Ff - Grundlagen und erste
←
→
Transkription von Seiteninhalten
Wenn Ihr Browser die Seite nicht korrekt rendert, bitte, lesen Sie den Inhalt der Seite unten
Osnabrücker Natunryissenschaft liche M itteilungen
Band 2O/21,S. 275-337, 1995
^ff
Biozönotische Untersuchungen in Hudelandschaften
Nordwestdeutschlands - Grundlagen und erste
Ergebnisse -+
Thorsten Aßmann & Anselm Kratochwill
unter Mitarbeit von Anja Niemann und Elke Vossel
Kurzfassung: Hudelandschaften stellen im nordwestdeutschen Tiefland Lebensräume besonders
hoher biologischer Diversität dar. Während einer über lange Zeiträume andauernden extensiven Bewirt-
schaftung durch den Menschen entstand ein vielfältiges Mosaik unterschiedlicher Vegetationseinhei-
ten. Die von geobotanischer Seite gut untersuchten Pflanzengesellschaften und Vegetationskomplexe
bieten sich als Objekte biozönologischer Forschung an. Von besonderer wissenschaftlicher Bedeutung
ist das Studium von Reliktarten-Gemeinschaften alter Wälder. Unter den Carabiden, Chilopoden und
Limaciden können Reliktarten benannt werden. Freilandexperimente zum Ausbreitungsverhalten und
populationsgenetische Untersuchungen an Carabiden wurden durchgeführt, um Unterschiede zwi-
schen einer Reliktart und einer weit verbreiteten Art untersuchen zu können. Strukturreiche, ehemalige
Hudewälder besitzen im Vergleich zu den strukturarmen Wirtschaftshochwäldern unterschiedliche
Artengemeinschaften an Chilopoden, Diplopoden und Carabiden. Die Hudelandschaften weisen eine
Reihe von Teil-Lebensräumen auf, die heute landesweit hochgradig gefährdet sind. Hierzu gehören z.B.
die Tot- und Altholzbestände, die durch besonders typische Zersetzer-Gemeinschaften unter den
Arthropoden charakterisiert sind; auch diese sind Gegenstand unserer biozönologischen Untersuchun-
gen. Ein Schwerpunkt unserer Forschung liegt in der Gharakterisierung einzelner Zoo-Taxozönosen
(Coleoptera, Lepidoptera, Diptera, Orthoptera) auf der Basis des pflanzensoziologischen Rasters. lm
Bereich der Ornithofauna haben wir uns bisher intensiver mit den Meisen-Gemeinschaften ausgewähl-
ter Hudegebiete befaßt und die verschiedenen Bindungsgrade an spezifische Ressourcen der Stand-
orte analysiert. lm Rahmen dieserArbeit wird gezeigt, daß die Erforschung von Biozönosen unter Land-
schaftsbezug und unter Berücksichtigung von Pflanzengesellschaften und Vegetationskomplexen
wichtige Ergebnisse liefert. lm Falle der Hudelandschaften kommt auch kulturgeschichilichen Zusam-
menhängen eine große Bedeutung für das Verständnis des Aufbaues solcher Ökosysteme zu.
Abstract tn North-West Germany grazed woodland landscapes represent ecosystem complexes of
especially high biological diversity. Owing to a long-lasting extensive management, a diverse mosaic
of different vegetation structures has developed. Plant communities and vegetation complexes which
have been well investigated by geobotanists are suitable subjects for biocoenological research. The
study of relic communities of ancient woodlands is of particular scientific importance. Among Carabi-
dae, Chilopoda, and Limacidae, relic species can be named. Outdoor experiments concerning sprea-
ding characteristics as well as population genetical investigations were performed to study differences
between a relic species and a wide-spread species. Former grazed woodlands diverse in structures
possess different coenoses in Chilopoda, Diplopoda, and Carabidae, contrary to the economically used
high forests which are poor in structures (planted even-aged stands). The grazed woodland landsca-
pes contain a number of species and biotopes which are today extremely endangered throughout the
country. One example for this is the stock of dead and old wood which is characterized by particularly
typical decomposing communities among the Arthropoda; this is also investigated by us. One main
focus of our research lies in the characterisation of specific zoo-taxocoenoses (Coleoptera, Lepidopt-
* Veröffentlichung der Arbeitsgemeinschaft für Biologisch-Ökologische Landeserforschung, ABöL,
Nr. 109, Münster. Gefördert mit Forschungsmitteln des Landes Niedersachsen.
1 Meinem verehrten Lehrer Prof. Dr. G. Os+re, Freiburg gewidmet.
275
Thorsten Aßmann & Anselm Kratochwil Osnabrücker Naturwiss. Mitt. 20121 1995
era, Diptera, Orthoptera) on the basis of the phytosociological system. ln the field of ornithofauna, we
have dealt more intensively with the titmouse communities of selected grazed woodland areas and
have analysed the different levels of binding to specific resources of the sites. ln the frame of this work
we show that the analysis of biocoenoses - in view of the landscape, plant communities and vegeta-
tion complexes - leads to important results. ln the case of woodland pasture landscapes, the histo-
rico-cultural influences are highly important in order to comprehend the structure of such ecosystems.
Key words: grazed woodland ecosystems, relation fauna,/vegetational complexes, research concept,
ancient woodlands, relict fauna, Coleoptera, enclosure, power of dispersal, Chilopoda, Diplopoda, tit-
mouses
Autoren:
Dr. Th. Aßmann, Prof. Dr. A. Kratochwil, Fachgebiet Ökologie, Fachbereich Biologie/Chemie, Universi-
tät Osnabrück, Barbarastraße 1 1, D-49069 Osnabrück.
lnhalt
1 Einleitung. ...277
2 Einige grundsätzliche Anmerkungen zur biozönologischen Arbeitsweise . . . 279
3 Vegetationskomplexe in den untersuchten Hudelandschaften . . . . 281
3.1 Die potentielle natürliche Vegetation und die hudebedingten
Ersatzgesellschaften ...281
3.2 DerEichen:Birkenwald-Vegetationskomplex. ...282
3.3 DerBuchen-Eichenwald-Vegetationskomplex ....285
3.4 DerEichen-Hainbuchen-Vegetationskomplex.. ...286
3.5 Auen-Vegetationskomplexe. ......287
4 Fragestellung und Zielsetzung unserer Forschung in den
HudelandschaftenNordwestdeutschlands ....287
5 Biozönologische Untersuchungen in Hudelandschaften . . . . 289
5.1 Reliktarten und Reliktarten-Gemeinschaften alter Wälder . . . 289
5.1.1 Beispiele für Reliktarten alter Wälder . . . . 289
5.1.2 Reliktarten-Gemeinschaften von Hudewald-Standorten im Vergleich
zumWirtschaftshochwald . . . . .303
5.2 Coleopteren-Zönosen charakteristischer Landschafts-Strukturen . . 311
5.2.1 Xylobionten-Synusien . . . 311
5.2.2 Coleopteren-Zönosen unterschiedlicher Offenland-Standorte . . 312
5.3 Hudelandschaften, ihre Bedeutung für die Ornithofauna . . . . 315
5.3.1 Einführung ....315
5.3.2 Die Bindung der Weiden- und Schwanzmeise (Parus montanus,
Aegithalos caudatus) an den Wacholder (Juniperus communis\ . . . . . 318
5.3.3 Die Bedeutung hudespezifischer Baumarten und -strukturen für
Ganzhöhlenbrüter. ...323
5.3.4 Die unterschiedlichen Nahrungshabitate in verschiedenen Landschaften . . - 325
6 AusblickundDank.. ...329
Literatur. .---.330
276
Hudelandschaften, Biozönosen
1 Einleitung heute unter Naturschutz stehende Gebiete
mit Hudelandschafts-Charakter (Pohl 1 975,
lm nordwestdeutschen Tiefland zählen 1983, Runge 1992) ein Abbild und ein Zeug-
Hudelandschaften zu den Lebensräumen nis der landschaftsgestaltenden Wirkung
höchster biologischer Diversität, dies gilt dieser für Nordwestdeutschland kulturhisto-
sowohl auf Artniveau (a-Diversität) als auch risch bedeutsamen Form der Landnutzung.
auf Zönosen-Ebene (p-Diversität). lhre Ent- Das Landschaftsbild, wie es sich uns heu-
stehung verdanken sie der langen, teilweise te zeigt, spiegelt die unterschiedliche Art
über mehrere Jahrtausende währenden und Weise, Dauer und lntensität extensiver
extensiven Bewirtschaftung durch den Men- Nutzung wider. Besonders typisch ist der
schen, im wesentlichen seiner Weide- sich dem Besucher vermittelnde Eindruck
(= Hude-) Wirtschaft. Nach pollenanaly- einer ,,Parklandschaft" (Abb. 1). Sie setzt
tischen Befunden dürfte sich die Dauer der sich zusammen aus einem Mosaik vegeta-
Waldhude allein über einen Zeitraum von ca. tionsfreier oder -armer Dünen, grasreicher
5000 Jahren erstreckt haben (Pott & Hüppe offener Triften, aus Staudensäumen, reich-
1991). Somit geben in ihrem besonderen gegliederten Waldmänteln, eingesprengt
Strukturreichtum und ihrem charakteristi- sind dazwischen Einzelbäume und Baum-
schen Pflanzen- und Tierartengefüge viele, gruppen unterschiedlicher Ausdehnung.
