhung der Prozentwerte. Zur Abschätzung
Institut für Ökologie der relativen Pollendichte/Sedimentvolumen sind
der TU Berlin, Fach- den Proben bei der Aufbereitung im Labor Lycopo-
gebiet Ökosystem- dium-Sporen als Marker der Pollenkonzentration
kunde/Pf lanzenöko- beigegeben. Hohe Lycopodium-Werte verweisen
logie, Arbeitsgruppe demnach auf eine geringe Pollendichte infolge
Historische Ökolo- Waldoffenheit bzw. hoher Torfzuwachsraten. Die
Abb. 6: Torfbohrung im Moor der Grubwiesalm am 21.8.1999. gie/Pollenanalytisches Diagrammgliederung wurde mithilfe lokaler Pol-
(Foto: K. Splechtna) Labor (Leitung A. lenzonen (LPZ) vorgenommen. Die an den vier
Brande) durchge- Bohrprofilen untersuchten chemischen Parameter
3.1 Methodik führt. Die graphische Darstellung der Pollendia- sowie deren Auswertung und Verknüpfung mit
Die Bohrkerne der Torfe wurden am 20.8. (Rot- gramme erfolgte mit dem Programm TGView, den pollenanalytisch-vegetationsgeschichtlichen
mösel, Bärwies) und 21.8.1999 (Grubwies Moor Version 2.0.2 (Grimm 2004). Die Nomenklatur Befunden sind bei Ripl et al. (2004, Kap. 6, S. 41-
und Doline) von A. Brande und W. Ripl jr. mit der Pollentypen folgt Faegri & Iversen (1993) und 44) kurz dargestellt.
dem Eij-kelkamp-Kammerbohrer (System Jowsey) Beug (2004). Die pollenmorphologische Bezeich-
und die Basisschichten mit der Dachnowski-Sonde nung Cerealia steht für Getreide-Pollentypen au- 3.2 Datierungen
ßer Secale (Roggen) Zu den Pollendiagrammen liegen 14C-Daten aus
und Zea (Mais), p.p. dem Leibniz-Labor für Altersbestimmung und Iso-
Tab. 1: Kenndaten zu den pollenanalytischen Untersuchungspunkten
für pro parte (zum topenforschung der Christian-Albrecht-Universität
Moorfläche Höhenlage Tiefe erbohrt Teil) und indet. für Kiel (Dr. Helmut Erlenkeuser) aus den Jahren 2001-
Untersuchungspunkt Abkürzung indeterminata (unbe- 2004 vor (Tab. 2). Für das Rotmösel-Diagramm
(ha) (m ü. NN) (cm)
stimmbar aufgrund wurde von der Torfbasis beginnend eine Datierung
Rotmösel RM 0,9 1140 320
mangelnder Pollener- der Übergänge der lokalen Pollenzonen angestrebt
Großer Bärwies- haltung). Dabei sind (vgl. Tab. 1 bei Brande et al. 2002), für die übri-
BW 4,4 1110 95
boden in Anlehnung und zur gen Diagramme der Beginn der Torfbildung und,
Grubwiesalm Moor GM 0,25 1380 100 Vergleichbarkeit mit soweit bei datierungsfähiger Torfqualität möglich,
den Ergebnissen von der Übergang zu den Pollenzonen der nutzungsin-
Grubwiesalm Doline GD 0,12 1370 75 Kral & Mayer (1968) tensiven Phasen.12 Silva Fera, Bd. 6/Mai 2017
Tab. 2: Radiocarbon-Daten zu den Pollendiagrammen. Pollenstratigraphische Kriterien (Brande): + Anstieg oder Zunahme; – Rückgang; ohne Zeichen: kontinuierliche
Präsenz; kNB: krautige Nutzpflanzen und Begleiter (Cerealia bis Cannabis). 14C-Jahre BP +/- 1 σ: gemessene Radiocarbonjahre vor 1950 n. Chr. mit 1 σ-Standardabweichung.
Die gemessenen δ 13C-Werte schließen einen Hartwassereffekt (Alterung) durch fossiles, gesteinsbürtiges Karbonat aus. cal BC/AD: Kalenderjahre, kalibriert nach Stuiver et
al. (1998) mit 1,65 σ, d.h. 90 % Wahrscheinlichkeit. Jahr n./v. Chr.: Mittelwert aus Spalte 6.
Tiefe Labor Kiel 14
C-Jahre BP δ 13C cal BC/AD
Pollenstratigraphische Kriterien Jahr n./v. Chr.
(cm) Proben-Nr. +/- 1 σ ‰ 1,65 σ, 90 %
Rotmösel
16 - 19 Abies – Fagus – Larix + kNB + KI-4886,01 120 +/- 65 -27,67 1655 - 1955 AD 1805 n.
38 - 44 Picea + Abies – KI-4886,02 1310 +/- 35 -27,07 655 – 775 AD 715 n.
58 - 64 Betula + Pinus + KI-4886,03 1960 +/- 70 -26,56 95 BC - 135 AD 20 n.
91 - 95 Abies Fagus Picea KI-4886,04 2620 +/- 65 -26,03 895 – 595 BC 745 v.
121 - 129 Betula + Pinus + Fagus – KI-4886,05 3050 +/- 70 -25,90 1435 - 1125 BC 1280 v.
162 - 169 Cerealia + KI-4886,06 3230 +/- 70 -25,69 1685 - 1405 BC 1545 v.
192 - 198 Fagus Picea Abies KI-4886,07 3450 +/- 70 -25,51 1895 - 1615 BC 1755 v.
233 - 239 Fagus +Plantago lanceolata + KI-4886,08 3690 +/ 65 -26,26 2275 - 1915 BC 2095 v.
273 - 279 Fagus + Abies Sphagnum-Torf KI-4886,09 3910 +/- 35 -26,31 2475 - 2285 BC 2380 v.
305 - 309 Fagus + Picea – KI-4886,10 4970 +/- 45 -28,10 3915 - 3655 BC 3785 v.
310 - 314 Fagus + Ulmus – KI-4886,11 5220 +/- 80 -27,97 4225 - 3815 BC 4010 v.
Bärwies
47 - 57 Picea Abies Fagus KI-5157,01 1470 +/- 25 -28,43 535 – 645 AD 590 n.
72 - 82 Fagus Picea Abies KI-5157,02 1790 +/- 30 -28,26 125 – 345 AD 235 n.
Grubwies Moor
50 - 55 Fagus Picea Abies KI-5159,01 520 +/- 30 -26,57 1325 - 1445 AD 1385 n.
80 - 85 Fagus Picea Abies KI-5159,02 1090 +/- 25 -27,76 895 - 1025 AD 960 n.
Grubwies Doline
21 - 25 Picea + Abies – KI-5158,01 60 +/- 55 -27,14 1675 - 1945 AD 1800 n.
52 - 59 Fagus Picea Abies KI-5158,02 620 +/- 35 -26,97 1285 - 1405 AD 1345 n.Silva Fera, Bd. 6/Mai 2017 13
3.3 Vegetationsentwicklung bei Kral & Mayer (1968) weitgehend identisch. Die stufen finden sich bei Brande et al. (2002) sowie zu
3.3.1 Rotmösel Autoren haben nur die oberen 1,6 m erbohrt und deren heutiger Verbreitung im Dürrensteingebiet
Grundlage zur Rekonstruktion der Vegetationsge- untersucht (vgl. Brande et al. 2002). im Kap. 2 sowie bei Splechtna & König (2001), zu
schichte im Dürrenstein- und Rothwaldgebiet ist Taxus außerdem bei Brande (2004b).
das Pollendiagramm aus dem Rotmösel bei Neu- In der Zeit zwischen 2000 BC und 700 AD besteht
haus (Abb. 7 und Anhang I). als Klimaxvegetation ein voll entwickelter Fichten- Erste Nachweise von Siedlungs- und Nutzungstä-
Tannen-Buchenwald. Es ist die längste Zeitdauer, tigkeit durch Polleneinwehung aus den Tieflagen
In den 3,2 m mächtigen Torfen des kleinen Hoch- in der dieser regionaltypische Wald im Untersu- sind durch Spitzwegerich (Plantago lanceolata) um
moores in Plateaulage („Auf den Mösern“, sauer- chungsgebiet als „primärer Urwald“ (Kral & May- 2275 - 1915 BC am Übergang von LPZ 2 zu 3
oligotrophes Regenmoor nach Steiner 1992: 457) er 1968) existiert hat. Sein Beginn liegt vor mehr bezeugt (Wende Jungsteinzeit/Bronzezeit), deut-
setzt in einer flachen Karstmulde auf Dachsteinkalk als vier Jahrtausenden am Übergang der LPZ 2 zu lich später als der tatsächliche Beginn von Ackerbau
die Wald- und Moorentwicklung vor mehr als 6000 3 um 2275 - 1915 BC. Die stabilste Phase erleb- und Viehhaltung im Alpenvorland. Auch Getrei-
Jahren ein. Sie erfasst an der Torfbasis die Einwan- te er zwischen 2000 und 1600 BC. Anschließend denachweise (Cerealia) gibt es erst am Übergang 3
derung und Ausbreitung der Buche, während Fich- wechseln Anteile von Fichte, Tanne und Buche mit zu 4a um 1685 - 1405 BC (Mittlere Bronzezeit).
te und Tanne bereits im Gebiet vorhanden sind. einem Fagus- und Abies-Maximum um 1500 bzw. Seit etwa 1300 BC und im Verlauf des LPZ 4a-c
Durch elf 14C-Daten sind sieben Entwicklungs- 1000 BC. Sie sind Ausdruck der Urwalddynamik erfasst ein wechselnder menschlicher Einfluss (Be-
phasen der Vegetation, z. T. weiter untergliedert, und der Sukzessionsvorgänge im Bestandsaufbau tula, Pinus, krautige Kulturpflanzen und -beglei-
bis zum Jahr 1998/99 chronologisch belegt. Der (im Rothwald: Mayer et al. 1979, Schrempf 1986) ter) die weitere Umgebung des Moores (Bronze-,
Bohrpunkt im Zentrum des Moores ist mit dem durch externe und interne Faktoren, u.a. vermutlich Hallstatt-, Römerzeit). Das entspricht im Wesentli-
in Verbindung mit den chen der Besieldungs- und Nutzungsgeschichte im
Klimaschwankungen mittleren und östlichen Alpenraum (Oeggl 2012).
des späten Subbore- Splechtna (2001) weist vor allem auf die keltische
als und des Älteren und die hochmittelalterliche Besiedlung und Wald-
Subatlantikums (Löb- nutzung (überwiegend Rodungen und Umwand-
ben- und Göschenen lung in Almweide) bis in die höheren Berglagen
I-Kaltphase; Kral 1979) hin, die in der LPZ 4b (oberer Teil) und 5 (unterer
sowie der unterschied- Teil) anzusetzen ist. Im Pollendiagramm macht sich
lichen Blühphasen ein Nutzungseinfluss besonders am Übergang 4b
(Pollenproduktion) zu 5 im frühen Mittelalter um 665 - 775 AD mit
bei gleichzeitig ho- einem ersten rodungsbedingten Tannenrückgang
her zeitlicher Auflö- zugunsten der Fichte stärker bemerkbar, wenige
sung der Pollenpro- Jahrhunderte zuvor auch schon mit den Pionierge-
ben aufgrund hoher hölzen Lärche und Grünerle (Larix, Alnus viridis).