Abb. 1: Die ,,Haselünner Kuhweide"erweckt in besonders bezeichnender Art und Weise den Charakter einer
,,Parklandschaft". Auf Querco-Ulmetum-Standorten finden sich heute großflächig Weide-Gesellschaften, ein-
gesprengt sind Wacholder-Gebüsche, die in einigen Bereichen des Gebietes besonders großflächige
Bestände bilden. Kulissenartig stocken lückige Eichenwäldchen, vorgelagert sind Strauchmäntel. Tiefer lie-
gende lehmige und feuchte Flutmulden verzahnen sich hier mosaikartig mit höher gelegenen sandigen Ter-
rassen. Foto Kratochwil.
277
Thorsten Aßmann & Anselm Kratochwil Osnabrücker Natunrviss. Mitt. 20121 1 995
o Hofgehölz Möhr
i a (Wachendorfer Wacholderhain
50 km
Abb. 2: Lage unserer Untersuchungsgebiete in Nordwestdeutschland
Bezeichnend sind ferner lückige Waldgesell- landschaften Nordwestdeutschlands" leg-
schaften mit einem hohen Anteil besonders ten Pott & Hüppe (1991)vor. Auf der Basis
charakteristischer Baum-lndividuen, die einer solchen grundlegenden Analyse, wie
häufig Zeichen und Merkmale der einstigen sie von geobotanischer Seite vorgenommen
Hude tragen: Kappungsformen, Schneitel- wurde, kann auch eine edolgversprechende
male, Verbißmarken und Verbißteller (s. Bur- biozönologische Forschung einsetzen. Wir
richter 1984, Burrichter & Pott 1983, vgl. beschränken uns hierbei zunächst nur auf
auch Kap. 5.1.2). Tiefland-Standorte und klammern die von
Die große biologische Vielfalt der Hude- Pott & Hüppe (1991) randlich berücksichtig-
landschaften Nordwestdeutschlands ist aus ten Mittelgebirgslagen (westfälische Hügel-
geobotanischer und landschaftsökologi- und Berglandschaften, Ravensberger
scher Sicht durch Untersuchungen von Bur- Hügelland, das Weser- und Wiehengebirge
richter und seinen Schülern inhaltsreich sowie die Sand- und Kalkstein-Züge des
dokumentiert worden (2.8. Burrichter 1970, Teutoburger Waldes und des Eggegebirges)
1977, 1988, Burrichter et al. 1980, Pott aus. Die von uns untersuchten und im Text
1983, 1990, Pott & Burrichter 1983). Eine erwähnten Gebiete sind in der Abb. 2 darge-
umfassende Monographie über die ,,Hude- stellt.
278
Hudelandschaften, Biozönosen
lm folgenden seien zunächst einige 2 Einige grundsätzliche Anmer-
grundsätzliche Gesichtspunkte zur biozöno- kungen zur biozönologischen
logischen Arbeitsweise vorangestellt. Da Arbeitsweise
das Grundmuster der Vegetationszusam- (s. ausführlich bei Kratochwil 1987, 1991)
mensetzung einschließlich ihres Entwick-
lungspotentials durch die jeweils herrschen- Biozönosen setzen sich zusammen aus
den Standoftsbedingungen vorgegeben ist, Phytozönosen, den Pflanzengesellschaften,
bieten sich als Gliederungsschema oder aus Zoozönosen, den Tiergemeinschaften,
Matrix für biozönologische Zusammen- und bestimmte, durch Pflanzen- oder Tier-
hänge am besten die pflanzensoziologisch arten aufgebaute Synusien. Synusien sind
charakterisierten Vegetationskomplexe an, ökologisch und symmorphologisch einan-
die als eindeutig unterscheidbare Land- der nahestehende und unter annähernd
schaftstypen präzise voneinander differen- gleichen kleinstandörtlichen Bedingungen
ziert werden können (Kratochwil 1987, 1991 , lebende Artengruppen, z.B. Moos- oder
Schwabe 1988, 1990). Sie sind der Schlüs- Flechtensynusien innerhalb von Pflanzenge-
sel für viele biozönologische Gesetzmäßig- sellschaften. Sehr reichhaltig ist die Anzahl
keiten, wie wir sie auch hier am Beispiel ein- der verschiedenen Tiersynusien, z.B. die
zelner Zoozönosen in Hudelandschaften Zersetzergemeinschaft eines Holzstubbens
zeigen möchten. Aus diesem Grund werden oder einer Tierleiche.
zunächst in einem eigenen Kapitel diejeni- Die Basis für die Erfassung einer Biozö-
gen Landschaftstypen vorgestellt, in denen nose sollte immer die pflanzensoziologische
unsere biozönologischen Untersuchungen Charakterisierung des zu untersuchenden
durchgeführt wurden. Lebensraumes sein. Hierdurch wird präzise
ln Anschluß daran seien Fragestellungen die einheitliche Standortsbeschaffenheit ei-
und Zielsetzungen unserer Forschung in nes Geländeausschnittes ausgedrückt.
Hudelandschaften vorgestellt. Es folgen Besonders mit pflanzensoziologischen
dann ausgewählte Beispiele zu den The- Methoden erfaßte Vegetationskomplexe
men: sind für die Charakterisierung von Tierhabi-
taten, sowohl auf der Ebene der Einzelart
Reliktarten und Reliktarten-Gemeinschaf- (Schwabe & Mann 1990, Schwabe et al.
ten unter ausgewählten Tiergruppen (Car- 1992), als auch auf der Ebene von Zoo-
abidae, Chilopoda, Limacidae, Amphibia); Taxozönosen und Gilden besonders genaue
Kap. 5.1.1. Bezugseinheiten (Kratochwil 1984, 1989a,
Die Struktur von Hudewald-Standorten 1989b, Kratochwil & Klatt 1989, Schwabe &
und ihr Einfluß auf die Zusammensetzung Kratochwil 1984). Hierbei werden Pflanzen-
von Zönosen (Carabidae, Chilopoda, gesellschaften aufgrund ihres regelmäßigen
Diplopoda); Kap. 5.1.2. gemeinsamen Vorkommens zu landschaft s-
Coleopteren-Zönosen charakteristischer bezogenen Einheiten, eben den Vegeta-
Hudewald-Strukturen (Xylobionten-Ge- tionskomplexen zusammengestellt. Vegeta-
meinschaften, Käfergemeinschaften an tionskomplexe stellen - wie die Pflanzenge-
Offenland-Standorten); Kap. 5.2. sellschaften auch - typisierbare Einheiten in
Hudelandschaften, ihre Bedeutung für die ökologischer, struktureller, dynamischer,
Ornithofauna (das Beispiel der Meisen); arealkundlicher und evolutionsbiologischer
Kap.5.3. Sicht dar.