Torf bildungsraten. Erst nach 1655 AD erreicht dann das Rodungsge-
Weitere Hinweise zur schehen in der LPZ 6 mit dem massiven Rückgang
Bestandsdynamik der der Tanne, etwas schwächer auch der Buche – pro-
übrigen Gehölze (Be- zentual zugunsten von Kiefer und Birke – die un-
tula bis Taxus) in den mittelbare Umgebung des Moores. Ursache ist vor
Abb. 7: Das Rotmösel mit Lagepunkt der Torfbohrung. (Foto: K. Splechtna) verschiedenen Höhen- allem der Holzeinschlag seit Mitte bis Ende des 18.14 Silva Fera, Bd. 6/Mai 2017
Jahrhunderts (vor allem Holzkohleproduktion zur 3.3.2 Bärwies In dem Pollendiagramm (Anhang II) lassen sich
Eisenverhüttung), gefolgt von Fichtenaufforstungen Der Große Bärwiesboden ist ein Dolinenkessel, zwei Phasen unterscheiden. In der LPZ A herrscht
in der LPZ 7 bis an den Moorrand. Weiderodung dessen Anlage nach Fink (1973) vermutlich eiszeit- der regionaltypische Fichten-Tannen-Buchenwald
am nördlich anschließenden Zwieselberg, die nach lich vorgeprägt wurde. Ein Blockmoränenfeld am mit Fagus-Dominanz. Die örtliche Vegetation am
Splechna (2004) nur wenig älter sein mag, kommt Südrand deutet auf die Entstehung als Kar. Der Untersuchungspunkt besteht aus Nasswiesen mit
hinzu. Weit- und Fernflugpollen unter den Gehölzen Bohrpunkt liegt 60 m SSW der ehemaligen Alm- Mädesüß, Ampfer, Süß- und Sauergräsern (Fili-
und Kräutern treten vermehrt in Erscheinung (vgl. hütte in einem Carex rostrata-Bestand (Abb. 8). pendula, Rumex, Poaceae, Cyperaceae), in der an-
entsprechende Angaben bei Bärwies und Grubwies). Nach eigenen Vorsondierungen und dem aktuellen, fänglichen Vernässungsphase auch mit Wasserpflan-
Örtlich ist im Moorumkreis vorübergehend der pro- kleinräumig stark wechselnden Vegetationsmosaik zen (Potamogeton, Pediastrum) sowie Farnen und
zentuale Pollenniederschlag von Spitzwegerich, Bren- dürfte sich hier die größte Torfmächtigkeit der Sen- Schachtelhalm (Polypodiaceae, Equisetum).
nessel (Urtica) und anderen Kräutern häufiger. Damit ke befinden. Seit dem Älteren Subatlantikum hat
ist am Rotmösel die LPZ 6, d.h. das 19. Jahrhundert, sich um 125 - 345 AD (Tab. 2) auf zeitweilig über- Der abrupte Vegetationswandel an der Grenze
die Periode der stärksten menschlichen Eingriffe. flutetem Lehm ein 0,8 m mächtiger Braunmoos- A/B fand im Jüngeren Subatlantikum deutlich
Die Moorentwicklung mit der Torfbildung als Trä- und Radizellentorf gebildet. nach 535 - 645 AD statt. Da die Grenze A/B mit
ger der vegetationsgeschichtlichen Überlieferung
durchläuft in LPZ 1 und 2 nach Vernässung und
Versumpfung als Nieder- und Übergangsmoor
verschiedene Hochmoorbildungsphasen mit Torf-
moos-Dominanz (Sphagnum) unter Beteiligung
von Wollgras und Moosbeere (Eriophorum vagina-
tum, Vaccinium oxycoccus). Sie spiegelt sich auch im
mooreigenen Pollenniederschlag wider (z.B. Poten-
tilla-Typ, Vaccinium-Typ, Cyperaceae, Poaceae p.p.,
Equisetum- und Sphagnum-Sporen, Amphitrema-
Amöbenschalen). Die maximale Torfbildungsrate
liegt bei 1,2 mm/Jahr in LPZ 3 und 4a, die mi-
nimale bei 0,4 mm/Jahr in LPZ 4c (schwach bzw.
stark zersetzter Sphagnum-Torf). Letztere beruht
möglicherweise auf höheren Temperaturen und/
oder geringeren Niederschlägen zwischen den kli-
matischen Kaltphasen Göschenen I und II (Kral
1979). In LPZ 6 und 7 treten schließlich Sauer- und
Süßgräser am und auf dem Moor stark hervor. Sie
sind mit Trichophorum cespitosum, Carex rostrata,
C. limosa sowie Molinia coerulea auch heute präsent,
dazu Pinus silvestris/mugo mit dem Pollenmaximum
in der jüngsten Probe. Einen näheren Vergleich mit
den nachgewiesenen Pollentypen bietet die Vegeta-
tionskartierung von Kusel-Fetzmann (1981). Abb. 8: Der Große Bärwiesboden vom Sperriedel mit Lagepunkt der Torfbohrung. (Foto: K. Splechtna)Silva Fera, Bd. 6/Mai 2017 15
einem Wechsel der Torfbildung zusammenfällt, stellenweise lehmigen
ist nicht auszuschließen, dass hier eine zeitliche Einschwemmungen
Schichtlücke (Hiatus) vorliegt, bedingt durch die dokumentiert. Somit
instabilen hydrologischen Bedingungen in der Sen- ist diese Dynamik ei-
ke (Quellschüttungen; vgl. Haseke 2016). Sowohl nerseits durch Ände-
ein vorübergehender Stillstand der Torfbildung rungen der Vernässun-
(Austrocknung) als auch eine Abschwemmung gen und Bachläufe mit
(Erosion) der jüngsten bis zu dieser Zeit gebildeten der Quellschüttung
Torfe könnte die Ursache sein. Die LPZ B beginnt am Rand der Niede-
mit einem massiven Rückgang der Tanne und ra- rung, andererseits aber
scher Ausbreitung der Kiefer (Wald- und vor allem auch durch wechseln-
Bergkiefer, Pinus sylvestris, P. mugo). Erst später de Beweidung und
nimmt der Pollenniederschlag der Fichte aus den Wildäsung bedingt.
forstlichen Beständen zu. Nach historischen Quel- So zeigt eine Zunah-
len (Splechtna 2004) setzte hier schon im 12./13. me von Spitzwegerich
Jahrhundert, also deutlich früher als am 7,5 km (Plantago lanceolata)
entfernten Rotmösel, unter dem Einfluss des Stiftes in der oberen Hälfte
Admont und später der Kartause Gaming die ört- der LPZ B ein vorü-
liche Waldrodung für Holznutzung und Almweide bergehendes Nachlas-
ein („Blumbsuch“, Splechtna 2001), vor allem seit sen der Weideinten- Abb. 9: Das Moor auf der Grubwiesalm mit Lagepunkt der Torfbohrung, vgl.
Mitte des 16. Jahrhunderts. sität, wodurch die Abb. 6. (Foto: K. Splechtna)
sonst stark verbissenen
Erst in der LPZ B tritt die Entwicklung der Almwei- Pflanzen vermehrt zur Blüte und Pollenverbreitung 3.3.3 Grubwies, Moor und Doline
den mit Wacholder, Adlerfarn, Eisenhut (Juniperus, kommen. Das Moor auf der Grubwiesalm, aktuell überwie-
Pteridium, Aconitum) und anderen Pollennachwei- gend ein Carex rostrata-Bestand mit randlichem
sen deutlich hervor. Blühende Eisenhutwiesen prä- Unter den Pioniergehölzen ist in LPZ B eine Aus- Quellaustritt, weist über 20 cm Lehm und lehmig
gen auch heute gelegentlich weite Teile der Senke. dehnung von Lärche und Grünerle nachweisbar. durchsetztem Radizellentorf eine 80 cm mächtige
Die staudenreiche Moorvegetation ist mit Trollblu- Darüber hinaus nimmt in der nun nicht mehr von Wechselfolge verschiedener Riedtorfe mit Beteili-
me, Fingerkraut, Ampfer, Baldrian (Trollius-, Poten- dichtem Wald der LPZ A umgebenen Senke der gung von Braun- und Torfmoosen auf. Der Bohr-
tilla-, Eurumex-Typ, Valeriana) und weiteren Arten prozentuale Anteil von Weit- und Fernflugpollen punkt liegt 6 m von einem Schluckloch entfernt
mit örtlich und zeitlich unterschiedlichen Anteilen zu, sowohl unter den Kulturgehölzen wie Wein, (Abb. 9).
nachgewiesen. Auch die trockenen Randpartien Walnuss, Esskastanie, Platane (Vitis, Juglans, Cas-
sind z.B. mit Spitzwegerich, Sonnenröschen und tanea, Platanus) und mediterranen Bäumen Stein- Die Moorentwicklung (Anhang III) über dem was-
Natternkopf (Plantago, Helianthemum, Echium) eiche, Blumenesche, Steinlinde (Quercus ilex-Typ, serstauenden Lehm ist hier als Versumpfung mit
vertreten. Insgesamt unterliegt die krautige Vege- Fraxinus ornus, Phillyrea), als auch unter den Ge- Ampfer (Rumex acetosa-Pollentyp mit zwei Grö-
tation in und an der Senke einer starken Dynamik, treiden und krautigen Nutzpflanzen (Cerealia p.p., ßenklassen in GM und Eurumex-Typ in GD, ent-
die sich besonders in den Sauer- und Süßgräsern so- Secale, Cannabis) und ihren Begleitern wie Korn- sprechend dem Rumex acetosa- und R. scutatus-Typ
wie Farnen (Cyperaceae, Poaceae, Polypodiaceae), blume, Knäuel, Winde (Centaurea cyanus, Scleran- nach Drescher-Schneider 2007, hier wahrschein-
aber auch in unterschiedlicher Torfzersetzung und thus annuus, Convolvulus arvensis). lich Rumex alpinus und/oder R. arifolius) und16 Silva Fera, Bd. 6/Mai 2017
anhaltender Wassersättigung (Grünalge Botryococ-
cus) fassbar. Der Beginn der Moorbildung in der
Fichten-Tannen-Buchenzeit des Älteren Subatlati-
kums um 895 - 1025 AD ist deutlich jünger als im
Bärwiesboden. Außerdem läuft auf der Grubwies
seit 1325 - 1445 AD dem Tannenrückgang an der
Grenze LPZ a/b ein Rückgang der Buche mit Zu-
nahme der Fichte (und Kiefer) voraus, und schon
um 1200 AD (interpoliert) treten mit dem Wachol-
der erste Anzeichen einer Beweidung auf. Nach his-
torischen Quellen sind örtliche Almweiderodungen
auf der Grubwies allerdings erst seit dem 15. Jahr-
hundert anzunehmen (Splechtna 2004).