279
Thorsten Aßmann & Anselm Kratochwil Osnabrücker Naturwiss. Mitt. 20121 1995
Der 1. Schritt einer biozönologischen voneinander zu unterscheiden, zum einen
Bestandsanalyse muß immer die Charakte- die zönobionten (spezifischen) Arten, die
risierung der räumlichen Strukturvielfalt sein ausschließlich oder nahezu ausschließlich in
(Kratochwil 1987), wobei in der Regel ver- einer bestimmten Zönose vorkommen, zum
schiedene Strukturtypen unterschieden anderen die zönophilen (präferenten) Arten,
werden können, die jeweils auch eigene tie- die sich in einer bestimmten Zönose optimal
rische, zum Teil aber auch pflanzliche Arten- entwickeln, aber auch in anderen vorkom-
gruppen beherbergen. ln Anlehnung an men. Diese euzönen Arten stellen die Cha-
Tischler (1947) sind 3 Strukturtypen, denen rakterarten einer Zönose dar. Keine Charak-
eigene Zönosen zugeordnet werden kön- terarten sind die tychozönen, azönen und
nen, voneinander abgrenzbar (s. Tab. 1). Für xenozönen Arlen.Zu den tychozönen Aden
diese Bereiche können charakteristische gehören solche, die aufgrund ihrer großen
Tiergruppen (Zoo-Taxozönosen) oder ökologischen Amplitude in vielen verschie-
bestimmte ökologische Gilden ausgewählt denen Zönosen vorkommen können (eury-
werden. öke Arten). Für die Charakterisierung von
Der 2. Schritt besteht in der Festlegung Zoozönosen können tychozöne Arten eine
von Charakterarten. Hierzu gehören nur die große Bedeutung als Differentialarten ha-
euzönen Arten (stenöke Standorts-Speziali- ben. Azöne Arten (Ubiquisten) zeigen keine
sten). lnnerhalb dieser Gruppe sind 2 Typen erkennbare Bindung an eine Zönose, zu den
Tab. 1 : Die verschiedenen Raumstruktur-Typen innerhalb eines Biotops (in Anlehnung an Tischler 1947).
TOPISCHE EINHEIT BIOTISCHE EINHEIT BEISPIELE
STRATOTOP STRATOZÖNOSE
horizontale Strukturen Kronenschicht
Strauchschicht
Krautschicht
Streuschicht
Krautschicht
CHORIOTOP cHoRtozÖNosE
eigenständi ge vertikale Baum
Strukturen des gesamten Baumstumpf
Raumes oder Teile des Strauch
Stratotopes Tierleiche
Exkremente
Ameisenhaufen
Vogelnest
MEROTOP MEROZÖNOSE
Struktur-Element inner- Blattbewohner
halb eines Strato- oder Holzbewohner
Choriotopes Rindenbewohner
Blütenbewohner
Blütenbesucher
280
Hudelandschaften, Biozönosen
Ass.lV
x
o
E-
E
voo
C
o
(6
o
o,
o
Ass. I
I
synta(o-
nomische
Ass.5 Einheiten
Klasse 2
-
Abb. 3: Unterschiedliche Bindungsgrade von Tierarten und Tierarten-Gruppen an verschiedene pflanzenso-
ziologische Einheiten: a) Assoziations-spezifische Tierarten und Tierarten-Gruppen, b) Vegetationskomplex-
spezifische Arten und Artengruppen, c) Verbands-, Ordnungs- oder Klassen-spezifische Arten- und Arten-
gruppen.
xenozönen Arten gehören zufällig in der xen vorhanden sind (Beispiele s. Kratochwil
betreffenden Zönose auftretende Arten. Sol- 1991).
che lrrgäste können unter Umständen Cha- Der 3. Schritt besteht in der Abgrenzung
rakterarten anderer Zönosen sein. gemeinsam vergesellschafteter Artengrup-
Tiere müssen keinesfalls wie im Falle a pen, in der Regelvon Stratotop-, Choriotop-
des Diagramms (Abb.3), in dem auf der oder Merotop-spezifischen Zoo-Taxozöno-
Abszisse verschiedene pflanzensoziolo- sen.
gisch-syntaxonomische Einheiten, auf der
Ordinate Vegetationskomplexe schemati-
siert aufgetragen sind, nur an eine Assozia- 3 Vegetationskomplexe in den
tion gebunden sein. Dies ist der seltenere untersuchten Hudelandschaften
Fall. Für die meisten Tierarten ist ein Mosaik
bestimmter Pflanzengesellschaften als 3.1 Die potentielle natürliche Vegetation
Lebensraum notwendig (Fall b). Das Aufsu- und die hudebedingten Ersatzgesell-
chen verschiedener Pflanzengesellschaften schaften
bedeutet dabei nicht, daß solche Tierarten
euryök sein müssen. Das Gegenteil ist häu- Maßgeblicher Faktor für die naturräumliche
fig der Fall: Gerade bestimmte Requisiten Gliederung der Vegetation eines Gebietes
und Ressourcen sind es, die in Kombination sind die verschiedenen Einflüsse der Aus-
nur in ganz bestimmten Vegetationskomple- gangsgesteine und des Bodens. Neben die-
281
Thorsten Aßmann & Anselm Kratochwil Osnabrücker Naturwiss. Mitt. 20121 1995
sen primären Faktoren und Faktorengrup- U lmen-Auenwald (Querco-Ulmetum) reprä-
pen treten mit dem Menschen und seinen sentiert.2
Siedlungs- und Wirtschaftsweisen jedoch Mit Bezug zur lntensität des anthropo-
sekundäre Faktoren und Faktorengruppen zoogenen Einflusses sind zwei Epochen
hinzu, die mit besonderer Nachhaltigkeit das voneinander zu unterscheiden: ein erster
Vegetationsbild einer Landschaft prägen Abschnitt, der von der vorgeschichtlichen
können. Ausgehend von einer mehr oder Zeit über das Mittelalter bis zu Beginn der
weniger geschlossenen Laubwald-Land- Neuzeit reicht (Epoche der Extensiv-Wirt-
schaft hat der Mensch, so auch im nord- schaft), und ein 2. Abschnitt seit dem 18./19.
westdeutschen Tiefland, seit dem Neolithi- Jahrhundert (Epoche der lntensiv-Wirt-
kum (Atlantikum) den Wald zunehmend schaft) (Pott & Hüppe 1991).
zurückgedrängt und für halbnatürliche bis Umfang und lntensität des anthropo-zoo-
hin zu stark anthropogen bedingten Vegeta- genen Einflusses variieren. Zu alten Wald-
tionseinheiten Lebensraum geschaffen. Die- standorten zählen die zu Beginn des 12. und
ser bis heute andauernde Einfluß des Men- 13. Jahrhunderts und in darauffolgenden
schen (und seiner Weidetiere) verläuft paral- Jahrhunderten in herrschaftlichen Bann ge-
lel mit der Großklima-bedingten Sukzession legten, ehemaligen Allmend-Wälder, die z.T.
der Vegetationsabfolge (M itteleuropäische seither ununterbrochen bei extensiver Hude-
Grundfolge der Waldentwicklung), die eben- und Schneitelnutzung Wald tragen. Als Bei-
falls einen besonders starken Einfluß auf die spiele seien der Bentheimer Wald, der Neu-
Dynamik und Genese der Vegetationszu- enburger ,,Urwald" oder der,,Hasbruch" bei
sammensetzung genommen hat (postglazi- Delmenhorst angeführt. Hierbei handelt es
ale Wärmezeit, mittlere und späte Wärme- sich um ganz besonders wichtige biozöno-
zeit, Atlantikum, Subboreal, Subatlantikum). logische Untersuchungsobjekte, wie wir noch
Beide Prozesse zueinander in Beziehung zu sehen werden. lm folgenden werden die für
setzen, ist ein Schlüssel zum Verständnis unsere Untersuchungen zugrundegelegten
der heutigen potentiellen natürlichen Vege- Vegetationskomplexe detail ierter vorgestellt.
I
tation (Burrichter et a|.1988). Somit existiert
die ursprüngliche natürliche Vegetation seit
dem jüngeren Atlantikum im Gebiet nicht 3.2 Der Eichen-Birkenwald-Vegetations-
mehr (Pott & Hüppe 1991). komplex (Abb.4, Tab. 2)
Die potentielle natürliche Vegetation des
näheren Untersuchungsgebietes (s. Abb. 2) lnnerhalb der bodensauren Eichen-
bilden einerseits bodensaure Eichen- Mischwälder (Unterverband Quercenion
Mischwälder (Quercion robori-petraeae) auf roboris), die artenarme, acidophytische Wäl-
den Sandflächen, im Bereich der stau- und der umfassen, und die im wesentlichen eine
grundwasserbeeinflußten Standorte mit an die Geest gebundene, nordwesteuropäi-
Lehmböden Eichen-Hainbuchenwälder (Stel- sche Verbreitung besitzen, stockt auf reinen
lario-Carpinetum). Hinzu treten auf nassen Quarzsanden, z.T. durch Podsolierungsvor-
und zeitweise überstauten Böden Bruchwäl- gänge gekennzeichnet, der Eichen-Birken-
der (im wesentlichen Carici elongatae-Alne- wald, das Betulo-Quercetum roboris. Unter
tum und Betuletum pubescentis). Entlang
größerer Fließgewässer wird die potentielle 2 Die pflanzensoziologischen Bezeichnungen fol-
natürliche Vegetation durch den Eschen- gen im wesentlichen Pott (1992).