In der LPZ b ist die Vegetationsentwicklung der
Neuzeit vor allem in der umgebenden bis heute
bestehenden Fichtendominanz der lichten Alm-
weidelandschaft und einem vermehrten Auftreten
der Lärche deutlich. Die Pollennachweise aus den
tieferen Lagen und weiter entfernten Gebieten (Ju-
glans, Castanea, Cerealia, Secale, Cannabis) reichen
erwartungsgemäß und im Gegensatz zu der älteren
LPZ A der Bärwies in die LPZ a zurück. Dagegen
sind Südalpen- bis mediterrane Elemente (Ostrya-
Typ, Fraxinus ornus, Olea) hier erst in LPZ b be-
legt. Abb. 10: Die Doline auf der Grubwiesalm mit Lagepunkt der Torfbohrung. (Foto: A. Brande)
Das 250 m entfernte, nahezu kreisrunde Kleinst- offensichtlich weitgehend synchron. Besonders in 4. Diskussion
moor der Grubwiesalm-Doline (Abb. 10) ist ein LPZ b fällt ein kurzzeitiges Maximum der Süßgrä-
Polytrichum commune-Bestand auf 20 cm gleich- ser (Poaceae, hier vermutlich die in der Almweide 4.1 Der Beginn der Moorbildung
artigem Torf. Darunter lagern 40 cm Ried- und typische Deschampsia cespitosa, s. Kap. 2 und 4.4) Eine Übersicht zum Beginn der Torfbildung gibt
Braunmoostorfe auf lehmigem Untergrund. auf, verbunden mit Pollennachweisen u.a. bei den Tab. 3. Der Torf liegt im Rotmösel auf Dachstein-
Cichorioideae (vermutlich Leontodon hispidus). Es kalk, in den übrigen drei Untersuchungspunkten
Das Einsetzen der Torfbildung am Bohrpunkt im könnte durch eine kurzzeitige Unterbrechung der auf Lehm. Die Torfbildung setzt im Rotmösel im
Zentrum des Moores (Anhang IV) ist mit 1285 - Almweiderodung in gegenwartsnaher Zeit zurück- zeitlichen Übergang vom Atlantikum zum Sub-
1405 AD etwa 400 Jahre jünger als im Moor der zuführen sein, die sich möglicherweise auch in his- boreal, d.h. der Mittleren zur Späten Wärmezeit,
Grubwies. Die Vegetationsentwicklung der Gehöl- torischen Quellen fassen lässt. Sie fällt in die Zeit ein. Daher ist eine klimatische Ursache der begin-
ze und der örtlichen Moorbestände in den LPZ a nach dem Beginn der Polytrichum-Torfbildung, der nenden Torfbildung (höhere Niederschläge, etwas
und b ist in beiden Mooren jedoch sehr ähnlich und jünger als 1675 AD ist. geringere Jahresmitteltemperaturen) nahe liegend.Silva Fera, Bd. 6/Mai 2017 17
Die durch Karstphänomene wie z.B. Schlucklöcher Der Bärwies-Talboden ist nach Fink (1973: 35) tien, die heute 40 % der Talbodenfläche einnehmen
geprägte Topographie am Rotmösel (Kusel-Fetz- „aus einer glazialen Wanne zu einer Karstmulde (Splechtna & König 2001), wird aber erst mit dem
mann 1981) schließt die Möglichkeit ein, dass es am umgestaltet worden“ und stellt demnach im glazi- mittelalterlichen Tannenrückgang und der Alm-
Untersuchungspunkt im Verlauf der Torfbildung alen Erosionsrelief eine als Kar ausgeformte Karst- weidegewinnung durch das Stift Admont und der
von älteren Moorzentren ausgehend bereits zu ei- hohlform dar. Es ist daher anzunehmen, dass ältere Kartause Gaming stattgefunden haben. Seit dieser
ner ersten Transgression der Torfbildung kam. Ent- Bodensedimente und glazialer Schutt den pollen- Zeit dürfte der gesamte Talboden weitgehend wald-
sprechendes ist auch in anderen Gebieten mit soli- analytisch erfassten eingeschwemmten Basislehm frei geblieben sein (vgl. Abb. 11). Ob die trocke-
topogener Hochmoorentstehung typisch (vgl. die unterlagern. Die heute bestehenden fünf bis sechs neren Partien des Bärwiesbodens heute auch nach
detaillierten Untersuchungen im Harz von Beug Karstquellen am Ost- und Nordrand der Senke so- Jahrhunderte langer Beweidung bzw. Wildäsung
et al. 1999). Ältere Abschnitte der Vegetationsge- wie drei Bäche mit Tümpeln/Lacken/Teichen (Ha- waldfrei bleiben würden, ist trotz der klimatischen
schichte als im Rotmösel wurden 2,5 km nördlich seke 2016) haben vor allem im südlichen Teil der Inversionslage und Schneebedeckung nicht sicher.