282
Hudelandschaften, Biozönosen
Abb. 4: Eichen-Birkenwald-(Betulo-Quercetum-) Vegetationskomplex im ,,Wachendorfer Wacholderhain":
Dicrano-Juniperetum und Genisto-Callunetum mit eingesprengten Birken (Betula pendula) als Relikte des
Betulo-Quercetum. Foto Kratochwil.
Waldweide lichtet sich der Bestand des Sandginster-Heiden (Genisto pilosae-Callu-
Betulo-Quercetum auf und ein Wacholder- netum). Sie bilden häufig mosaik- und kulis-
busch, das ,,anspruchslose" und Acidophy- senartige Komplexe mit dem Dicrano-Juni-
ten-reiche Dicrano-Juniperetum communis peretum. Auf wechselfeuchten Quazsand-
dringt als Weide-Gesellschaft in die böden können sich sekundär im Bereich des
Bestände ein (s. auch Tüxen 1974). Unter Betulo-Quercetum molinietosum auch Eri-
den hier behandelten Waldgesellschaften ist ca-Heiden entwickeln. Dort, wo offene Flug-
die ,,Widerstandskraft" gerade des Betulo- sande anstehen, kommt als Pioniergesell-
Quercetum gegen Beweidung am gering- schaft die Frühl n gsspark-Sil bergrasf u r, das
i I
sten. Schon Ellenberg (1954)wies daraufhin, Spergulo vernalis-Corynephoretum cane-
daß das Betulo-Ouercetum besonders scentis, vor und bildet je nach Standortsdy-
anfällig gegenüber Beweidung ist und eine namik recht unterschiedliche Ausbildungen.
,,Öffnung" des Waldes durch den anthropo- Auf zur Ruhe gekommenen, festgelegten,
zoogenen Einfluß ohne Mühe erfolgte. Somit schwach humosen Sanden finden sich oft
erklärt sich auch die Koinzidenz zwischen nur kleinflächig Kleinschmielenrasen, so z.B.
Betulo-Quercetum-Gebiet und den Altsie- die Gesellschaft des Frühen Schmielenha-
del-Landschaften (Ellenberg, ebd.). fers, das Airetum praecocis, auf sauren,
lm Betulo-Quercetum-Gebiet finden wir stark verdichteten Gleypodsolen die Borst-
als hudebedingte Ersatzgesellschaften grasrasen-Gesellschaft der Sparrigen Binse,
283
Thorsten Aßmann & Anselm Kratochwil Osnabrücker Naturwiss. Mitt. 20121 1995
Tab.2: Übersichtstabelle: Betulo-Quercetum- und Lonicero-Fagetum-Vegetationskomplexe als Elemente der
Quercenion roboris-Landschaft unter Berücksichtigung der hudebedingten Ersatzgesellschaften. Die Reihen-
folge der Anordnung folgt der zunehmenden Öffnung des Waldes und der steigenden lntensität des mensch-
lichen Einflusses.
Quercenion roboris-Vegetationskomplex (Landschaft)
Unterverband: Bodensaure Eichen-Mischwälder
,,Primärwald" Betulo-Quercetum roboris Lonicero-Fagetum
(Eichen-Birkenwald) (Buchen-Eichenwald)
ehemaliger beweidetes Betulo-Quercetum beweidetes Lonicero-Fagetum:
Weidewald, z.T. im Komplex mit dem Fazies mit Pteridium aquilinum, llex
heute z.T. Dicrano-Juniperetum aquifolium-Vorkommen, Komplex mit
Bannwälder (Wacholder-Gebüsch) Sarothamnus scoparius-Gebüschen
Hqdewaldreste und hudebedingte Ersatzgesellschaften im Gebiet des
Quercenion roboris-Vegetationskomplexes:
Wald- Betulo-Quercetum roboris Lonicero-Fagetum
fragmente Hudewaldreste Hudewaldreste
Heiden Genisto pilosae-Gallunetum reichere Calluna vulgaris
(Sandginster-Heide) Bestände und Juniperus
im Komplex mit dem communis-Gebüsche
Dicrano-Juniperetum
z.T. Erica tetralix- Heiden
Vormäntel, Sarothamnus scoparius-Vormantel,
Säume Teucrium scorodonia-Säume
Sand- Spergulo-Gorynephoretum Diantho-Armerietum
Trockenrasen (Frühlin gsspark-Silbergrasf lur) (Grasnelken-Flur)
Pionierfluren Thero-Airion-Gesellschaft en : Thero-Airion-Gesel lschaft en:
Airetum praecocis Filagini-Vulpietum myurus
(Ges. d. Frühen Schmielenhafers) (Federschwingel-Rasen)
Borstgras- Nardo-Juncetum squarrosi
rasen (Ges. d. Sparrigen Binse)
Grasland Lolio-Cynosuretum luzuletosum Lolio-Gynosuretum typicum
284Hudelandschaften, Biozönosen
Abb. 5: Das Lonicero-Fagetum als Element des Buchen-Eichenwald-Vegetationskomplexes; etwa 200 Jahre
alter Buchenbestand im Gebiet des ,,Wachendorfer Wacholderhains". Foto Niemann.
das Nardo-Juncetum squarrosi. Eine die aufgrund bestimmter lnhaltsstoffe ge-
magere Ausbildung der Weidelgras-Weiß- mieden werden, z.B. der Adlerfarn (Pteri-
klee-Weide (Lolio-Cynosuretum luzuleto- dium aquilinum), andererseits die bewehrten
sum)schließt bei intensiverer Beweidung in Arten, z.B. die Hülse (llex aquifolium) und
der Vegetationsentwicklung an. der Besenginster (Sarotha mnus scoparius).
Die Ersatzgesellschaften des Lonicero-
Fagetum vermitteln von der Physiognomie
3.3 Der Buchen-Eichenwald-Vegeta- und vom Artenspektrum her ein anderes Bild
tionskomplex (Abb. 5, Tab. 2) als die des Betulo-Quercetum. Analog treten
hier Heiden mit Calluna vulgaris, ebenso
Ebenfalls zu den bodensauren Eichen- Wacholderbüsche (Juniperus communis)
Mischwäldern (Unterverband Quercenion auf. Die wüchsigeren Standorte bieten je-
roboris) zählt der im Vergleich zum Betulo- doch im Vergleich zum Betulo-Quercetum
Quercetum auf nährstoffreicheren, anleh- eine,,Lizenz" für reichstrukturiertere Stand-
migen Sandböden vorkommende Buchen- orte. Als Relikt des lichten Hudewaldes bildet
Eichenwald, das Lonicero-Fagetum (früher der Besenginster (Saroth amnus scoparius)
als Fago-Quercetum bezeichnet). lm Loni- Vormantel-Gesellschaften aus, davor wach-
cero-Fagetum förden Weidetiere die Verbiß- sen Saum-Gesellschaften, u.a. mit dem Sal-
toleranten Pflanzenarten, einerseits solche, bei-Gamand er (Teu criu m scorodonia).
285Thorsten Aßmann & Anselm Kratochwil Osnabrücker Naturwiss. Mitt. 20121 1995
Ein bezeichnendes Element ist die Gras- 3.4 Der Eichen-Hainbuchenwald- Kom-
nelken-Flur (Diantho deltoidis-Armerietum plex (Abb. 6, Tab. 3)
elongatae) auf leicht anlehmigen trockenen
Sandböden, dichte blumenbunte Rasen mit Die zweite große Hudewald-Landschaft, hier
einem für die Region hohen Anteil an sub- als Carpinion-Landschaft bezeichnet, wird
mediterran (2.8. Ranunculus bulbosus) und durch den Eichen-Hainbuchenwald-Vegeta-
subkontinental verbreiteten Pflanzenarten tionskomplex geprägt. Das Stellario-Carpi-
(2.8. Veronica spicatal. Standörllich vikariiert netum stockt hier als azonale Vegetation auf
zu dem Airetum praecocis des Betulo-Quer- nährstoffreichen und stau- und grundwas-
cetum-Vegetationskomplexes im Lonicero- serfeuchten vergleyten Lehmböden. Die
Fagetum-Vegetationskomplex der Feder- Hudewälder sind auch hier besonders
schwingelrasen, das
Filagini-Vulpietum strauchreich mit bewehrten Arten, so z.B.
myurus, analog wird das Lolio-Cynosuretum mit der Hülse (//ex aquifolium). Ebenso exi-
luzuletosum durch das Lolio-Cynosuretum stieren auch hier - hudebedingt - Ersatzge-
typicum, letzteres auf stärker humosen, sellschaften. Kennzeichnend ist z.B. das nur
anlehmigen Standorten ersetzt. auf anspruchsvolleren Standorten vorkom-
Tab. 3: Ubersichtstabelle: Carpinion-Vegetationskomplex unter Berücksichtigung der hudebedingten Ersatz-
gesellschaften. Die Reihenfolge der Anordnung folgt der zunehmenden Öffnung des Waldes und der stei-
genden lntensität des menschlichen Einflusses.
Garpinion-Vegetationskomplex (Landschaft)
(Eichen-Hainbuchenwälder)
,,Primärwald" Stellario-Garpinetum
(Sternmieren-Eichen-Hainbuchenwald)
ehemaliger Stellario-Carpinetum
Hudewald llex aquifolium- und Strauch-reich
Hudewaldreste und hudebedingte Ersatzgesellschaften
im Gebiet des Carpinion-Vegetationskomplexes:
Hudewald-Reste Stellario-Carpinetum
Heiden Roso-Juniperetum
(Rosen-Wacholderbusch)
Mantel-Gesellschaften Gorno-Prunetum (Prunetalia)
(Schlehenbusch)
Hochstauden-Gesellschaft Artemisietea
Alliarion
Grasland Lolio-Gynosuretum typicum
286Hudelandschaften, Biozönosen
Abb. 6: Stellario-Carpinetum als Element des Eichen-Hainbuchenwald-Komplexes. Das Stellario-Carpinetum
(hier: periclymenetosum) ist hudebedingt reich an Hülse (//ex aqu ifolium). Die Hainbuchen (Carpinus betulus)
tragen Zeichen ehemaliger Schneitelnutzung, heute sind einzelne Stämme aus den ehemaligen Schneitelbe-
reichen des Baumes ausgewachsen (Bentheimer Wald). Foto: Kratochwil.
mende Roso-Juniperetum (s. dazu Barkman schwemmungsbereich Auenwälder (Quer-
1968). Besonders ausgeprägt sind Mantel- co-Ulmetum), die Anklänge an das Stellario-
Gesellschaften, z.B. das Corno-Prunetum. Carpinetum zeigen flrautmann & Lohmeyer
Diese Wuchsorte des Stellario-Carpinetum 1962, Pott & Hüppe 1991), und Galeriewäl-
sind besonders reich an anspruchsvolleren der des Pruno-Fraxinetum. Die hudebeding-
Dornensträuchern, so z.B. der Schlehe (Pru- ten Ersatzgesellschaften entsprechen denen
nus spinosa). Reichhaltig vertreten sind der Carpinion-Landschaft (Corno-Prune-
auch nitrophytische Saumgesellschaften tum, Roso-Juniperetum).
und Staudensäume der Artemisietea oder
Säume des Alliarion. Letztlich schließt auch
hier unter Beweidung das Lolio-Cynosure- 4 Fragestellung und Zielsetzung
tum typicum an. unserer Forschung in den Hude-
landschaften Nordwestdeutsch-
lands
3.5 Auen-Vegetationskomplexe
Extensive und seit dem Neolithikum prakti-
ln den z.T. übersandeten Talbereichen der zierte Formen der Landnutzung haben zu
Ems finden sich im periodischen Über- einer starken parklandschaftsartigen Dif-
287Thorsten Aßmann & Anselm Kratochwil Osnabrücker Naturwiss. Mitt. 20121 1995 ferenzierung der ehemals von Wäldern und ren Gemeinschaften geworden. Aber auch Mooren beherrschten Naturlandschaft in diesen Landschaftstypen blieben Verän- Nordwestdeutschlands geführt. Ein so ent- derungen nicht aus. So haben während der standenes Mosaik unterschiedlichster vergangenen Jahre eine Aufgabe tradierter anth ropo-zoogen-bed ingter Vegetationssta- Nutzungsformen sowie der Einfluß einer dien von Offenland bis hin zu Waldstandor- intensiven Agrar- und Forstwirtschaft auch ten stellt einen Lebensraum von hoher klein- in Hudegebieten zum sukzessiven Umbau räumlicher und zeitlicher Variabilität dar. Er bezeichnender Biozönosen geführt, z.B. weist im Vergleich zur Urlandschaft bezogen über Strukturnivellierung, Eutrophierung, auf eine bestimmte Raumeinheit eine weit- Ruderalisierung u.a. Ein langfristiger Schutz aus größere Faunen- und Florenmannigfal- dieser Lebensräume und Lebensgemein- tigkeit auf. schaften ist deshalb nur möglich, wenn Die Landschaftsgenese folgt dem Prinzip umfassende Kenntnisse über Phyto- und der ,,variety in space" (hoher Grad des stän- Zoozönosen in ihrem ,,biozönotischen Mit- digen räumlichen Wechsels der Faktoren- einander", d.h. in ihrer funktionalen Verknüp- Konstellationen, s. v. Leuwen 1966); somit fung, vorliegen, und wenn Wirkungsweisen entstehen Ökosysteme, die sich durch hohe und -mechanismen gegenwär.tiger Störgrö- Kontinuität in der Erhaltung des Mosaik- ßen und Stressoren auf die bezeichnenden Charakters auszeichnen. Der Stabilisator Zönosen und ihre Einzelglieder bekannt und systemerhaltende Faktor sind in diesem sind. Falle nicht die von Remmert (1991) für Wald- Die von uns durchgeführten und auch in ökosysteme postulierten natürlichen Fakto- Zukunft geplanten Untersuchungen haben ren, sondern gerade die anthropo-zoogenen daher zum Ziel, anhand ausgewählter Bei- Einflüsse, zumal sich diese seit dem Neo- spiele typische Biozönose-Umwelt-Bezie- lithikum auf die Vegetationsentwicklung und hungen in Hudelandschaften beschreiben natürlich auch auf die Tierwelt ausgewirkt und erklären zu können. Sie verstehen sich haben. Ziel unserer Forschung muß es vor somit einerseits als Beitrag zum Schutz von allem sein, solche Extensiv-Biozönosen in wissenschaftlich besonders bedeutsamen ihrem Mosaik-Charaktel in ihrer Dynamik Ökosystem-Typen, andererseits als Beitrag (Wechsel von Degradations- und Regenera- zur Erhaltung aus der Sicht des Natur- und tionskomplexen, s. Pott & Hüppe 1991), in Kulturschutzes besonders wertvoller Le- ihrer landschaftsräumlichen Bindung und bensgemeinschaften. Vor dem skizzierten gesetzmäßig wiederkehrenden Artenkombi- Hintergrund gehören aufbauend auf den nation und Vergesellschaftung zu erfassen vorliegenden geobotanischen und land- und zu typisieren. schaftsökologischen Untersuchungen bio- Die in den vergangenen Jahrzehnten zönologische Fragen zu den wesentlichen durch land- und forstwirtschaftliche lntensi- Forschungszielen: vierung erfolgten Landschaftsveränderun- gen haben ein Überleben ,,extensiver" . eine Erfassung einzelner und für Hude- Lebensgemeinschaften nur noch an weni- landschaften besonders bezeichnender gen Standorten, so z.B. in solchen Hude- Zoozönosen (Schutzwürdigkeit); landschaften ermöglicht. Somit sind sie zu . eine Analyse der Strukturpräferenzen und besonders wichtigen Refugialräumen der Strukturbindung bestimmter Zoozö- bedrohter Tier- und Pflanzenarten sowie de- nosen; 288
Hudelandschaften, Biozönosen
. eine Analyse und Bewertung ehemaliger Bis zur Mitte des 18. Jahrhunderts nah-
und gegenwärtiger Einflußgrößen auf Bio- men Wälder nur einen Bruchteil ihres poten-
zönosen der Hudelandschaften ; tiellen Wuchsgebietes ein (Brüning 1961,
r Untersuchungen zum lndikatorwert be- Hesmer & Schroeder 1963). Diese Situation
stimmter Organismengruppen; wird für das Weser-Ems-Gebiet auch durch
r Analysen zum Flächenbedarf einzelner Kadenmaterial belegt, das unter Leitung des
Zoozönosen im Hinblick auf deren Stabi- preußischen Generalmajors von LeOoq
lität und den Möglichkeiten ihres Schut- (1797-1813) erstellt wurde (von LeCoq
zes unter gegenwärtig wirksamen Einfluß- 1805). Für das östliche Niedersachsen ist
größen (Schutzfähigkeit). eine frühere kartographische Aufnahme ver-
Daraus lassen sich Pflege-Empfehlungen fügbar (Kurhannoversche Landesaufnahme
sowie Forderungen an eine Naturschutzpla- 1 7 64-1 786). Die Abb . 7 zeigt beispiethaft für
nung ableiten (s. auch Pott & Hüppe 1994). die nördliche Ankumer Höhe einen Vergleich
lm folgenden werden nun unter der Berück- der zu Zeilen von LeCoq vorhandenen
sichtigung der unterschiedlichen Land- Waldgebiete sowie die heutige Situation.