des Dürrenstein-Hauptgipfels (OS in Abb. 5) auf Senke zu dauernder Vernässung und somit Torfbil-
der Fichten-Moorinsel im Obersee (14 ha, 1113 m dung geführt. Dabei wurde die wasserstauende, am Die Grubwiesalm ist dem Namen nach ein von
ü. NN) in einer 4,7 m tiefen Bohrung erfasst (Kri- Bohrpunkt mehr als 15 cm mächtige Lehmdecke größeren und kleinen Senken geprägtes Bergwei-
sai 1993). Dort sind über Seeablagerungen 1,8 m zwischen Felsuntergrund und Torf in einer offenbar degebiet. Fink (1973) kartierte eine Vielzahl von
Torfe aufgewachsen. Die vegetationsgeschichtliche voll bewaldeten Umgebung noch ohne menschliche Karstmulden, -wannen und -dolinen auf Lias- über
Überlieferung reicht in den älteren Teil der Mittle- Einwirkungen eingeschwemmt. Die Ursache dürfte Dachsteinkalk. Das Grubwies-Moor im Nordteil
ren Wärmezeit (Atlantikum) und damit in die Zeit ein starker oder eine Folge von episodischen Nie- der Alm weist ebenso wie der Bärwiesboden unter
der beginnenden Buchen-Einwanderung vor ca. derschlagsereignissen in der Naturlandschaft der dem Torf eine pollenführende Lehmabdichtung
7000 Jahren. Im 7 km westlich des Dürrenstein- Nachwärmezeit, dem Älteren Subatlantikum, sein, auf, die gleichfalls der Nachwärmezeit, dem Älte-
Massivs gelegenen Leckermoos (25 ha, 860 m ü. durch das die nachfolgende Moorbildung ausgelöst ren Subatlantikum, angehört. Dasselbe gilt für das
NN) sind in einer 8 m tiefen Bohrung 4,5 m mäch- wurde. Ein farnreicher Talboden (Polypodiaceae Kleinstmoor der Grubwies-Doline. Hier scheint der
tige Hochmoor- und gegenwartsnahe Bruchwald- p.p., wahrscheinlich Dryopteris und Athyrium) bil- Beginn der Torfbildung einige Jahrhunderte jünger
torfe aus einer Seeverlandung hervorgegangen, die dete die Ausgangsvegetation vor der Vernässung. zu sein (Tab. 3), liegt aber ebenfalls noch in der
an den Beginn der Nacheiszeit oder in die Späteis- Zeit vor Einsetzen der örtlichen Landnutzung. Sie
zeit vor 12000 Jahren zurückreicht (Kral 1979). Die Entwaldung des Bärwies-Talbodens hat sich kann daher nicht auf rodungsbedingte Lehmein-
phasenhaft vollzogen. schwemmung und Abdichtung der Hohlformen
Tab. 3: Beginn der Torfbildung in den untersuchten Bohrungen Die Einschwemmung zurückgeführt werden. Ob auf der Grubwies die
des wasserstauenden Lehmeinschwemmung direkt der Entstehung der
Bohrung/Pollendiagramm Torfbasis (cm u. O.) Alter 1 Lehms und die örtli- Karsthohlformen folgte, ist fraglich. Dazu wären
Grubwies Doline (GD) 64 > 1300 AD chen Überflutungen Nachweise von historischen Überlieferungen derar-
mit nachfolgender tiger Einsturzvorgänge erforderlich. Im Grubwies-
Grubwies-Moor (GM) 96 > 900 AD Torfbildung haben Moor spricht der bis in die Gegenwart ungestörte
zum Verlust einiger Torf in unmittelbarer Nähe des Schluckloches da-
Bärwiesboden (BW) 62 > 200 AD Waldstandorte ge- für, dass letzteres erst in jüngster Zeit durch Karst-
Rotmösel (RM) 320 > 4000 BC führt. Eine großflächi- prozesse entstanden ist. In der Grubwies-Doline
ge Freilegung auch der wurde an der Basis des eingeschwemmten Lehms
1
Jahre BC (v. Chr.) und AD (n. Chr.) nach Tab. 1, Spalte 7, gerundet; trockeneren, nördlich das rote Bodenrelikt der ehemaligen Oberfläche
> älter als das genannte Datum gelegenen Senkenpar- über dem Felsuntergrund angetroffen. In beiden18 Silva Fera, Bd. 6/Mai 2017
tigen Torfe (Schmidt 1990, Kral 1994b, Drescher-
Schneider 2007).
In den 78 und 34 cm mächtigen pollenanalytisch
untersuchten Rohhumusprofilen Rothwald I und II
im „Großen Urwald“ liegt der Ablagerungsbeginn
nach Kral & Mayer (1968) ca. bei 1000 AD und
1000 BC. Als Ursache für die Entstehung dieser
dem Felsgestein direkt aufliegenden Ablagerungen
nennen die Autoren einen Bergsturz im Atlanti-
kum, der Mittleren Wärmezeit.
4.2 Die ursprüngliche Waldzusammensetzung
Eine Vorstellung der ursprünglichen Waldzusam-
mensetzung der Klimaxbaumarten Buche, Tanne
und Fichte vor den starken menschlichen Eingrif-
fen geben bei Berücksichtigung der artspezifischen
Pollenproduktion die mittleren Prozentwerte des
Pollenniederschlags aus LPZ A1, a1 und 5 in Tab.
4 entsprechend dem waldgeschichtlichen Abschnitt
IXb bei Kral & Mayer (1968). Auch die Teildiagram-
me Picea/Abies/Fagus = 100 % bieten einen Vergleich.
Abb. 11: Der Große Bärwiesboden und Buchenwaldverjüngung am Sperriedel im Herbstaspekt. Der Zeitraum für die ursprüngliche Waldzusammen-
(Foto: K. Splechtna), aus Splechtna (2001). setzung liegt nach den 14C-Daten in BW im 1. Jahr-
tausend n. Chr., in GM im Bereich der Wende zum 2.