schaftstypen (Betu lo-Quercetum-Landschaft , Das Ausmaß der historischen Wddverwü-
Lonicero-Fagetum-Landschaft , Carpinion- stung hatte im nordwestdeutschen Tiefland
Landschaft , Querco-Ulmetum-Landschaft) ein unterschiedliches Ausmaß: Während im
ausgewählte biozönologische Fragestellun- Weser-Ems-Gebiet nur wenige, kleine
gen behandelt. Reliktwälder erhalten blieben, war im Weser-
Elbe-Gebiet der relative Anteil an Wald
erheblich größer. lm Hügelland und in den
5 Biozönologische Untersuchun- Mittelgebirgen Niedersachsens entsprach
gen in Hudelandschaften vor ca. 200 Jahren die Ausdehnung der
Waldfläche fast dem aktuellen Zustand.
5. 1 Reliktarten und Reliktarten-Gemein- Die verbliebenen Waldflächen wurden auf
schaften alter Wälder vielfältige Weise genutzt. Nicht allein durch
Holzgewinnung, sondern vor allem durch
5.1.1 Beispiele für Relikarten alter Wälder andere Nutzungsformen (Waldhude,
Schneitel-, Fallaub- und Plaggenwirlschaft)
Von besonderer Bedeutung für die Wissen- kam es zu einer drastischen Veränderung
schaft und den Naturschutz sind die zu und oft Übernutzung der Wälder. ln diesen
Beginn des 12. und 13. Jahrhunderts und in aufgelichteten Beständen waren die
der Folgezeit mit einem herrschaftlichen Lebensbedingungen für viele Wddarlen
Bann belegten Wälder. Zwar wurden auch sicherlich viel ungünstiger als in geschlosse-
sie stets genutzt, dennoch tragen sie in vie- nen Hochwäldern. Man sollte deshalb
len Fällen mindestens seither, z. T. wahr- erwarten, daß viele Reliktarten alter Wälder
scheinlich aber ununterbrochen seit dem nur lokal bis in die Zeiten mit planmäßiger
Postglazial Wald. Damit stellen sie ,,Relikt- forstlicher Nutzung überleben konnten. Dies
standorte" für Pflanzen und Tiere dar. Unter könnte eine Erklärung für die spärliche Ver-
anthropo-zoogenem Einfluß haben sich dort breitung vieler Reliktarten sein.
Biozönosen primärer Waldstandorte auch in
ihrem Artengefüge erhalten können.
289Thorsten Aßmann & Anselm Kratochwil Osnabrücker Naturwiss. Mitt. 20121 1995 Abb. 7: Ausschnitt der nördlichen Ankumer Höhe mit dem Stift Börstel. Abb. 7a: Um 1800 (von LeCoqsche Landesaufnahme), die Endmoräne ist fast völlig entwaldet und trägt überwiegend eine Heidevegetation, nur in der Umgebung des Stiftes ist ein Hudewald erhalten geblieben. lm Norden dehnt sich das Hahnenmoor aus. Kartengrundlage: Vergrößerung der Karte von Nordwestdeutschland von v. LeOoq (1797-1813), Sec- tion lX. Vervielfältigt mit Erlaubnis des Niedersächsischen Landesverwaltungsamtes - Landesvermessung - 85-664/95. 290
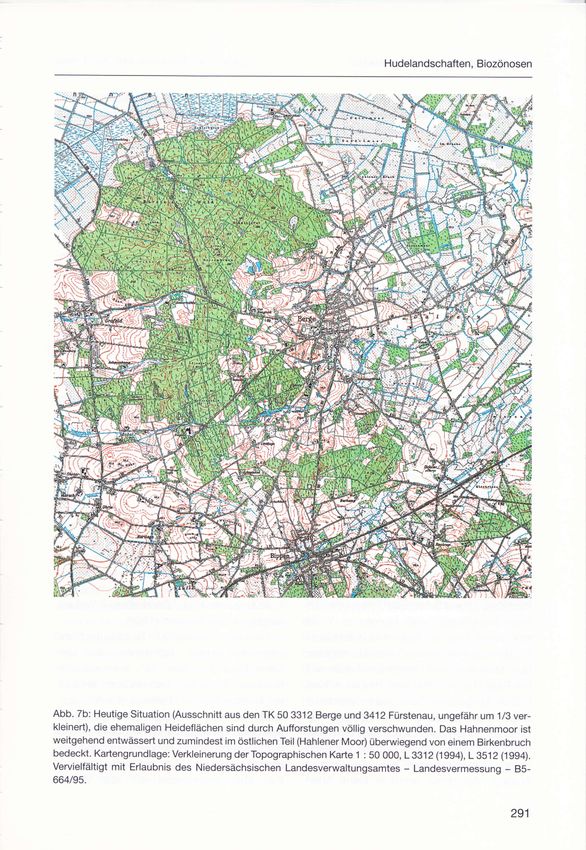
Hudelandschaften, Biozönosen Abb. 7b: Heutige Situation (Ausschnitt aus den TK 50 3312 Berge und 341 2 Fürstenau, ungefähr um 1/3 ver- kleinert), die ehemaligen Heideflächen sind durch Aufforstungen völlig verschwunden. Das Hahnenmoor ist weitgehend entwässert und zumindest im östlichen Teil (Hahlener Moor) überwiegend von einem Birkenbruch bedeckt. Kartengrundlage: Verkleinerung der Topographischen Karte 1 : 50 OOO, L 3312 (1994), L 3512 (1994). Vervielfältigt mit Erlaubnis des Niedersächsischen Landesverwaltungsamtes - Landesvermessung - 85- 664/95.
Thorsten Aßmann & Anselm Kratochwil Osnabrücker Naturwiss. Mitt. 20121 1995
lnsbesondere während der Höhepunkte sondern versucht ausschließlich über die
der mittelalterlichen und frühneuzeitlichen Artenzahl in unterschiedlich alten Wäldern
Waldverwüstungsphasen stellten Bannwäl- die Zönosen zu beschreiben. Dabei stellt er
der lnseln in einer ansonsten vom Men- fest, daß alte Wälder sich durch eine höhere
schen fast völlig entwaldeten Landschaft Artenzahl an Carabidenarten von jungen
dar (s. Darstellungen bei Pott 1993, 1994). Wäldern unterscheiden. Eine Nennung von
Viel kleinflächiger waren die intensiver Reliktarten unterbleibt leider.
genutzten Hofgehölze und Bauernwäldern. ln Mitteleuropa blieben entsprechende
Heute ist der lnselcharakter dieser Wälder Fragestellungen lange unberücksichtigt. Nur
oftmals durch die nach 1800 er{olgten Auf- wenige ältere Arbeiten liegen von Botani-
forstungen verloren gegangen. Besonders kern vor (2. B. Runge 1959). Sporadisch fin-
häufig erfolgten die Aufforstungen auf Hei- den sich auch in der malakozoologischen
deflächen. Durch die zusätzliche Anlage von Literatur Hinweise auf Reliktarten alter Wäl-
Hecken führte man damit - aus der Sicht der (2. B. Ant 1963). Erst in den letzten Jah-
des heutigen Naturschutzes - schon vor lan- ren erlangten diese Themen auch bei uns
ger Zeil eine ,,Biotopvernetzung" durch. vermehrt Beachtung (für die Botanik: Pott
Alte Wälder sind hervorragende Objekte 1994, Wulf & Kelm 1994,Zacharias 1994, für
für ökologische und evolutionsbiologische die Zoologie:Aßmann 1994, 1995).