Mooren sind die unteren Torfe lehmig und dem- des Subatlantikums, in Betracht. Dabei bleibt die Jahrtausend und reicht in GD bis in das 14. Jahrhun-
nach noch unter dem abklingenden Einfluss von auf die gesamte Nacheiszeit gesehen sehr junge und dert. RM nimmt den zeitlichen Übergang zwischen
Abschwemmungen oder Nachsackungen in der Na- geringmächtige, aber kontinuierliche Torfbildung BW und GD ein (vgl. Tab. 5: Beginn des Buchen-
turlandschaft gebildet worden. bemerkenswert. Wie in der Bärwies existierte im und Tannenrückgangs). Zusammen genommen ist
Übergang von der Vernässung zur Torfbildung ein damit eine Zeitspanne zwischen 200 und 1350 AD
Den Zeitpunkt der Abdichtung von Karsthohl- örtlicher farnreicher Bestand. Ein solcher ist mit für diese letzte Phase der ursprünglichen, nahezu
formen während, bald oder lange nach ihrer Ent- Dryopteris und Athyrium auch heute in Senken am unbeeinflussten Waldentwicklung im Umkreis der
stehung diskutiert Smettan (2000a) anhand der Rand der Bärwies und im Zentrum mancher klei- Untersuchungspunkte erfasst. Nur in den Rothwald-
Pollenführung. Demzufolge weisen die Befunde nen Dolinen der Grubwiesalm anzutreffen. Diagrammen RW I + II KM reicht diese Phase bis in
der Grubwies und Bärwies darauf hin, dass die pol- das 20. Jahrhundert hinein. Die älteren Phasen der
lenführenden wasserstauenden Lehme erst deutlich Demgegenüber führte in drei pollenanalytisch un- natürlichen Waldentwicklung seit 4000 BC mit der
nach der Ausbildung der Senken abgelagert worden tersuchten Karstsenken des Dachstein-Plateaus erst endgültigen Ausbildung des voll entwickelten Fich-
sind. Hier kommen wie in der Bärwies starke Nie- die hochmittelalterliche rodungs- und almweidebe- ten-Tannen-Buchenwaldes um 2000 BC sind in RM
derschlagsereignisse der Älteren Nachwärmezeit, dingte Lehmabschwemmung zur Bildung der dor- belegt und bei Brande et al. (2002) diskutiert.Silva Fera, Bd. 6/Mai 2017 19
Tab. 4: Pollenniederschlag der Klimaxbaumarten vor Beginn des anthropogenen Buchenrückgangs. KM – Kral & Mayer (1968, Tab. 3 und 4). GM – Grubwies Moor, GD
– Grubwies Doline, BW – Bärwiesboden, RM – Rotmösel, RM KM – Rotmösel bei Kral & Mayer (1968; Korrelation von RM und RM KM bei Brande & al. 2002, fig. 3),
RW I + II KM: Rothwald I und II bei Kral & Mayer (1968).
Höhenlage Probentiefe
Pollendiagramm Pollenzone Probenzahl Picea Abies Fagus
(m ü. NN) (cm u. O.)
GM 1380 a1 9 56-99 29 18 53
GD 1370 a1 5 36-65 38 15 47
BW 1110 A1 3 72-92 26 26 48
RM 1140 5 5 22-40 47 22 31
RM KM 1150 IXb 5 36-52 58 18 24
RW I KM 1200 IXb 6 43-63 36 19 45
RW II KM 1200 IXb 4 15-22 59 6 35
RW I+II KM 1200 IXb 10 15-63 47 13 40
In GM liegt der Buchenanteil des Pollennieder- in Dolinen der nördlichen Kalkalpen liefern Mohr ten ein höhenstufenabhängiger Gradient zwischen
schlags so hoch wie der von Fichte und Tanne (1961) für die Gstettneralm in 1270 m Höhe am 1100 und 1400 m pollenanalytisch nicht nachweis-
zusammen genommen und zeigt damit deutlich Dürrenstein, dem mitteleuropäischen „Kältepol“ 6 bar.
die Buchen-Dominanz in dieser Höhenlage. (Die km nordnordwestlich der Bärwies, sowie Smettan
Fichte ist generell im Pollenniederschlag um das (2000b) für den Grubalmkessel in den Chiemgauer Eine starke örtliche und zeitlich unterschiedliche
Zweifache gegenüber Tanne und Buche überreprä- Alpen in 1230 m Höhe, allerdings in beiden Fällen Prägung des Polleneinzugsgebietes und Standortes
sentiert; vgl. Lang 1994). In dem kleineren Pollen- mit einer Überprägung der inversen Vegetations- mit entsprechend gegensätzlichen Prozentwerten
einzugsgebiet von GD kommt dagegen offenbar stufen durch die Almweidenutzung. Ähnliches wies weisen die Diagramme der Rohhumusprofile RW I
ein örtlicher Anteil der Fichte vom Rand der Do- auch Kral (1969) pollenanalytisch in den Venezia- + II KM aus dem Rothwald-Urwald auf, so dass hier
line vermehrt zum Ausdruck. Dadurch ist auch der ner Alpen nach. von Kral & Mayer (1968) eine Mittelwertbildung
Tannenanteil vergleichsweise am geringsten. In BW vorgenommen wurde. Sie kommt den Verhältnissen
besteht eine – allerdings weniger ausgeprägte – Bu- Das Verhältnis Fichte/Buche ist in RM gegenüber von RM am nächsten, wobei allerdings die Fichte
chen-Dominanz. Buche und Tanne dürften hier an den übrigen drei Untersuchungspunkten zuguns- im Rothwald nicht als Moorrandkomponente wie
den Hängen zwischen 1150 und 1350 m geherrscht ten der Fichte verschoben, in RM KM sogar noch am Rotmösel, sondern als regional gleichrangig be-
haben (Splechtna 2004), während am Bärwies- stärker. Hier sind die vernässten Randbereiche des teiligte Baumart vertreten ist. Zukrigl et al. (1963)
Talboden aufgrund der klimatischen Inversion der Hochmoores zugleich bevorzugte Fichtenstandor- geben im Rothwald-Kerngebiet für die Altbestände
Fichtenanteil höher gewesen sein kann. Aktuelle te, wie sie in geringerem Umfang in GD gegenüber von Fichte/Tanne/Buche bei allen standörtlichen
Beobachtungen zu derartiger Vegetationsumkehr GM belegt ist. Somit ist für die drei Hauptbaumar- und sukzessionsdynamischen Unterschieden ein20 Silva Fera, Bd. 6/Mai 2017
Verhältnis von 18/22/60 an. Demgegenüber er- Tab. 5: Beginn des Buchenrückgangs und des starken Tannenrückgangs
gibt der aus den Diagrammen GD, GM, BW und
RM gemittelte Pollenniederschlag von Fichte/Tan- Buche Tanne
ne/Buche ein Verhältnis von 36/20/44, wobei im Probentiefe Probentiefe
Einzelnen die oben genannten örtlichen Faktoren Pollendiagramm Alter 1 Alter 1
(cm u. O.) (cm u. O.)