Studien an Reliktarten und Relikt-Biozöno- Seit 1981 wurden von einem der Autoren
sen. Erste grundlegende Untersuchungen Laufkäfer im nordwestdeutschen Raum so-
hierzu fanden auf den Britischen lnseln statt, wohl in alten (29 Lokalitäten) als auch in jun-
wo aufgrund der günstigen urkundlichen gen Wäldern (32 Lokalitäten)mit jeweils min-
Situation die kontinuierliche Präsenz von destens 10 Barber-Fallen pro Fangstelle (3 x
Wäldern oft bis ins 16. Jahrhundeft zurÜck- für ca. 20 Tage/Jahr oder Ganzjahresfallen)
verfolgt werden kann. Floristische und vege- untersucht (Aßmann 1994). Diese Unter-
tationsökologische Fragen bearbeiteten suchungen zeigen, daß unter den Carabiden
besonders Peterken (1977, 1993) und Peter- einige Arten nur lokal und dabei fast aus-
ken & Game (1981 , 1984). Erste zoologische schließlich an alten Waldstandorten auf-
Untersuchungen erfolgten bereits in der treten:
ersten Hälfte dieses Jahrhunderts (Boycott
1934). Viele der von Boycott (1.c.) für alte Arten mit einer Präferenz für alte Wälder
Wälder angeführten Mollusken gelten auch vom Typ des Lonicero-Fagetum
heute als charakteristische Arten von ,,an- Abax ovalis (Col., Carabidae) - Verbrei-
cient woodlands" und können z. T. als tungskarte bei Aßmann (1994)
Reliktarten auch für das niedersächsische Abax ovalis besiedelt in Norddeutschland
Tiefland bestätigt werden. Weitere Angaben unterschiedlichste Waldtypen, die vom
über Mollusken und Chilopoden finden sich Carici-Fagetum bis zu humusreichen
bei Paul (1975, 1978) und Barber & Keay Buchenwäldern der Endmoränen reichen.
(1988). Epigäische Coleopteren wurden in lm Norddeutschen Tiefland werden aus-
England bisher wenig bei entsprechenden schließlich wasserzügige, sandige Böden
Untersuchungen berücksichtigt. Nur Terrell- bewohnt. Gleichzeitig kommt die Art nicht
Nield (1990) beschäftigte sich mit vergleich- nur in Laubwäldern, sondern auch in Coni-
baren Fragestellungen wie die anderen feren-Forsten von Abax ovalrs ist zwar nicht
Autoren. Allerdings nennt er keine Arten, nur, aber vorwiegend auf alte Wälder
292Hudelandschaften, Biozönosen
beschränkt und kann somit als ,,Reliktart" Limax cinereoniger und Malacolimax
eingestuft werden. Von den 15 bekannten tenellus (Mollusca, Limacidae)
Fundpunkten im niedersächsischen Tiefland Auf die bemerkenswerte Standortstreue
liegen 13 in alten Wäldern. Auch die Verbrei- und den ausgeprägten Reliktarten-Charak-
tungsangaben aus den Niederlanden lassen ter vieler Mollusken haben bereits englische
dort ein ähnliches Verhalten der Art vermu- Malakozoologen hingewiesen (2. B. Boycott
ten fiurin et al. 1977, Turin & Heijermann 1934, Paul 1975, 1978). Die Schnegel-Arten
1e88). Limax cinereoniger und Malacolimax tenel-
Carabus glabrafus (Col., Carabidae) - Ver- /us zeigen vermutlich auch in Norddeutsch-
breitungskarte bei Aßmann (199a) land eine reliktäre Verbreitung in alten Wäl-
Wälder und Forsten unterschiedlicher dern. Erste von uns im Naturschutzgebiet
Ausprägung (Buchen- und Eichenbestände, ,,Lüneburger Heide" durchgeführte Untersu-
Fichten-, Kiefern- und Lärchenforsten) sind chungen ergaben ausschließlich Funde in
die Lebensräume von Carabus glabratus im Bannwäldern (2. B. ,,Hainköpen" bei Wil-
Norddeutschen Tiefland. lm nördlichen sede) und in einem kleinen, aber alten Hof-
Europa und in den Hochlagen der Mittelge- gehölz (Hof Möhr bei Schneverdingen), das
birge werden auch baumfreie Lebensräume auch auf der ersten kartographischen Bear-
besiedelt ffundren, z. T. mit Permafrostbö- beitung bereits verzeichnet ist (Kurhanno-
den, Hochheiden). Die Vorkommen in der versche Landesaufnahme 1 764-1 786).
niedersächsischen Tiefebene beschränken
sich weitgehend auf alte Wälder. Populatio- Arten mit einer Präferenz für alte Wälder
nen außerhalb alter Wälder sind aus dem vom Typ des Carpinion
Naturschutzgebiet ,,Lüneburger Heide" Abax parallelus (Col., Carabidae) - Ver-
bekannt, wo alte Wälder heute in kontinuier- breitungskarte bei Aßmann (1 994)
lich aufgeforsteten, angrenzenden Berei- Diese stenotope Art ist auf feuchte Wälder
chen liegen. Gebiete, die an alte Wälder beschränkt, im Norddeutschen Tiefland sind
grenzen, konnten in den letzten 200 Jahren 21 Populationen bekannt. lm Norddeut-
von Carabus g/abratus wieder besiedelt schen Tiefland werden überwiegend Stella-
werden. Einen erst jetzt bekannt geworde- rio-Carpineten besiedelt (2. B. ,,Urwald Has-
nen Fundpunkt stellt ein Wald südlich von bruch",,,Urwald Herrenholz", Bentheimer
Ankum (Ldkr. Osnabrück) dar: das ,,Rüsse- Wald, ,,Clemenswerther Gehölz"). Die
ler Holz". Dieses Waldgebiet wird bei den bereits von Barner (1954) hervorgehobene
kartographischen Aufnahmen von LeOoq Bevorzugung lehmiger Böden hängt viel-
zwar nicht aufgeführt, allerdings ist auch hier leicht mit der Brutfürsorge der Weibchen
ein langfristiges Überdauern möglich, da zusammen: Wenige Tage vor der Eiablage
das angrenzende Gut ,,Starten" vermutlich wird eine Brutkammer im Boden angelegt.
in historischer Zeil über ein Hofgehölz ver- Das Weibchen bleibt in diesem Hohlraum,
fügt haben dürfte. ln Hofgehölzen der Lüne- bis die Larven geschlüpft sind (Löser 1970,
burger Heide konnten Reliktarten überleben Brandmayr 1977). Die Anlage solcher Brut-
(s. unten). Noch nicht bei Aßmann (1994) kammern ist nur dort möglich, wo Lehmbö-
wurde ein Reliktvorkommen im nördlichen den existieren.
Bereich der Ankumer Höhe (Stiftswald bei
Börstel) angeführt. Auf dieses Beispiel wird
später noch intensiver eingegangen.