sowie die baumartenspezifische Pollenproduktion
und -verbreitung (Lang 1994) zu berücksichtigen GM 50 1400 30 < 1400
sind. GD 35 < 1350 - > 1800 30 < 1350 - > 1800
Zu den Klimaxbaumarten gehört als schwacher BW 70 < 250 - > 600 40 < 600
Pollenproduzent auch der Bergahorn. Die höchsten RM 25 1800 20 < 1800
Werte von Acer mit 1 und 1,5 % des Baumpollens
treten in den beiden Grubwies-Diagrammen auf,
1
Jahre n. Chr. nach Tab. 1, Spalte 7, gerundet; < jünger als, > älter als das genannte Datum
weil hier die Waldbestände in unmittelbarer Nähe
der Bohrpunkte am Rand der kleinen Senken lie- Stillstand der Torfbildung, Zäsur mehrerer Pollen- 2.3.2) nach den historischen Quellen (Splechtna
gen. kurven, s. Kap. 2.3.2). 2001) vermutlich in der Zeit bald nach 1700.
In RM ist der Übergang in die tannenarme Zeit
4.3 Der Buchen- und Tannenrückgang Bei den drei Klimaxbaumarten ist der nachfolgende ziemlich abrupt. Zur Ursache des Tannenrückgangs
Schon vor dem markanten Tannenrückgang sind Tannenrückgang besonders einschneidend. Er mar- durch Holzeinschlag und -verwendung vgl. allge-
Eingriffe in die Waldbestände nachweisbar. In al- kiert die Grenze der LPZ 5/6 in RM, A/B in BW mein z.B. Kral (1994a: 31): Waldweide und Kahl-
len vier Diagrammen geht dem starken Buchen- und a/b in DM und GD. Die Abies-Minimalwerte schlagwirtschaft, Flößbarkeit der Tanne im Gegen-
und Tannenrückgang ein erster, vermutlich bereits liegen nunmehr bei 1 - 2 % des Baumpollens. Mit satz zur Buche.
durch menschliche Nutzung verursachter schwä- dem Tannenrückgang ist meist eine Zunahme der
cherer Rückgang der Buche in A2, a2 bzw. 5 p.p. Lichtbaumart Lärche verbunden, dazu in RM und Ein näherer Vergleich mit den Rohhumus-Pollen-
voraus (Tab. 5). So nimmt die Buche besonders in BW auch von krautigen Pflanzen der örtlichen Um- diagrammen Rothwald I und II aus dem „Großen
der LPZ a2 von GM und GD mehr oder minder gebung (Plantago lanceolata, Urtica u.a.) sowie Ge- Urwald“ (Kral & Mayer 1968) ist wegen der speziel-
stetig ab, wobei Fichte und Kiefer zunehmen. Zu- treide (Cerealia p.p., Secale) aus den tieferen Lagen. len Ablagerungs-, Pollenniederschlags- und Stand-
gleich wird Wacholder als Weidezeiger etwas häu- ortbedingungen in dem dortigen Bergsturzgebiet
figer. Die 14C-Daten weisen hier auf das Ende des Nach den 14C-Daten in RM und GD – übertrag- nicht möglich. Der unterschiedlich stark ausgepräg-
14. Jahrhunderts (Tab. 5). Im Übrigen ist aber der bar auf GM – ist die tannenarme Zeit überwiegend te und zeitversetzte Tannenrückgang in den beiden
Einfluss der Almweide auf der Grubwies noch ge- jünger als 1655/75 AD und umfasst die Zeit nach Rohhumusprofilen (in RW II nach 800 v. Chr., in
ring. Aus den tieferen Lagen stammt der Pollen von 1800. Der Beginn ist in RM wohl frühestens um RW I um 1200 n. Chr.) kann jedoch nicht, wie z. B.
Getreide und Hanf (Cerealia p.p., Secale, Cannabis) 1750 anzusetzen (Brande et al. 2002, vgl. Splecht- Küster (1994: 26) für das bayerische Alpenvorland
und in GM auch von Walnuss (Juglans) und Ess- na 2001, Pekny 2012). Er fällt in die Zeit intensiver schreibt, mit einem römerzeitlichen Gebrauch von
kastanie (Castanea). Schwächer sind die genannten Rodungen für den gesteigerten industriellen Ener- Tannenholz in Verbindung gebracht werden. Er ist
Einflüsse in RM. Sie fehlen weitestgehend in BW, giebedarf und ist damit etwa 550 Jahre jünger als nach Kral & Mayer (1968) in RW II klimatisch und
wo die zeitgleichen Sedimente den 14C-Daten zu- von Kral & Mayer (1968), Kral (1974) und Mayer in RW I durch die beginnende mittelalterliche, nach
folge allerdings sehr komprimiert sind oder eine et al. (1979) angenommen. Auch in BW liegt der der o.g. Datierung wahrscheinlich sogar erst neu-
Schichtlücke vorliegt (Hiatus, vorübergehender rapide Tannenrückgang (evtl. Hiatus, vgl. Kap. zeitliche Holznutzung bedingt.Sie können auch lesen

















