293Thorsten Aßmann & Anselm Kratochwil Osnabrücker Naturwiss. Mitt. 20121 1995 Lith o bi u s cu tti pes (Lithobiomorpha, Litho- nis bestätigt werden (Feldmann 1981). Am biidae) Nordwestrand des Verbreitungsgebietes im Der Chilopode Lithobius curtipes kann mit Weser-Ems-Gebiet ist Salamand ra salaman- hoher Wahrscheinlichkeit als Charakterart dra noch spärlicher verbreitet als im Westfä- alter Wälder angesehen werden, wie bereits lischen Tiefland. Viele Fundpunkte sind Barber & Keay (1988) für die Britischen bereits seit langem bekannt und liegen alle lnseln hervorheben. Vossel (1994) konnte L' in alten Wäldern (2. B. ,,Urwald Hasbruch", curtipes im Bentheimer Wald nachweisen. vgl. Borcherding 1887). Trotzdem wurden in Außerdem tritt die Art im Herrenholz bei den letzten Jahren auch noch bisher unbe- Vechta auf. Jeekel (1964) nennt noch eine kannte Populationen entdeckt, die aus- weitere Population aus dem ,,Forst Beimoor" schließlich mit alten Wäldern in Beziehung in Schleswig-Holstein.,,Forst Beimoor" stellt stehen (2. B. zwei Populationen in der einen alten Waldstandort dar, der bei der Umgebung des ,,Hasbruches" bei Delmen- ,,Varendorf-Kartierung" (Landesaufnahme horst, Stegink mdl. Mitt.). Während die des Herzogtums Holstein 1789-1796) und Oldenburger und Zevener Geest besiedelt bei der,,Preußischen Landesaufnahme" von sind, fehlt die Art in der von Moor- und Fluß- 1878, die während der maximalen Waldver- niederungen eingeschlossenen Osterholzer wüstung in Schleswig-Holstein durchgefühtt Geest. Offenbar hat S. sa/amandra wie auch wurde, verzeichnet ist. der Bergmolch (Iriturus a/pestns) diese Vermutlich kommen nicht nur im Nord- Gebiete postglazial nicht erreicht (Nettmann deutschen Tiefland, sondern auch in ande- 1991). lm südwestlichen Niedersachsen ist ren Regionen Reliktarten alter Wälder vor. der Feuersalamander ausgesprochen sel- Ant et al. (1984) konnten für den ,,Hamba- ten. Eine Überprüfung von faunistischen cher Forst" (Niederrhein) Trichotichnus lae- Nachweisen aus dem Tiefland ergab auch vicollis und Abax ovalrs als Reliktvorkommen hier eine Kongruenz mit alten Waldstandor- feststellen. Der Reliktcharakter dieser Vor- ten (2. B. ,,Stiftswald" bei Börstel, Ldkr. kommen wird auch dadurch gestützt, daß Osnabrück, vgl. Schreiber & Wellinghorst beide Arten in der betreffenden Faunistik fÜr 1e91). den Niederrhein nicht aufgeführt werden (Koch 1968). Das gleiche gilt auch für Cara- Kausalanalytische U ntersuchungen zur F rage bus glabratus, der vor wenigen Jahren durch der Reliktvorkommen von Carabiden Scharf (mündl. Mitt.) in einem Wald am Nie- Während sich die oben vorgestellten Cara- derrhein (,,Diersfordter Forst") nachgewie- bidenarten nicht oder nur unwesentlich in sen wurde. Hier konnte bisher Abax ovalis den letzten 200 Jahren ausgebreitet haben, nicht gefunden werden. sind andere Arten heute in Wäldern weit ver- Salamandra salamandra (Amphibia' breitet. Als Beispiel möge an dieser Stelle Salamandridae) der Vergleich der Verbreitung von Carabus Der Feuersalamander stellt im Norddeut- glabratus und Carabus problematicus im schen Tiefland ebenfalls eine ,,exklusive" nördlichen Teil der Ankumer Höhe dienen Charakterart alter Wälder dar. FÜr die West- (Abb. I und 9). Dieses Gebiet wurde hin- fälische Bucht wurde das ausschließliche sichtlich seiner Waldentwrcklung von der Auftreten in alten Wäldern bereits von West- ersten kartographischen Bearbeitung (um hoff (1893) erkannt. Auch bei neueren groß- 1800) bis heute bereits in Abb. 7 vorgestellt. flächigen Kartierungen konnte dieses Ergeb- Deutlich ist zu erkennen, daß sich Carabus 294
Hudelandschaften, Biozönosen
glabratus praktisch nicht ausgebreitet hat findet sich auch im südlichen Hümmling (-
und heute noch im ,,Stiftswald" vorkommt, Aßmann 1994).
während Carabus problematicus auch in Angesichts der Unterschiede im Verbrei-
jungen Wäldern weit verbreitet ist (Abb. 9). tungsbild beider Arten stellt sich die Frage,
Eine ähnliche Verteilung der beiden Arten warum Carabus glabratus nur Reliktvorkom-
NNs\ .N \
I zKM I
STIFT
"\
S '\t
N s \\....\
Abb. 8: Verbreitung von Carabus glabratus im nördlichen Teil der Ankumer Höhe (vgl. auch Abb. 7). Kreuz-
schraffiert sind alte Waldgebiete, einfach schraffiert neuzeitliche Aufforstungen und Birkenbestände auf ent-
wässertem Hochmoorboden. Gefüllte Kreise kennzeichnen Nachweise, offene Kreise Fangstellen, an denen
die Art nicht nachgewiesen werden konnte.
295Thorsten Aßmann & Anselm Kratochwil Osnabrücker Naturwiss. Mitt. 2Ol21 1995
men aufweist, während Carabus problema- Erklärung unterschiedlicher Verbreitungsbil-
ticus sich in Norddeutschland nach den der angeführt (vgt. Endter'1982):
Wiederaufforstungen großflächig ausbreiten 1) ökologische Gründe: Aufgrund einer spe-
konnte. ln der modernen Biogeographie zifischen Habitatbindung ist das Vorkom-
werden zwei alternative Hypothesen zur men an alte Wälder gebunden. Außerhalb
\ \**Nt
N
N
s,N"
SN
STIFT
s\ \
Abb' 9: Verbreitung von Carabus problematicus im nördlichen Teil der Ankumer Höhe (vgl.
auch Abb. 7).
Kreuzschraffiert sind alte waldgebiete, einfach schraffiert neuzeitliche Aufforstungen
und Birkenbestände auf
entwässertem Hochmoorboden. Gefüllte Kreise kennzeichnen Nachweise, offLne
Kreise Fangstellen, an
denen die Art nicht nachgewiesen werden konnte.
296Hudelandschaft en, Biozönosen
Abb. 10: Rund,,enclosure" im Hofgehölz Möhr (Lüneburger Heide bei Schneverdingen). Foto Aßmann.
solcher Habitate kann Carabus glabratus Gebieten, die zuvor nicht von dieser Art
nicht überleben. besiedelt waren, Tiere in einem solchen
2)historische Gründe: Carabus glabratus ,,Gehege" ausgesetzt werden. lm Verlauf
kann außerhalb alter Wälder leben, hat von mehreren Jahren kann die Populations-
junge Wälder aber noch nicht erreicht. dynamik dieser Tiere dann verglichen wer-
zu 1) ökologische Gründe den mit derjenigen im natürlichen Habitat.
Ökologische Gründe können in der Regel zu 2) historische Gründe
nur für solche Arten als verbreitungsbestim- Falls Carabus glabratus aufgrund der Um-
mend angenommen werden, bei denen Hin- weltparameter auch in jungen Wäldern vor-
weise auf eine enge Habitatbindung vodie- kommen kann, liegt die Annahme nahe, daß
gen; bei zahlreichen Carabiden sind ökolo- das Ausbreitungspotential der Art, verglichen
gische Gründe wahrscheinlich für ihre mit dem von Carabus problematicus, gering
Verbreitungen (vgl. Angaben zur Bindung an ist. Diese Annahme läßt sich u. a. durch
biotische und abiotische Parameter beiThiele Orientierungsversuche in Rund,,enclosures"
1977). Auch für Abax paral/e/us können öko- (Rundgehegen), andererseits durch Radar-
logische Gründe bestimmend für die lokalen untersuchungen überprüfen.
Vorkommen in alten Wäldern sein. Bei einer lm Hofgehölz Möhr (östlich von Schnever-
so eurytopen Art wie Carabus glabratus er- dingen, Lüneburger Heide) konnte ein
scheinen diese unwahrscheinlich, wenn sie Rund,,enclosure" errichtet werden (Durch-
auch bei aktuellem Kenntnisstand nicht mit messer 20 m, Folie 40 cm tief in den Wald-
letzter Sicherheit ausschließbar sind. Auf- boden eingegraben, ca. 50 cm über den
schluß können zu diesem Faktor nur,,enclo- Boden emporragend) (Abb. 10). ln einem
sure"-Versuche mit ausgesetzten Carabus Abstand von 10 m befanden sich an der
glabratus-Gruppen geben. Dazu müssen in lnnenseite der Folie ebenerdig eingegra-
297Sie können auch lesen

















































